
Rapid Spread of the Invasive Brown Alga Rugulopteryx okamurae in a National Park in Provence (France, Mediterranean Sea)

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Observation and Species Expansion
3.2. Life Cycle and Impact
4. Discussion
4.1. Spread
4.2. Origin
4.3. Impact
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Content Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Studied Specimens
References
- Galil, B.S. A sea under siege-alien species in the Mediterranean. Biol. Invasions 2000, 2, 177–186. [Google Scholar] [CrossRef]
- Zenetos, A. Trend in alien species in the Mediterranean. An answer to Galil, 2009 “Taking stock: Inventory of alien species in the Mediterranean Sea”. Biol. Invasions 2010, 12, 3379–3381. [Google Scholar] [CrossRef]
- Ulman, A.; Ferrario, J.; Occhipinti-Ambrogi, A.; Arvanitidis, C.; Bandi, A.; Bertolino, M.; Bogi, C.; Chatzigeorgiou, G.; Çiçek, B.A.; Deidun, A.; et al. A massive update of non-indigenous species records in Mediterranean marinas. PeerJ 2017, 5, 1–59. [Google Scholar]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A. East is east and West is west? Management of marine bioinvasions in the Mediterranean Sea. Estuar. Coast. Shelf Sci. 2018, 201, 7–16. [Google Scholar] [CrossRef]
- Boudouresque, C.F.; Verlaque, M. Is global warming involved in the success of seaweed introductions in the Mediterranean Sea? In Seaweeds and Their Role in Globally Changing Environments; Israel, A., Einav, R., Seckbach, J., Eds.; Springer Publisher: Dordrecht, The Netherlands, 2010; pp. 31–50. [Google Scholar]
- Boudouresque, C.F.; Perret-Boudouresque, M.; Verlaque, M. Donor and recipient regions for exotic species of marine macrophytes: A case of unidirectional flow, the Mediterranean Sea. Rapp. Comm. Intl. Mer Méditerr. 2016, 41, 426. [Google Scholar]
- Ruiz, H.; Ballantine, D.L. Occurrence of the seagrass Halophila stipulacea in the tropical West Atlantic. Bull. Mar. Sci. 2004, 75, 131–135. [Google Scholar]
- Willette, D.A.; Ambrose, R.F. The distribution and expansion of the invasive seagrass Halophila stipulacea in Dominica, West Indies, with a preliminary report from St. Lucia. Aquat. Bot. 2009, 91, 137–142. [Google Scholar] [CrossRef]
- Maréchal, J.P.; Meesters, E.H.; Vedie, F.; Hellio, C. Occurrence of the alien seagrass Halophila stipulacea in Martinique (French West Indies). Mar. Biodiv. Rec. 2013, 6, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Verlaque, M.; Steen, F.; De Clerck, O. Rugulopteryx (Dictyotales, Phaeophyceae), a genus recently introduced to the Mediterranean. Phycologia 2009, 48, 536–542. [Google Scholar] [CrossRef]
- Boudouresque, C.F.; Klein, J.; Ruitton, S.; Verlaque, M. Biological invasion: The Thau lagoon, a Japanese biological island in the Mediterranean Sea. In Global Change: Mankind-Marine Environment Interactions; Ceccaldi, H.J., Dekeyser, I., Girault, M., Stora, G., Eds.; Springer Publisher: Dordrecht, The Netherlands, 2011; pp. 151–156. [Google Scholar]
- Ocaña, O.; Alfonso-Carrillo, J.M.; Ballesteros, E. Massive proliferation of a dictyotalean species (Phaeophyceae, Ochrophyta) through the Strait of Gibraltar. Rev. Acad. Canaria Cienc. 2016, 28, 165–169. [Google Scholar]
