
CO2 and CH4 Emissions from an Arid Fluvial Network on the Chinese Loess Plateau


Abstract
1. Introduction
2. Materials and Methods
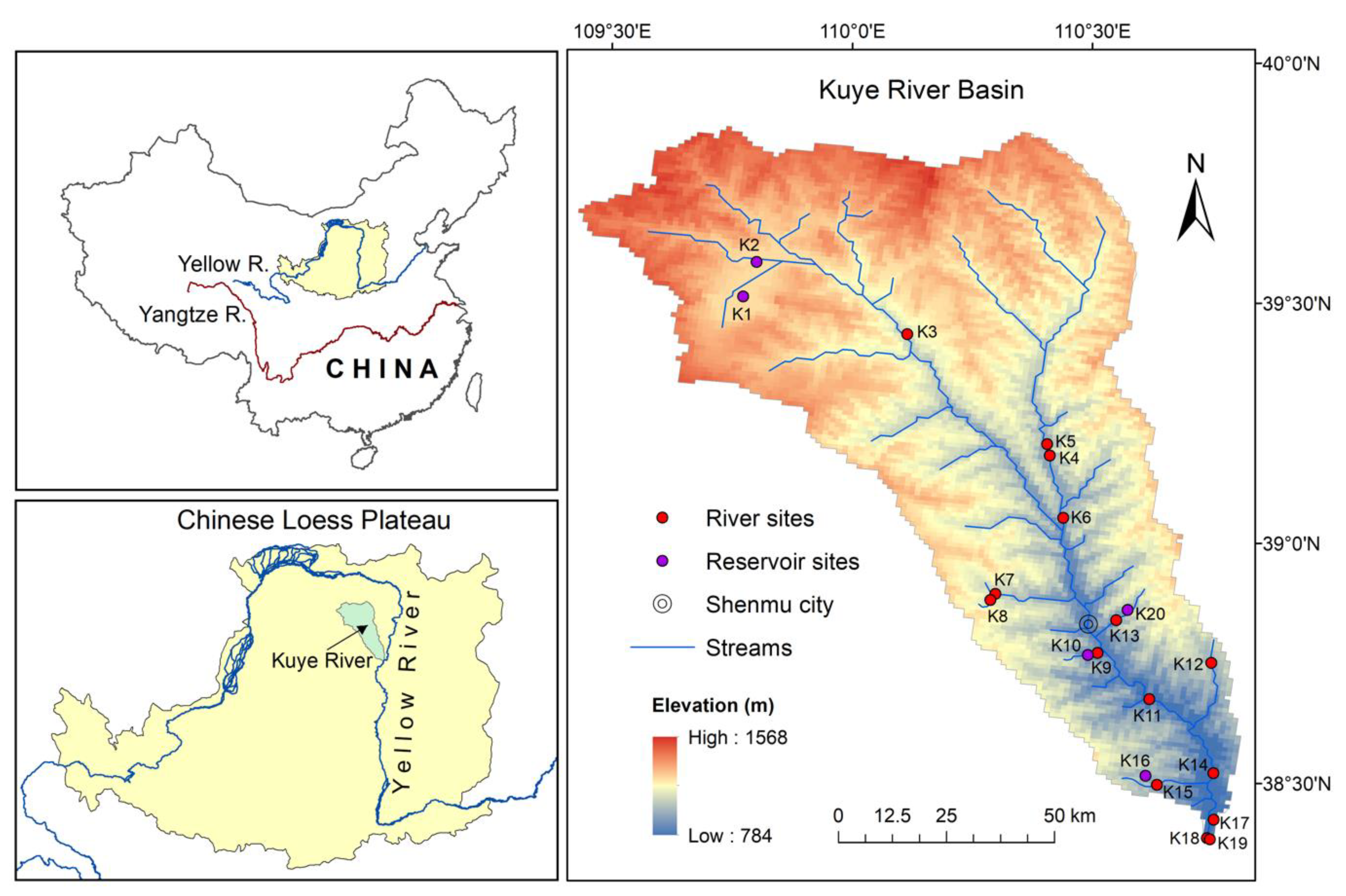
2.1. Study Site
2.2. Field Measurement of Water-Quality Parameters and CO2 and CH4 Emissions
2.3. Calculation of CO2- and CH4-Emission Flux
2.4. Statistical Analyses
3. Results
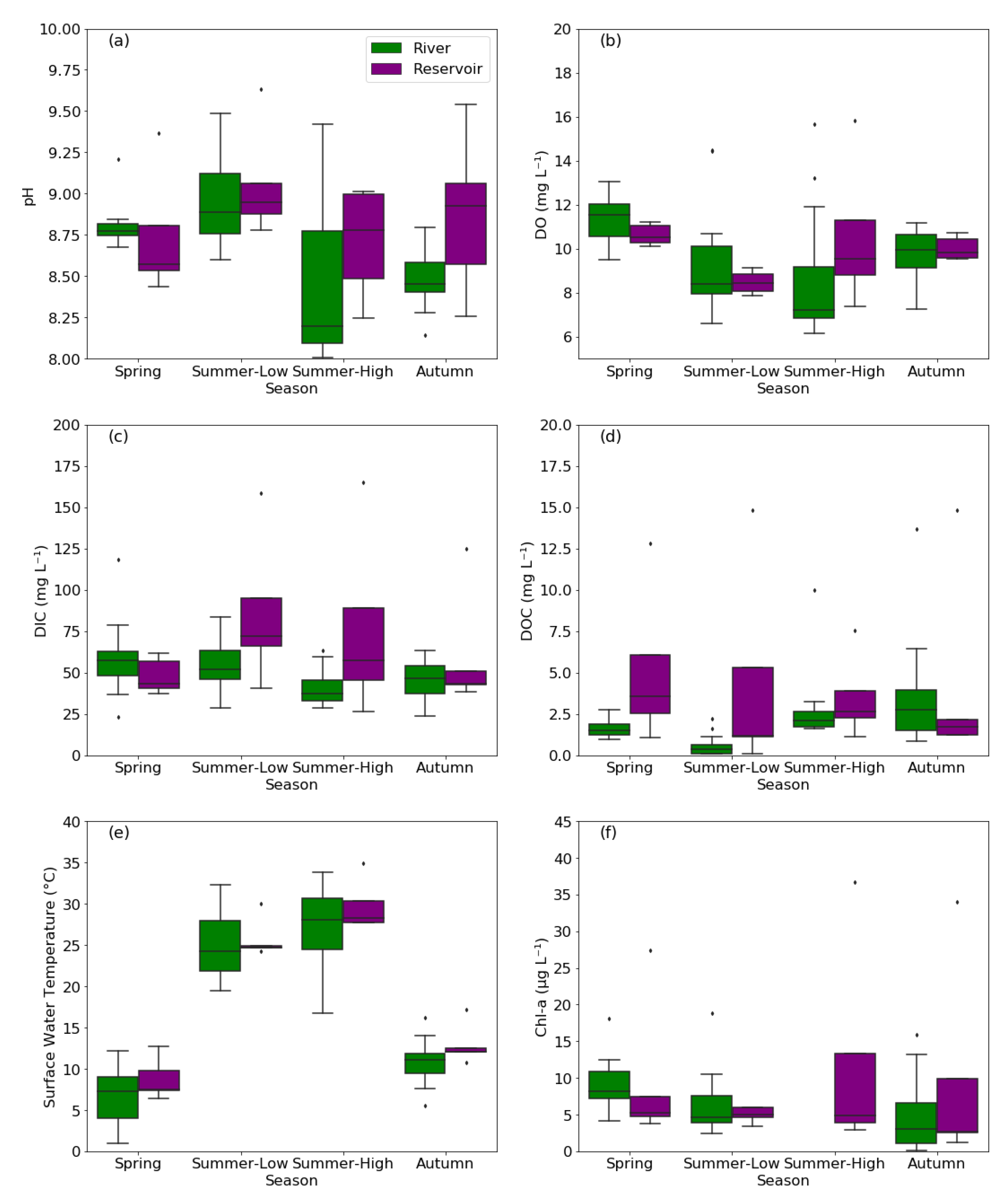
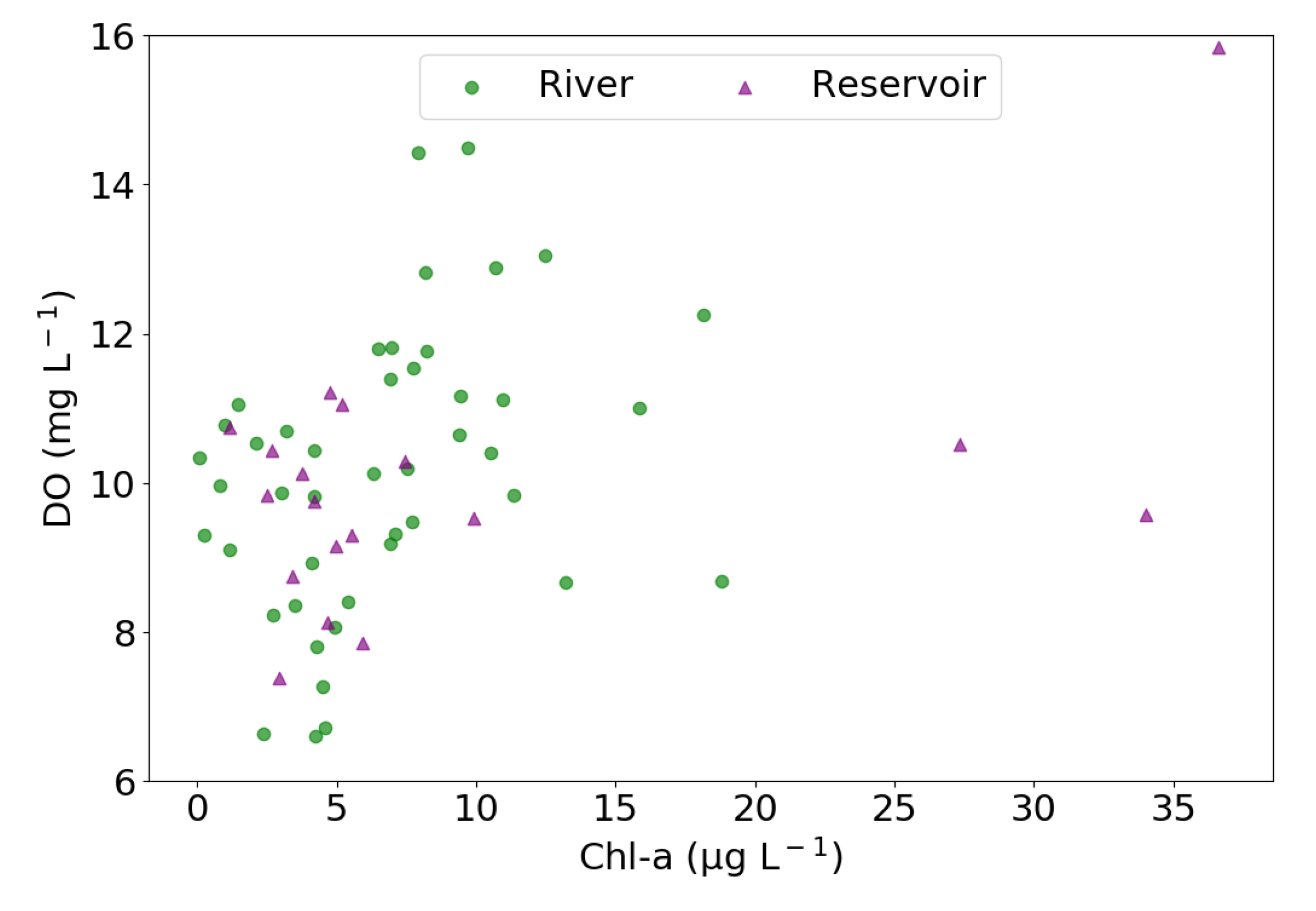
3.1. Environmental and Water-Quality Factors
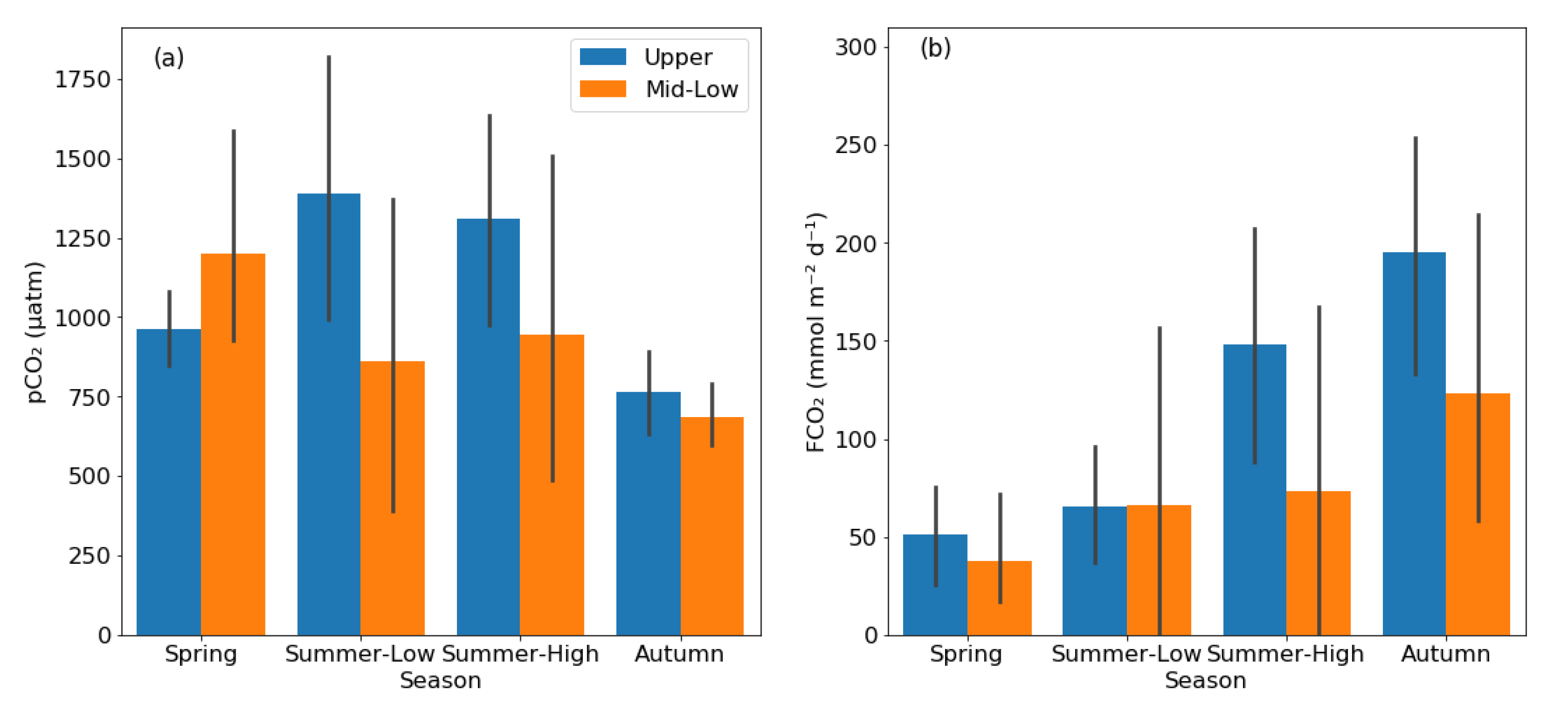
3.2. pCO2 and FCO2 in Rivers and Reservoirs
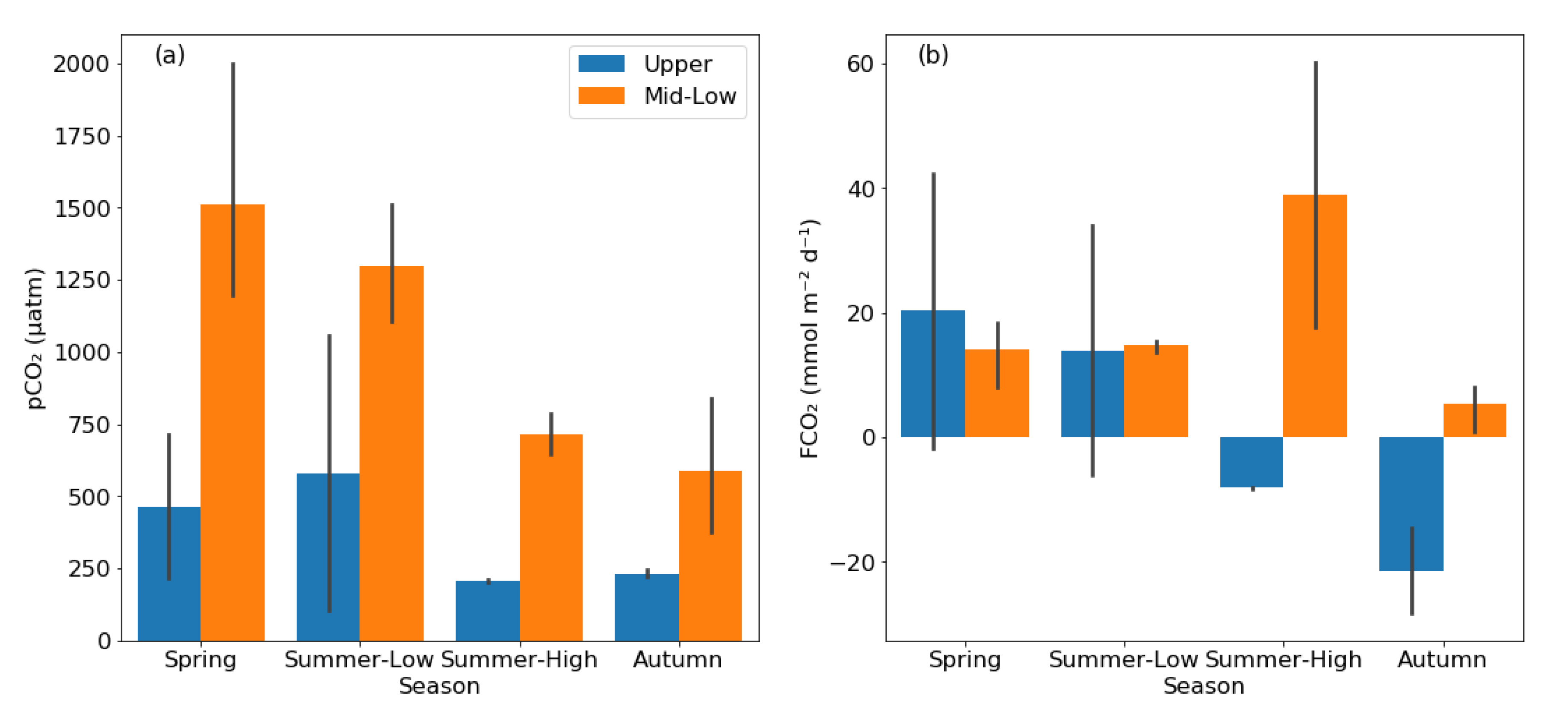
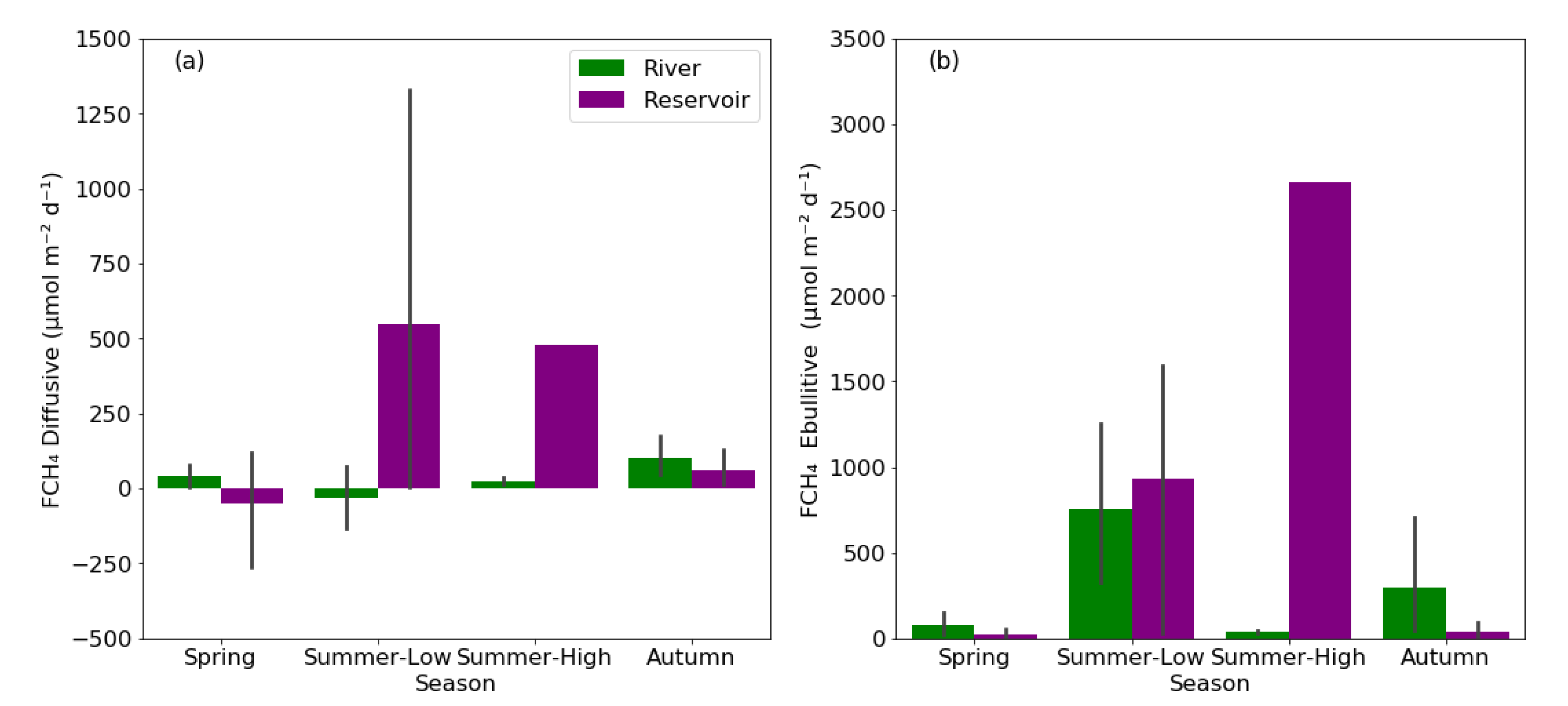
3.3. Diffusive and Ebullitive FCH4 in Rivers and Reservoirs
3.4. Estimates of pCO2 and FCH4 in Multiple Linear Regression Models
4. Discussion
4.1. Seasonal Differences of CO2 and CH4 Characteristics
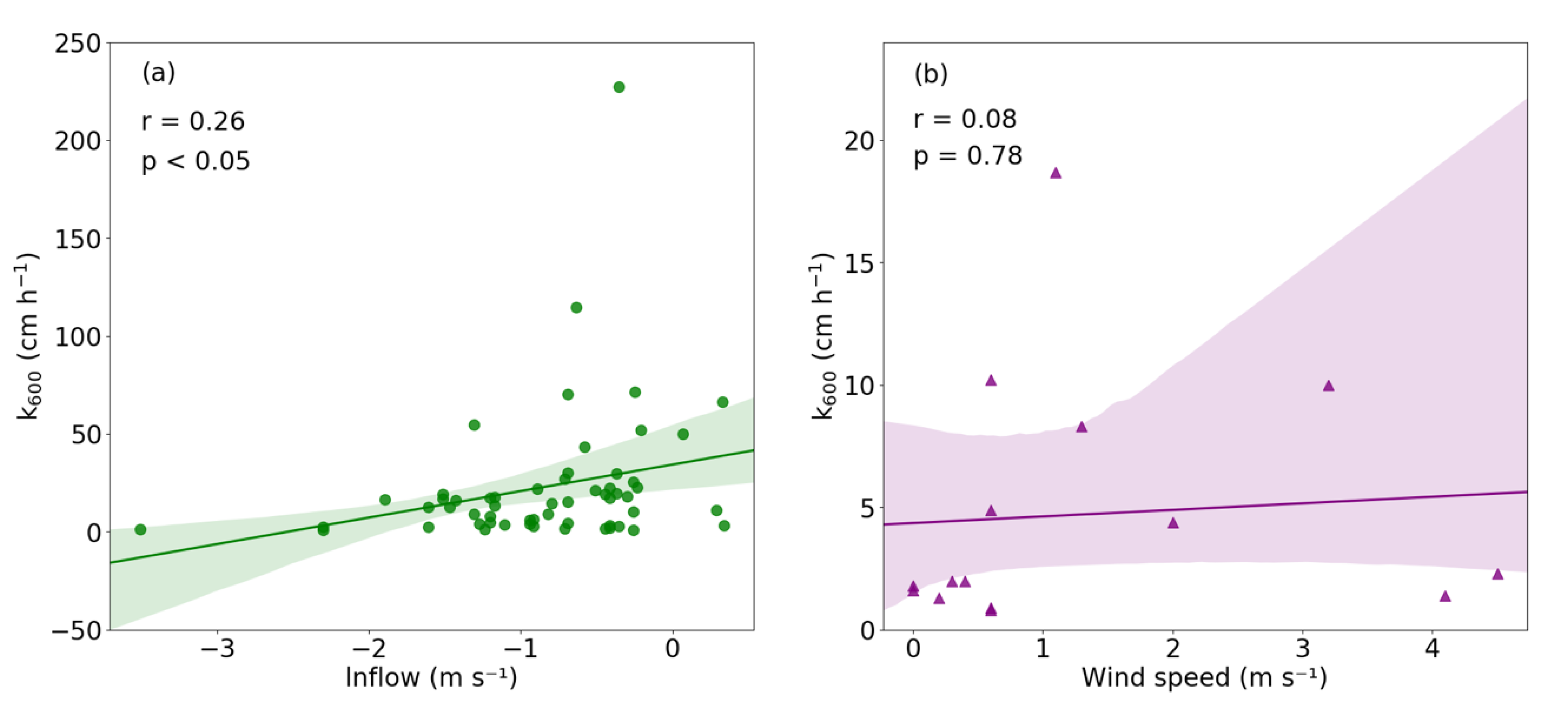
4.2. Environmental Drivers of CO2 and CH4 Emissions
4.3. Comparison of C Emissions with Other Regions and Its Implications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cole, J.J.; Prairie, Y.T.; Caraco, N.F.; McDowell, W.H.; Tranvik, L.J.; Striegl, R.G.; Duarte, C.M.; Kortelainen, P.; Downing, J.A.; Middelburg, J.J.; et al. Plumbing the Global Carbon Cycle: Integrating Inland Waters into the Terrestrial Carbon Budget. Ecosystems 2007, 10, 172–185. [Google Scholar] [CrossRef]
- Battin, T.J.; Luyssaert, S.; Kaplan, L.A.; Aufdenkampe, A.K.; Richter, A.; Tranvik, L.J. The boundless carbon cycle. Nat. Geosci. 2009, 2, 598–600. [Google Scholar] [CrossRef]
- Wallin, M.B.; Grabs, T.; Buffam, I.; Laudon, H.; Ågren, A.; Öquist, M.G.; Bishop, K. Evasion of CO2 from streams–The dominant component of the carbon export through the aquatic conduit in a boreal landscape. Glob. Chang. Biol. 2013, 19, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Raymond, P.A.; Hartmann, J.; Lauerwald, R.; Sobek, S.; McDonald, C.; Hoover, M.; Butman, D.; Striegl, R.; Mayorga, E.; Humborg, C.; et al. Global carbon dioxide emissions from inland waters. Nature 2013, 503, 355–359. [Google Scholar] [CrossRef]
- Teodoru, C.R.; del Giorgio, P.A.; Prairie, Y.T.; Camire, M. Patterns in pCO2 in boreal streams and rivers of northern Quebec, Canada. Glob. Biogeochem. Cycles 2009, 23. [Google Scholar] [CrossRef]
- Reiman, J.H.; Xu, Y.J. Diel Variability of pCO2 and CO2 Outgassing from the Lower Mississippi River: Implications for Riverine CO2 Outgassing Estimation. Water 2019, 11. [Google Scholar] [CrossRef]