- García-Gómez, J.C.; Sempere-Valverde, J.; Ostalé-Valriberas, E.; Martínez, M.; Olaya-Ponzone, L.; González, A.R.; Espinosa, F.; Sánchez-Moyano, E.; Megina, C.; Parada, J.A. Rugulopteryx okamurae (E.Y. Dawson) I.K. Hwang, W.J. Lee & H.S. Kim (Dictyotales, Ochrophyta), alga exótica “explosiva” en el estrecho de Gibraltar. Observaciones preliminares de su distribución e impacto. Almoraima. Rev. Estud. Campogibraltareños 2018, 49, 97–113. [Google Scholar]
- García-Gómez, J.C.; Sempere-Valverde, J.; González, A.R.; Martínez-Chacón, M.; Olaya-Ponzone, L.; Sánchez-Moyano, E.; Ostalé-Valriberas, E.; Megina, C. From exotic to invasive in record time: The extreme impact of Rugulopteryx okamurae (Dictyotales, Ochrophyta) in the strait of Gibraltar. Sci. Total Environ. 2020, 704, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sempere-Valverde, J.; García-Gómez, J.C.; Ostalé-Valriberas, E.; Martínez, M.; Olaya-Ponzone, L.; González, A.R.; Sánchez-Moyano, E.; Megina, C.; Parada, J.A.; Espinosa, F. Expansion of the exotic brown algae Rugulopteryx okamurae (E.Y. Dawson) I.K. Hwang, W.J. Lee & H.S. Kim in the Strait of Gibraltar, Proceedings of the 1st Mediterranean symposium on the non-indigenous species, Antalya, Turkey, 18 January 2019; Langar, H., Ouerghi, A., Eds.; RAC/SPA Publisher: Tunis, Tunisia, 2019; pp. 107–108. [Google Scholar]
- García-Gómez, J.C.; Florido, M.; Olaya-Ponzone, L.; Díaz de Rada, J.R.; Donázar-Aramendia, I.; Chacón, M.; Quintero, J.J.; Magariño, S.; Megina, C. Monitoring extreme impacts of Rugulopteryx okamurae (Dictyotales, Ochrophyta) in El Estrecho Natural Park (Biosphere Reserve). Showing radical changes in the underwater seascape. Front. Ecol. Evol. 2021, 9, 1–18. [Google Scholar] [CrossRef]
- Hwang, I.K.; Lee, W.J.; Kim, H.S.; De Clerck, O. Taxonomic reappraisal of Dilophus okamurae (Dictyotales, Phaeophyta) from the western Pacific Ocean. Phycologia 2009, 48, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Meinesz, A. Les Aires marines naturelles protégées du Parc national des Calanques. In Méditerranée mer Vivante, 20th ed.; Meinesz, A., Ed.; Lions Club Nice Doyen Publisher: Nice, France, 2020; pp. 232–234. [Google Scholar]
- Parc National des Calanques. Available online: www.calanques-parcnational.fr.2021 (accessed on 12 May 2021).
- Blanfuné, A.; Boudouresque, C.F.; Perret-Boudouresque, M.; Ruitton, S.; Sevaux, L.; Verlaque, M.; Thibaut, T. Inventaire des Macroalgues du Parc national des Calanques; Parc national des Calanques and MIO Publisher: Marseille, France, 2017; pp. 1–33. [Google Scholar]
- Verlaque, M.; Ruitton, S.; Mineur, F.; Boudouresque, C.F. CIESM Atlas of exotic species. In 4. Macrophytes; Briand, F., Ed.; CIESM Publisher: Monaco, Monaco, 2015; pp. 1–362. [Google Scholar]
- Thibaut, T.; Pinedo, S.; Torras, X.; Ballesteros, E. Long-term decline of the populations of Fucales (Cystoseira spp. and Sargassum spp.) in the Albères coast (France, North-western Mediterranean). Mar. Pollut. Bull. 2005, 50, 1472–1489. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfuné, A.; Boudouresque, C.F.; Verlaque, M. Decline and local extinction of Fucales in the French Riviera: The harbinger of future extinctions Mediterr. Mar. Sci. 2015, 16, 206–224. [Google Scholar]
- Ifremer. Réseau Velyger-Observatoire de la Reproduction et du Recrutement de l’huître en France. Lagune de Thau. 2021. Available online: https://wwz.ifremer.fr/velyger/Acces-aux-Donnees/Lagune-de-Thau/Temperature (accessed on 13 April 2021).
- Agatsuma, Y.; Kuwahara, Y.; Taniguchi, K. Life cycle of Dilophus okamurae (Phaeophyceae) and its associated invertebrate fauna in Onagawa Bay, Japan. Fish. Sci. 2005, 71, 1107–1114. [Google Scholar] [CrossRef]
- Ribera, M.A.; Boudouresque, C.F. Introduced marine plants, with special reference to macroalgae: Mechanisms and impact. In Progress in Phycological Research; Round, F.E., Chapman, D.J., Eds.; Biopress Ltd. Publisher: Bristol, UK, 1995; Volume 11, pp. 187–268. [Google Scholar]
- Feldmann, J. Sur la parthénogénèse du Codium fragile (Sur.) Hariot dans la Méditerranée. C.R. Acad. Sci. 1956, 243, 305–307. [Google Scholar]
- Meinesz, A. Modes de dissémination de l’algue Caulerpa taxifolia introduite en Méditerranée. Rapp. Comm. Intl. Mer Méditerr. 1992, 33, 44. [Google Scholar]
- Žuljević, A.; Antolić, A. Synchronous release of male gametes of Caulerpa taxifolia (Caulerpales, Chlorophyta) in the Mediterranean Sea. Phycologia 2000, 39, 157–159. [Google Scholar] [CrossRef]
- Verlaque, M. Contribution à la flore des algues marines de Méditerranée. Espèces rares ou nouvelles pour les côtes françaises. Bot. Mar. 1989, 32, 101–113. [Google Scholar] [CrossRef]
- Cormaci, M.; Furnari, G.; Alongi, G.; Serio, D. On three interesting marine red algae (Ceramiales, Rhodophyta) from the Mediterranean Sea. G. Bot. Ital. 1994, 128, 1001–1006. [Google Scholar] [CrossRef]