- Striegl, R.G.; Dornblaser, M.M.; McDonald, C.P.; Rover, J.R.; Stets, E.G. Carbon dioxide and methane emissions from the Yukon River system. Glob. Biogeochem. Cycles 2012, 26. [Google Scholar] [CrossRef]
- Richey, J.E.; Melack, J.M.; Aufdenkampe, A.K.; Ballester, V.M.; Hess, L.L. Outgassing from Amazonian rivers and wetlands as a large tropical source of atmospheric CO2. Nature 2002, 416, 617–620. [Google Scholar] [CrossRef] [PubMed]
- AcuÑA, V.; Wolf, A.; Uehlinger, U.; Tockner, K. Temperature dependence of stream benthic respiration in an Alpine river network under global warming. Freshw. Biol. 2008, 53, 2076–2088. [Google Scholar] [CrossRef]
- Marx, A.; Dusek, J.; Jankovec, J.; Sanda, M.; Vogel, T.; van Geldern, R.; Hartmann, J.; Barth, J.A.C. A review of CO2 and associated carbon dynamics in headwater streams: A global perspective. Rev. Geophys. 2017, 55, 560–585. [Google Scholar] [CrossRef]
- Wanninkhof, R. Relationship between wind speed and gas exchange over the ocean. J. Geophys. Res. Ocean. 1992, 97, 7373–7382. [Google Scholar] [CrossRef]
- Raymond, P.A.; Zappa, C.J.; Butman, D.; Bott, T.L.; Potter, J.; Mulholland, P.; Laursen, A.E.; McDowell, W.H.; Newbold, D. Scaling the gas transfer velocity and hydraulic geometry in streams and small rivers. Limnol. Oceanogr. Fluids Environ. 2012, 2, 41–53. [Google Scholar] [CrossRef]
- Wanninkhof, R. Relationship between wind speed and gas exchange over the ocean revisited. Limnol. Oceanogr. Methods 2014, 12, 351–362. [Google Scholar] [CrossRef]
- Beaulieu, J.J.; DelSontro, T.; Downing, J.A. Eutrophication will increase methane emissions from lakes and impoundments during the 21st century. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef]
- Crawford, J.T.; Striegl, R.G.; Wickland, K.P.; Dornblaser, M.M.; Stanley, E.H. Emissions of carbon dioxide and methane from a headwater stream network of interior Alaska. J. Geophys. Res. Biogeosci. 2013, 118, 482–494. [Google Scholar] [CrossRef]
- Maeck, A.; DelSontro, T.; McGinnis, D.F.; Fischer, H.; Flury, S.; Schmidt, M.; Fietzek, P.; Lorke, A. Sediment Trapping by Dams Creates Methane Emission Hot Spots. Environ. Sci. Technol. 2013, 47, 8130–8137. [Google Scholar] [CrossRef]
- Sobek, S.; DelSontro, T.; Wongfun, N.; Wehrli, B. Extreme organic carbon burial fuels intense methane bubbling in a temperate reservoir. Geophys. Res. Lett. 2012, 39. [Google Scholar] [CrossRef]
- DelSontro, T.; Beaulieu, J.J.; Downing, J.A. Greenhouse gas emissions from lakes and impoundments: Upscaling in the face of global change. Limnol. Oceanogr. Lett. 2018, 3, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Spawn, S.A.; Dunn, S.T.; Fiske, G.J.; Natali, S.M.; Schade, J.D.; Zimov, N.S. Summer methane ebullition from a headwater catchment in Northeastern Siberia. Inland Waters 2015, 5, 224–230. [Google Scholar] [CrossRef][Green Version]