- Nikolić, V.; Žuljević, A.; Antolić, B.; Despalotović, M.; Cvitković, I. Distribution of invasive red alga Womersleyella setacea (Hollenberg) R.E. Norris (Rhodophyta, Ceramiales) in the Adriatic Sea. Acta Adriatica 2010, 51, 195–202. [Google Scholar]
- Verdú Baeza, J. España y los problemas de aplicación del convenio de aguas de lastre en el área del estrecho de gibraltar. A propósito del alga invasora Rugulopterix okamurae. Rev. Esp. Derecho Intl. Sección Estudios 2020, 72, 261–287. [Google Scholar] [CrossRef]
- Guihou, K.; Marmain, J.; Ourmières, Y.; Molcard, A.; Zakardjian, B.; Forget, P. A case study of the mesoscale dynamics in the North-Western Mediterranean Sea: A combined data–model approach. Ocean Dynam. 2013, 63, 793–808. [Google Scholar] [CrossRef]
- Miller, R.L. Ascophyllum nodosum: A source of exotic invertebrates introduced into west coast near-shore marine waters. Veliger 1969, 12, 230–231. [Google Scholar]
- Mineur, F.; Davies, A.J.; Maggs, C.A.; Verlaque, M.; Johnson, M.P. Fronts, jumps and secondary introductions suggested as different invasion patterns in marine species, with an increase in spread rates over time. Proc. Roy. Soc. B Biol. Sci. 2010, 277, 2693–2701. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Barranco, C.; Muñoz-Gómez, B.; Guerra-García, J.M.; Altamirano, M.; Ostalé-Valriberas, E.; Moreira, J. Can invasive habitat-forming species play the same role as native ones? The case of the exotic marine macroalga Rugulopteryx okamurae in the Strait of Gibraltar. Biol. Invasions 2019, 21, 3319–3334. [Google Scholar] [CrossRef]