- Ran, L.; Lu, X.X.; Yang, H.; Li, L.; Yu, R.; Sun, H.; Han, J. CO2 outgassing from the Yellow River network and its implications for riverine carbon cycle. J. Geophys. Res. Biogeosci. 2015, 120, 1334–1347. [Google Scholar] [CrossRef]
- Fu, B.; Wang, S.; Liu, Y.; Liu, J.; Liang, W.; Miao, C. Hydrogeomorphic Ecosystem Responses to Natural and Anthropogenic Changes in the Loess Plateau of China. Annu. Rev. Earth Planet. Sci. 2017, 45, 223–243. [Google Scholar] [CrossRef]
- Ran, L.; Li, L.; Tian, M.; Yang, X.; Yu, R.; Zhao, J.; Wang, L.; Lu, X.X. Riverine CO2 emissions in the Wuding River catchment on the Loess Plateau: Environmental controls and dam impoundment impact. J. Geophys. Res. Biogeosci. 2017, 122, 1439–1455. [Google Scholar] [CrossRef]
- Müller, D.; Warneke, T.; Rixen, T.; Müller, M.; Jamahari, S.; Denis, N.; Mujahid, A.; Notholt, J. Lateral carbon fluxes and CO2 outgassing from a tropical peat-draining river. Biogeosci. 2015, 12, 5967–5979. [Google Scholar] [CrossRef]
- Campeau, A.; Lapierre, J.-F.; Vachon, D.; del Giorgio, P.A. Regional contribution of CO2 and CH4 fluxes from the fluvial network in a lowland boreal landscape of Québec. Glob. Biogeochem. Cycles 2014, 28, 57–69. [Google Scholar] [CrossRef]
- Hotchkiss, E.R.; Hall Jr, R.O.; Sponseller, R.A.; Butman, D.; Klaminder, J.; Laudon, H.; Rosvall, M.; Karlsson, J. Sources of and processes controlling CO2 emissions change with the size of streams and rivers. Nat. Geosci. 2015, 8, 696–699. [Google Scholar] [CrossRef]
- Denfeld, B.A.; Wallin, M.B.; Sahlée, E.; Sobek, S.; Kokic, J.; Chmiel, H.E.; Weyhenmeyer, G.A. Temporal and spatial carbon dioxide concentration patterns in a small boreal lake in relation to ice-cover dynamics. Boreal Environ. Res. 2015, 20, 679–692. [Google Scholar]
- Paquay, F.S.; Mackenzie, F.T.; Borges, A.V. Carbon dioxide dynamics in rivers and coastal waters of the “big island” of Hawaii, USA, during baseline and heavy rain conditions. Aquat. Geochem. 2007, 13, 1–18. [Google Scholar] [CrossRef]
- Brothers, S.M.; Prairie, Y.T.; del Giorgio, P.A. Benthic and pelagic sources of carbon dioxide in boreal lakes and a young reservoir (Eastmain-1) in eastern Canada. Glob. Biogeochem. Cycles 2012, 26. [Google Scholar] [CrossRef]
- Read, J.S.; Hamilton, D.P.; Desai, A.R.; Rose, K.C.; MacIntyre, S.; Lenters, J.D.; Smyth, R.L.; Hanson, P.C.; Cole, J.J.; Staehr, P.A.; et al. Lake-size dependency of wind shear and convection as controls on gas exchange. Geophys. Res. Lett. 2012, 39. [Google Scholar] [CrossRef]
- Gålfalk, M.; Bastviken, D.; Fredriksson, S.; Arneborg, L. Determination of the piston velocity for water-air interfaces using flux chambers, acoustic Doppler velocimetry, and IR imaging of the water surface. J. Geophys. Res. Biogeosci. 2013, 118, 770–782. [Google Scholar] [CrossRef]
- Cole, J.J.; Caraco, N.F. Atmospheric exchange of carbon dioxide in a low-wind oligotrophic lake measured by the addition of SF6. Limnol. Oceanogr. 1998, 43, 647–656. [Google Scholar] [CrossRef]
- Xing, Y.; Xie, P.; Yang, H.; Ni, L.; Wang, Y.; Rong, K. Methane and carbon dioxide fluxes from a shallow hypereutrophic subtropical Lake in China. Atmos. Environ. 2005, 39, 5532–5540. [Google Scholar] [CrossRef]
- Duc, N.T.; Crill, P.; Bastviken, D. Implications of temperature and sediment characteristics on methane formation and oxidation in lake sediments. Biogeochemistry 2010, 100, 185–196. [Google Scholar] [CrossRef]
- Sepulveda-Jauregui, A.; Hoyos-Santillan, J.; Martinez-Cruz, K.; Walter Anthony, K.M.; Casper, P.; Belmonte-Izquierdo, Y.; Thalasso, F. Eutrophication exacerbates the impact of climate warming on lake methane emission. Sci. Total Environ. 2018, 636, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Bastviken, D.; Cole, J.J.; Pace, M.L.; Van de Bogert, M.C. Fates of methane from different lake habitats: Connecting whole-lake budgets and CH4 emissions. J. Geophys. Res. Biogeosci. 2008, 113. [Google Scholar] [CrossRef]
- DelSontro, T.; Boutet, L.; St-Pierre, A.; del Giorgio, P.A.; Prairie, Y.T. Methane ebullition and diffusion from northern ponds and lakes regulated by the interaction between temperature and system productivity. Limnol. Oceanogr. 2016, 61, S62–S77. [Google Scholar] [CrossRef]
- Wu, S.; Li, S.; Zou, Z.; Hu, T.; Hu, Z.; Liu, S.; Zou, J. High Methane Emissions Largely Attributed to Ebullitive Fluxes from a Subtropical River Draining a Rice Paddy Watershed in China. Environ. Sci. Technol. 2019, 53, 3499–3507. [Google Scholar] [CrossRef]
- Wang, X.; He, Y.; Yuan, X.; Chen, H.; Peng, C.; Zhu, Q.; Yue, J.; Ren, H.; Deng, W.; Liu, H. pCO2 and CO2 fluxes of the metropolitan river network in relation to the urbanization of Chongqing, China. J. Geophys. Res. Biogeosci. 2017, 122, 470–486. [Google Scholar] [CrossRef]
- Zhai, W.; Dai, M.; Cai, W.-J.; Wang, Y.; Wang, Z. High partial pressure of CO2 and its maintaining mechanism in a subtropical estuary: The Pearl River estuary, China. Mar. Chem. 2005, 93, 21–32. [Google Scholar] [CrossRef]