- Paula, J.C.D.; Vallim, M.A.; Teixeira, V.L. What are and where are the bioactive terpenoids metabolites from Dictyotaceae (Phaeophyceae). Rev. Bras. Farmacogn. 2011, 21, 216–228. [Google Scholar] [CrossRef] [Green Version]
- García-Gómez, J.C.; Floridon, M.; Olaya-Ponzone, L.; Sempere-Valverde, J.; Megina, C. The invasive macroalga Rugulopteryx okamurae: Substrata plasticity and spatial colonization pressure on resident macroalgae. Front. Ecol. Evol. 2021, 9, 1–15. [Google Scholar] [CrossRef]
- Sempere-Valverde, J.; Ostalé-Valriberas, E.; Maestre, M.; González Aranda, R.; Bazairi, H.; Espinosa, F. Impacts of the non-indigenous seaweed Rugulopteryx okamurae on a Mediterranean coralligenous community (Strait of Gibraltar): The role of long-term monitoring. Ecol. Indic. 2021, 121, 1–5. [Google Scholar] [CrossRef]
- Simberloff, D. No reserve is an island: Marine reserves and nonindigenous species. Bull. Mar. Sci. 2000, 66, 567–580. [Google Scholar]
- Katsanevakis, S.; Issaris, Y.; Poursanidis, D.; Thessalou-Legaki, M. Vulnerability of marine habitats to the invasive green alga Caulerpa racemosa var. cylindracea within a marine protected area. Mar. Environ. Res. 2010, 70, 210–218. [Google Scholar]