- Duvert, C.; Bossa, M.; Tyler, K.J.; Wynn, J.G.; Munksgaard, N.C.; Bird, M.I.; Setterfield, S.A.; Hutley, L.B. Groundwater-Derived DIC and Carbonate Buffering Enhance Fluvial CO2 Evasion in Two Australian Tropical Rivers. J. Geophys. Res. Biogeosci. 2019, 124, 312–327. [Google Scholar] [CrossRef]
- Stets, E.G.; Butman, D.; McDonald, C.P.; Stackpoole, S.M.; DeGrandpre, M.D.; Striegl, R.G. Carbonate buffering and metabolic controls on carbon dioxide in rivers. Glob. Biogeochem. Cycles 2017, 31, 663–677. [Google Scholar] [CrossRef]
- Finlay, K.; Vogt, R.J.; Bogard, M.J.; Wissel, B.; Tutolo, B.M.; Simpson, G.L.; Leavitt, P.R. Decrease in CO2 efflux from northern hardwater lakes with increasing atmospheric warming. Nature 2015, 519, 215–218. [Google Scholar] [CrossRef]
- Zang, C.; Huang, S.; Wu, M.; Du, S.; Scholz, M.; Gao, F.; Lin, C.; Guo, Y.; Dong, Y. Comparison of Relationships Between pH, Dissolved Oxygen and Chlorophyll a for Aquaculture and Non-aquaculture Waters. WaterAirSoil Pollut. 2011, 219, 157–174. [Google Scholar] [CrossRef]
- Crawford, J.T.; Loken, L.C.; Stanley, E.H.; Stets, E.G.; Dornblaser, M.M.; Striegl, R.G. Basin scale controls on CO2 and CH4 emissions from the Upper Mississippi River. Geophys. Res. Lett. 2016, 43, 1973–1979. [Google Scholar] [CrossRef]
- Morales-Pineda, M.; Cózar, A.; Laiz, I.; Úbeda, B.; Gálvez, J.Á. Daily, biweekly, and seasonal temporal scales of pCO2 variability in two stratified Mediterranean reservoirs. J. Geophys. Res. Biogeosci. 2014, 119, 509–520. [Google Scholar] [CrossRef]
- Maurice, L.; Rawlins, B.G.; Farr, G.; Bell, R.; Gooddy, D.C. The Influence of Flow and Bed Slope on Gas Transfer in Steep Streams and Their Implications for Evasion of CO2. J. Geophys. Res. Biogeosci. 2017, 122, 2862–2875. [Google Scholar] [CrossRef]
- Raymond, P.A.; Cole, J.J. Gas Exchange in Rivers and Estuaries: Choosing a Gas Transfer Velocity. Estuaries 2001, 24, 312–317. [Google Scholar] [CrossRef]
- Ulseth, A.J.; Hall, R.O.; Boix Canadell, M.; Madinger, H.L.; Niayifar, A.; Battin, T.J. Distinct air–water gas exchange regimes in low- and high-energy streams. Nat. Geosci. 2019, 12, 259–263. [Google Scholar] [CrossRef]
- Beaulieu, J.J.; Shuster, W.D.; Rebholz, J.A. Controls on gas transfer velocities in a large river. J. Geophys. Res. Biogeosci. 2012, 117. [Google Scholar] [CrossRef]
- Ho, D.T.; Veron, F.; Harrison, E.; Bliven, L.F.; Scott, N.; McGillis, W.R. The combined effect of rain and wind on air–water gas exchange: A feasibility study. J. Mar. Syst. 2007, 66, 150–160. [Google Scholar] [CrossRef]