- Carlton, J.T.; Keith, I.; Ruiz, G.M. Assessing marine bioinvasions in the Galápagos Islands: Implications for conservation biology and marine protected areas. Aquat. Invasions 2019, 14, 1–20. [Google Scholar] [CrossRef]
- Giakoumi, S.; Pey, A.; Di Franco, A.; Francour, P.; Kizilkaya, Z.; Arda, Y.; Raybaud, V.; Guidetti, P. Exploring the relationships between marine protected areas and invasive fish in the world’s most invaded seas. Ecol. Appl. 2019, 29, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mannino, A.M.; Balistreri, P. Invasive alien species in Mediterranean Marine Protected Areas: The Egadi Islands (Italy) case study. Biodiversity 2021, 10, 1–11. [Google Scholar] [CrossRef]
- Elton, C.S. The Ecology of Invasions by Animals and Plants; Methuen: London, UK, 1958. [Google Scholar]
- Boudouresque, C.F.; Verlaque, M. An overview of species introduction and invasion processes in marine and coastal lagoon habitats. Cah. Biol. Mar. 2012, 53, 309–317. [Google Scholar]
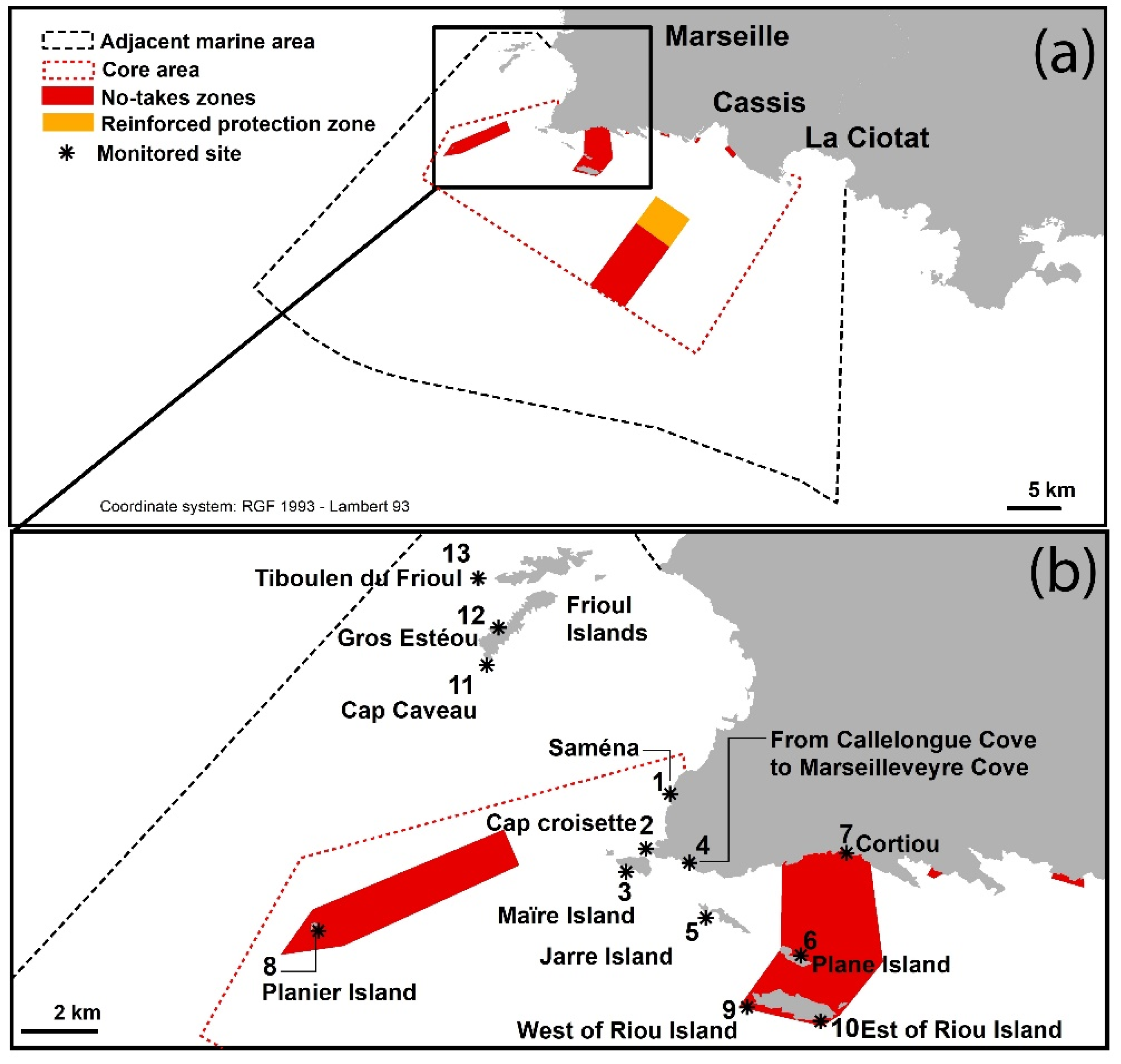




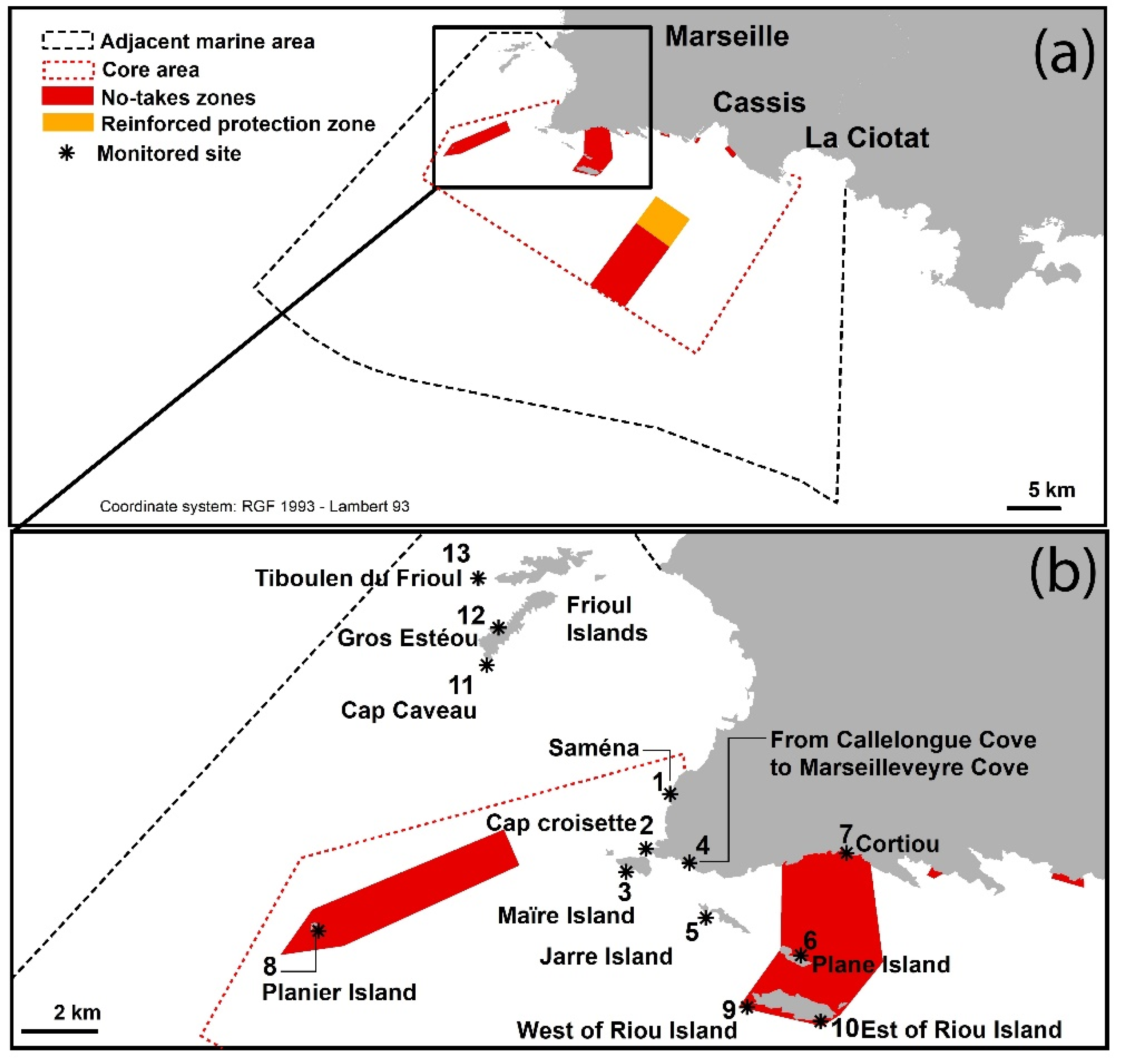{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Sites | 2018 | 2019 | 2020 |
|---|---|---|---|---|
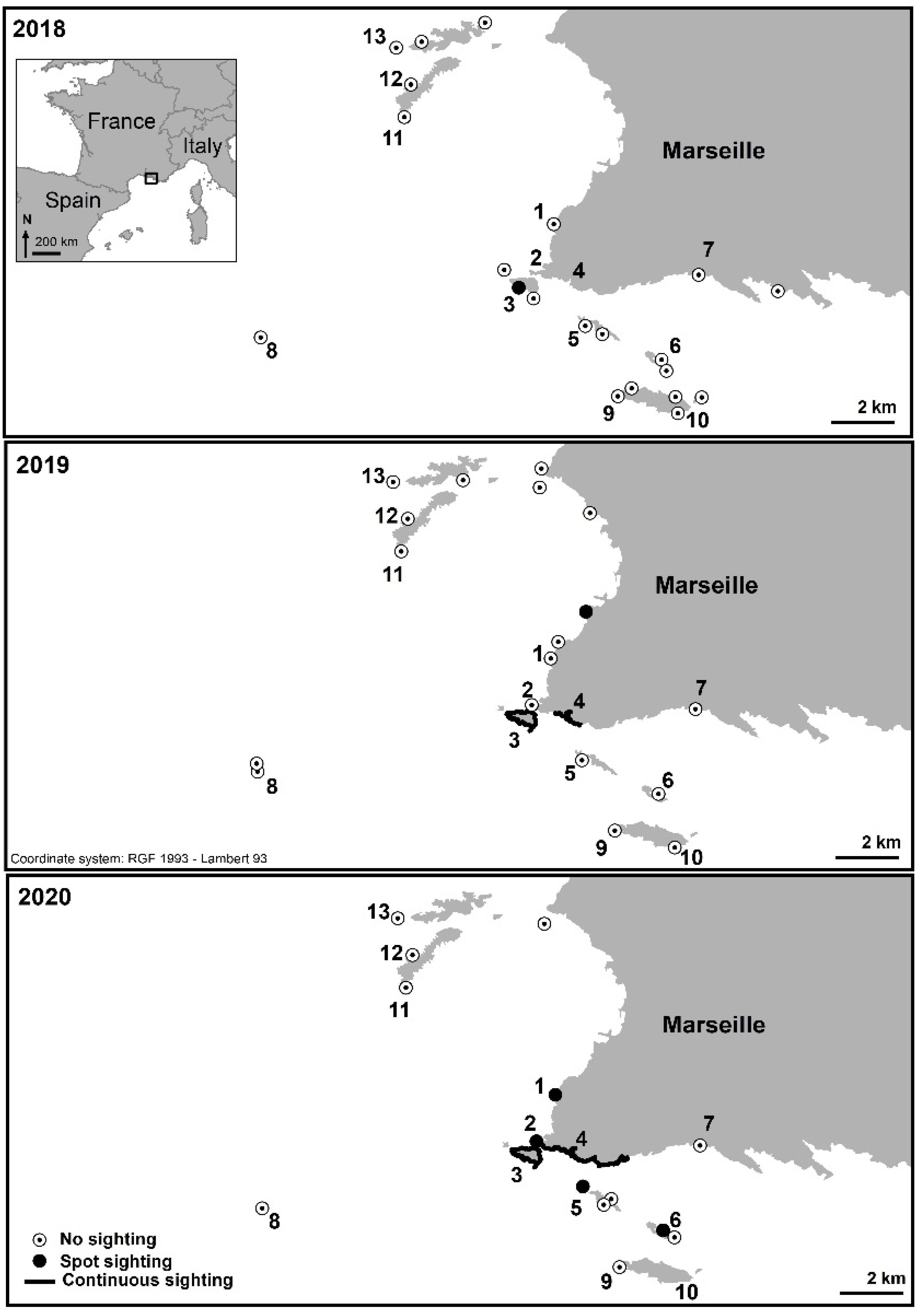
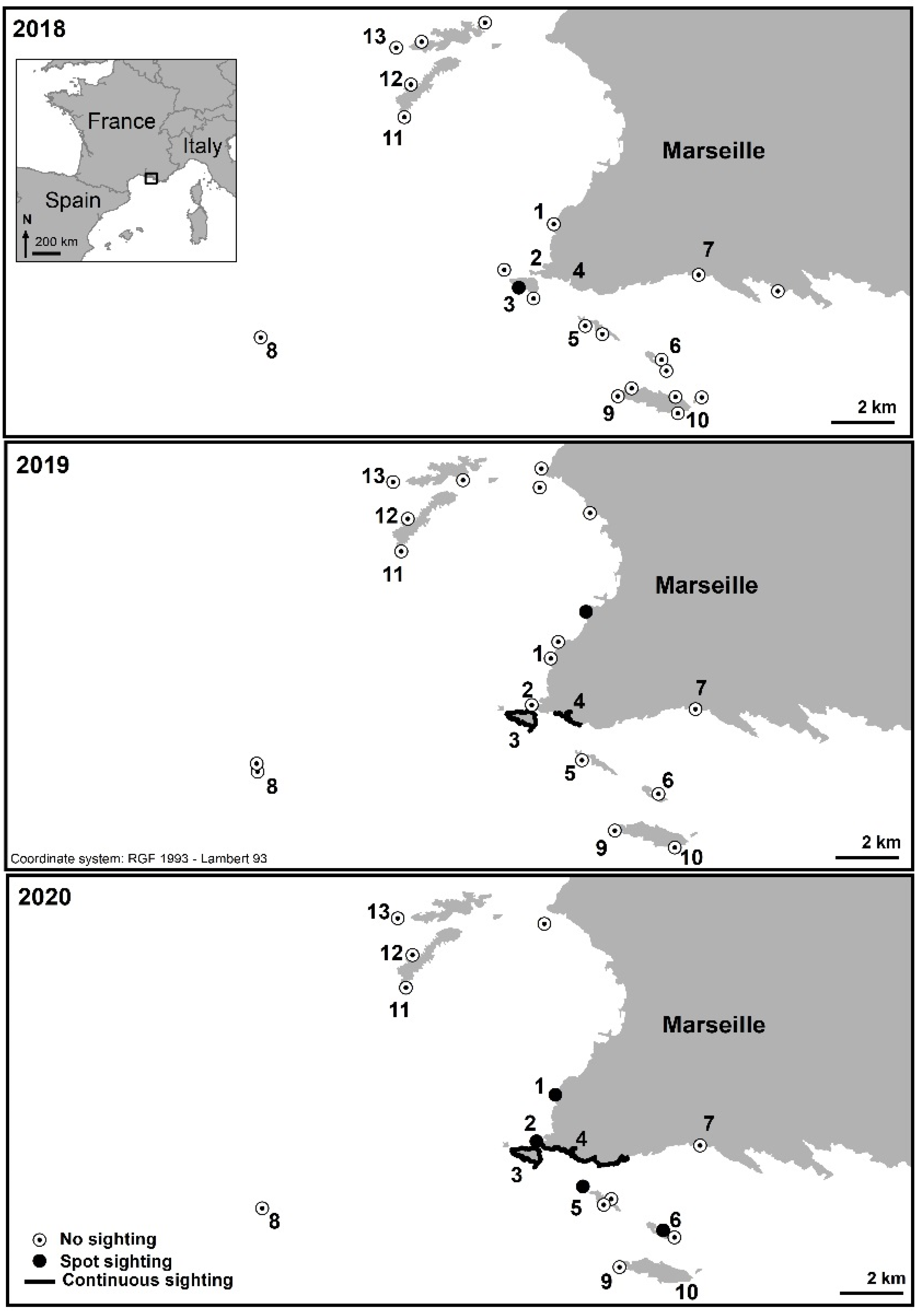
| 1 | Saména | No sighting | No sighting | Spot sighting, from 3 to 4 m deep. Percentage cover < 1%. |
| 2 | Cap Croisette | No sighting | No sighting | Spot sighting. Percentage cover < 1% from 1.4 to 3 m deep, reaching 20% at 3–6 m deep. |