- Zappa, C.J.; Ho, D.T.; McGillis, W.R.; Banner, M.L.; Dacey, J.W.H.; Bliven, L.F.; Ma, B.; Nystuen, J. Rain-induced turbulence and air-sea gas transfer. J. Geophys. Res. Ocean. 2009, 114. [Google Scholar] [CrossRef]
- Mu, C.; Li, L.; Wu, X.; Zhang, F.; Jia, L.; Zhao, Q.; Zhang, T. Greenhouse gas released from the deep permafrost in the northern Qinghai-Tibetan Plateau. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Brothers, S.; Köhler, J.; Attermeyer, K.; Grossart, H.P.; Mehner, T.; Meyer, N.; Scharnweber, K.; Hilt, S. A feedback loop links brownification and anoxia in a temperate, shallow lake. Limnol. Oceanogr. 2014, 59, 1388–1398. [Google Scholar] [CrossRef]
- Megonigal, J.P.; Hines, M.E.; Visscher, P.T. 10.8—Anaerobic Metabolism: Linkages to Trace Gases and Aerobic Processes. In Treatise on Geochemistry, 2nd ed.; Holland, H.D., Turekian, K.K., Eds.; Elsevier: Oxford, UK, 2014; pp. 273–359. [Google Scholar]
- Haider, H.; Ali, W.; Haydar, S. Evaluation of various relationships of reaeration rate coefficient for modeling dissolved oxygen in a river with extreme flow variations in Pakistan. Hydrol. Process. 2013, 27, 3949–3963. [Google Scholar] [CrossRef]
- Tang, K.W.; McGinnis, D.F.; Frindte, K.; Brüchert, V.; Grossart, H.-P. Paradox reconsidered: Methane oversaturation in well-oxygenated lake waters. Limnol. Oceanogr. 2014, 59, 275–284. [Google Scholar] [CrossRef]
- Huotari, J.; Haapanala, S.; Pumpanen, J.; Vesala, T.; Ojala, A. Efficient gas exchange between a boreal river and the atmosphere. Geophys. Res. Lett. 2013, 40, 5683–5686. [Google Scholar] [CrossRef]
- Rasilo, T.; Hutchins, R.H.S.; Ruiz-González, C.; del Giorgio, P.A. Transport and transformation of soil-derived CO2, CH4 and DOC sustain CO2 supersaturation in small boreal streams. Sci. Total Environ. 2017, 579, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Yang, T.; Sharma, M.P. Greenhouse gas measurement from Chinese freshwater bodies: A review. J. Clean. Prod. 2019, 233, 368–378. [Google Scholar] [CrossRef]
- Li, S.; Lu, X.X.; Bush, R.T. CO2 partial pressure and CO2 emission in the Lower Mekong River. J. Hydrol. 2013, 504, 40–56. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, M.P. Assessment of risk of GHG emissions from Tehri hydropower reservoir, India. Hum. Ecol. Risk Assess. Int. J. 2016, 22, 71–85. [Google Scholar] [CrossRef]