| 3 | Maïre Island | No in situ sighting but spot sighting in the south of the island (a posteriori photo analysis). | Continuous sighting all around the island. Percentage cover reaching 85% at 5 m deep and 69% at 12 m deep. Maximum sighted depth: 20 m. | Continuous sighting all around the island. Maximum sighted depth: 25 m. |
| 4 | Coast from Callelongue Cove to Marseilleveyre Cove | No sighting | Continuous sighting, from 3 to 12 m deep. Percentage cover reaching 60% at this depth. | Continuous sighting. Percentage cover reaching 80% in some areas, around 6 m deep. |
| 5 | Jarre Island | No sighting | No sighting | Spot sighting only on the north side of the island, at 3 to 5 m deep. Percentage cover < 1%. |
| 6 | Plane Island | No sighting | No sighting | Spot sighting only on the northwest side of the island, at 3 to 5 m deep. Percentage cover around 2%. |
| 7 | Cortiou | No sighting | No sighting | No sighting |
| 8 | Planier Island | No sighting | No sighting | No sighting |
| 9 | West of Riou Island | No sighting | No sighting | No sighting |
| 10 | East of Riou Island | No sighting | No sighting | No sighting |
| 11 | Cap Caveau, Frioul Island | No sighting | No sighting | No sighting |
| 12 | Gros Estéou, Frioul Island | No sighting | No sighting | No sighting |