- St. Louis, V.L.; Kelly, C.A.; Duchemin, É.; Rudd, J.W.M.; Rosenberg, D.M. Reservoir Surfaces as Sources of Greenhouse Gases to the Atmosphere: A Global Estimate: Reservoirs are sources of greenhouse gases to the atmosphere, and their surface areas have increased to the point where they should be included in global inventories of anthropogenic emissions of greenhouse gases. BioScience 2000, 50, 766–775. [Google Scholar]
- Wang, F.; Wang, B.; Liu, C.-Q.; Wang, Y.; Guan, J.; Liu, X.; Yu, Y. Carbon dioxide emission from surface water in cascade reservoirs–river system on the Maotiao River, southwest of China. Atmos. Environ. 2011, 45, 3827–3834. [Google Scholar] [CrossRef]
- Almeida, R.M.; Nóbrega, G.N.; Junger, P.C.; Figueiredo, A.V.; Andrade, A.S.; de Moura, C.G.B.; Tonetta, D.; Oliveira, E.S.; Araújo, F.; Rust, F.; et al. High Primary Production Contrasts with Intense Carbon Emission in a Eutrophic Tropical Reservoir. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Hao, Q.; Chen, S.; Ni, X.; Li, X.; He, X.; Jiang, C. Methane and nitrous oxide emissions from the drawdown areas of the Three Gorges Reservoir. Sci. Total Environ. 2019, 660, 567–576. [Google Scholar] [CrossRef] [PubMed]
- DelSontro, T.; McGinnis, D.F.; Wehrli, B.; Ostrovsky, I. Size Does Matter: Importance of Large Bubbles and Small-Scale Hot Spots for Methane Transport. Environ. Sci. Technol. 2015, 49, 1268–1276. [Google Scholar] [CrossRef] [PubMed]
- Linkhorst, A.; Hiller, C.; DelSontro, T.; Azevedo, G.M.; Barros, N.; Mendonça, R.; Sobek, S. Comparing methane ebullition variability across space and time in a Brazilian reservoir. Limnol. Oceanogr. 2020, 65, 1623–1634. [Google Scholar] [CrossRef]
- Kumar, A.; Yang, T.; Sharma, M.P. Long-term prediction of greenhouse gas risk to the Chinese hydropower reservoirs. Sci. Total Environ. 2019, 646, 300–308. [Google Scholar] [CrossRef]
- Zarfl, C.; Lumsdon, A.E.; Berlekamp, J.; Tydecks, L.; Tockner, K. A global boom in hydropower dam construction. Aquat. Sci. 2015, 77, 161–170. [Google Scholar] [CrossRef]
- Brothers, S.M.; del Giorgio, P.A.; Teodoru, C.R.; Prairie, Y.T. Landscape heterogeneity influences carbon dioxide production in a young boreal reservoir. Can. J. Fish. Aquat. Sci. 2012, 69, 447–456. [Google Scholar] [CrossRef]
- Teodoru, C.R.; Bastien, J.; Bonneville, M.-C.; del Giorgio, P.A.; Demarty, M.; Garneau, M.; Hélie, J.-F.; Pelletier, L.; Prairie, Y.T.; Roulet, N.T.; et al. The net carbon footprint of a newly created boreal hydroelectric reservoir. Glob. Biogeochem. Cycles 2012, 26. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Estimate | Std. Error | t | p-Value | |
|---|---|---|---|---|---|