| 13 | Tiboulen du Frioul, Frioul Island | No sighting | No sighting | No sighting |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruitton, S.; Blanfuné, A.; Boudouresque, C.-F.; Guillemain, D.; Michotey, V.; Roblet, S.; Thibault, D.; Thibaut, T.; Verlaque, M. Rapid Spread of the Invasive Brown Alga Rugulopteryx okamurae in a National Park in Provence (France, Mediterranean Sea). Water 2021, 13, 2306. https://doi.org/10.3390/w13162306
Ruitton S, Blanfuné A, Boudouresque C-F, Guillemain D, Michotey V, Roblet S, Thibault D, Thibaut T, Verlaque M. Rapid Spread of the Invasive Brown Alga Rugulopteryx okamurae in a National Park in Provence (France, Mediterranean Sea). Water. 2021; 13(16):2306. https://doi.org/10.3390/w13162306
Chicago/Turabian StyleRuitton, Sandrine, Aurélie Blanfuné, Charles-François Boudouresque, Dorian Guillemain, Valérie Michotey, Sylvain Roblet, Delphine Thibault, Thierry Thibaut, and Marc Verlaque. 2021. "Rapid Spread of the Invasive Brown Alga Rugulopteryx okamurae in a National Park in Provence (France, Mediterranean Sea)" Water 13, no. 16: 2306. https://doi.org/10.3390/w13162306
APA StyleRuitton, S., Blanfuné, A., Boudouresque, C.-F., Guillemain, D., Michotey, V., Roblet, S., Thibault, D., Thibaut, T., & Verlaque, M. (2021). Rapid Spread of the Invasive Brown Alga Rugulopteryx okamurae in a National Park in Provence (France, Mediterranean Sea). Water, 13(16), 2306. https://doi.org/10.3390/w13162306