| River | Intercept | 5835.841 | 1785.974 | 3.268 | 0.002 |
| pH | −524.591 | 236.221 | −2.221 | 0.031 | |
| DO | −84.801 | 36.166 | −2.345 | 0.023 | |
| DIC | 10.833 | 3.953 | 2.740 | 0.008 | |
| Reservoir | Intercept | 1217.026 | 180.097 | 6.758 | 0.000 |
| Chl-a | −16.802 | 14.624 | −1.149 | 0.273 | |
| Wind speed | −176.315 | 93.382 | −1.888 | 0.083 |
| Parameter | Estimate | Std. Error | t | p-Value | |
|---|---|---|---|---|---|
| River | Intercept | −8.188 | 3.852 | −2.126 | 0.044 |
| pH | 0.977 | 0.442 | 2.212 | 0.037 | |
| Chl-a | 0.028 | 0.024 | 1.196 | 0.244 | |
| Windspeed | −0.315 | 0.179 | −1.762 | 0.091 | |
| Reservoir | Intercept | −1.165 | 0.579 | −2.013 | 0.079 |
| Temperature | 0.106 | 0.028 | 3.752 | 0.006 |
| Water Type | Location | Climate Zone | FCO2 (mmol m−2 d−1) | FCH4 (µmol m−2 d−1) | References |
|---|---|---|---|---|---|
| Streams and rivers | Northwest China | Temperate | 91 | 352 | This study |
| China | Subtropical | 38–356 | 864–2938 | [59] | |
| Interior Alaska | Boreal | 450.1 | 631 | [15] | |
| Finland | Boreal | 83 | - | [57] | |
| Québec | Boreal | 66.3 | 2000 | [58] | |
| Chongqing | Subtropical | 447 | - | [38] | |
| Lower Mekong | Tropical | 195 | - | [60] | |
| Amazon | Tropical | 189 | - | [8] | |
| Lakes and reservoirs | Northwest China | Temperate | 10 | 779 | This study |
| New Hampshire | Boreal | 6.7–11.5 | - | [31] | |
| Brazil | Tropical | 42.5 | 2330 | [64] | |
| India | Subtropical | 18 | 4000 | [61] | |
| Switzerland | Temperate | 5.5 | 9820 | [17] | |
| China | Subtropical | 0.9–187 | 8.6–4840 | [59] | |
| Southwest China | Subtropical | 15–47.2 | - | [63] | |
| Three Gorges, China | Subtropical | - | 780 | [65] | |
| Global | Temperate | 32 | 1250 | [62] | |
| Global | Tropical | 79.5 | 18,800 | [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, C.-N.; Shi, H.; Liu, B.; Ran, L. CO2 and CH4 Emissions from an Arid Fluvial Network on the Chinese Loess Plateau. Water 2021, 13, 1614. https://doi.org/10.3390/w13121614
Chan C-N, Shi H, Liu B, Ran L. CO2 and CH4 Emissions from an Arid Fluvial Network on the Chinese Loess Plateau. Water. 2021; 13(12):1614. https://doi.org/10.3390/w13121614
Chicago/Turabian StyleChan, Chun-Ngai, Hongyan Shi, Boyi Liu, and Lishan Ran. 2021. "CO2 and CH4 Emissions from an Arid Fluvial Network on the Chinese Loess Plateau" Water 13, no. 12: 1614. https://doi.org/10.3390/w13121614
APA StyleChan, C.-N., Shi, H., Liu, B., & Ran, L. (2021). CO2 and CH4 Emissions from an Arid Fluvial Network on the Chinese Loess Plateau. Water, 13(12), 1614. https://doi.org/10.3390/w13121614