Using Chlorophyll a Fluorescence Imaging to Select Desiccation-Tolerant Native Moss Species for Water-Sustainable Green Roofs

 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Experimental Design
2.3. Imaging Pulse Amplitude Modulated Chlorophyll a Fluorescence
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oberndorfer, E.; Lundholm, J.; Bass, B.; Coffman, R.R.; Doshi, H.; Dunnett, N.; Gaffin, S.; Köhler, M.; Liu, K.K.Y.; Rowe, B. Green roofs as urban ecosystems: Ecological structures, functions, and services. BioScience 2007, 57, 823–833. [Google Scholar] [CrossRef]
- Clark, C.; Adriaens, P.; Talbot, F.B. Green roof valuation: A probabilistic economic analysis of environmental benefits. Environ. Sci. Technol. 2008, 42, 2155–2161. [Google Scholar] [CrossRef]
- Currie, B.A.; Bass, B. Estimates of air pollution mitigation with green plants and green roofs using the UFORE model. Urban Ecosyst. 2008, 11, 409–422. [Google Scholar] [CrossRef]
- Carter, T.; Keeler, A. Life-cycle cost-benefit analysis of extensive vegetated roof systems. J. Environ. Manag. 2008, 8, 350–363. [Google Scholar] [CrossRef] [PubMed]
- Rowe, D.B. Green roofs as a means of pollution abatement. Environ. Pollut. (Oxford, U.K.) 2011, 159, 2100–2110. [Google Scholar] [CrossRef] [PubMed]
- Veisten, K.; Smyrnova, Y.; Klæboe, R.; Hornikx, M.; Mosslemi, M.; Kang, J. Valuation of green walls and green roofs as soundscape measures: Including monetised amenity values together with noise-attenuation values in a cost-benefit analysis of a green wall affecting courtyards. Int. J. Environ. Res. Public Health 2012, 9, 3770–3778. [Google Scholar] [CrossRef]
- Anderson, M.; Lambrinos, J.; Schroll, E. The potential value of mosses for stormwater management in urban environments. Urban Ecosyst. 2010, 13, 319–332. [Google Scholar] [CrossRef]
- Alexandri, E.; Jones, P. Temperature decreases in an urban canyon due to green walls and green roofs in diverse climates. Build. Environ. 2008, 4, 480–493. [Google Scholar] [CrossRef]
- Berardi, U.; GhaffarianHoseini, A.; GhaffarianHoseini, A. State-of-the-art analysis of the environmental benefits of green roofs. Appl. Energy 2014, 115, 411–428. [Google Scholar] [CrossRef]
- Snodgrass, E.C.; Snodgrass, L.L. Green Roof Plants: A Resource and Planting Guide; Timber Press: Portland, OR, USA, 2006; 220p, ISBN 978-0881927870. [Google Scholar]
- Yang, J.; Yu, Q.; Gong, P. Quantifying air pollution removal by green roofs in Chicago. Atmos. Environ. 2008, 42, 7266–7273. [Google Scholar] [CrossRef]
- Loder, A. “There’s a meadow outside my workplace”: A phenomenological exploration of aesthetics and green roofs in Chicago and Toronto. Landsc. Urban Plan. 2014, 126, 94–106. [Google Scholar] [CrossRef]
- Sutton, R.K. Aesthetics for green roofs and green walls. J. Liv. Architect. 2014, 1, 1–20. Available online: https://livingarchitecturemonitor.com/s/JLIV2014Volume1_Issue2-Sutton.pdf (accessed on 1 May 2020).
- Schweitzer, O.; Erell, E. Evaluation of the energy performance and irrigation requirements of extensive green roofs in a water-scarce Mediterranean climate. Energy Build. 2014, 68, 25–32. [Google Scholar] [CrossRef]
- Sovocool, K.A.; Morgan, M. Xeriscape Conversion Study Final Report; Southern Nevada Water Authority: Las Vegas, NV, USA, 2005; p. 93. Available online: https://www.snwa.com/assets/pdf/reports-xeriscape.pdf (accessed on 1 May 2020).
- Belnap, J.; Lange, O. (Eds.) Structure and Functioning of Biological Soil Crusts: A synthesis. In Biological Soil Crusts: Structure, Function, and Management. Ecological Studies (Analysis and Synthesis) Volume 150; Springer: Berlin/Heidelberg, Germany, 2001; pp. 471–479. [Google Scholar] [CrossRef]
- Bowker, M.A.; Maestre, F.T.; Eldridge, D.; Belnap, J.; Castillo-Monroy, A.; Escolar, C.; Soliveres, S. Biological soil crusts (biocrusts) as a model system in community, landscape and ecosystem ecology. Biodivers. Conserv. 2014, 23, 1619–1637. [Google Scholar] [CrossRef]
- Cruz de Carvalho, R.C.; Branquinho, C.; Marques da Silva, J. Physiological consequences of desiccation in the aquatic bryophyte Fontinalis antipyretica. Planta 2011, 234, 195–205. [Google Scholar] [CrossRef]
- Cruz de Carvalho, R.; Catalá, M.; Marques da Silva, J.; Branquinho, C.; Barreno, E. The impact of dehydration rate on the production and cellular location of reactive oxygen species in an aquatic moss. Ann. Bot. 2012, 110, 1007–1016. [Google Scholar] [CrossRef]
- Cruz de Carvalho, R.; Bernardes da Silva, A.; Soares, R.; Almeida, A.M.; Coelho, A.V.; Marques da Silva, J.; Branquinho, C. Differential proteomics of dehydration and rehydration in bryophytes: Evidence towards a common desiccation tolerance mechanism. Plant Cell Environ. 2014, 37, 1499–1515. [Google Scholar] [CrossRef]
- Cruz de Carvalho, R.; Maurício, A.; Pereira, M.F.; Marques da Silva, J.; Branquinho, C. All for one: The role of colony morphology in bryophyte desiccation tolerance. Front. Plant Sci. 2019, 10, 1360. [Google Scholar] [CrossRef]
- Rixen, C.; Mulder, C.P.H. Improved water retention links high species richness with increased productivity in arctic tundra moss communities. Oecologia 2005, 146, 287–299. [Google Scholar] [CrossRef]
- Brandão, C.; Cameira, M.R.; Valente, F.; Cruz de Carvalho, R.; Paço, T.A. Wet season hydrological performance of green roofs using native species under Mediterranean climate. Ecol. Eng. 2017, 102, 596–611. [Google Scholar] [CrossRef]
- Paço, T.A.; Cruz de Carvalho, R.; Arsénio, P.; Martins, D. Green roof design techniques to improve water use under Mediterranean conditions. Urban Sci. 2019, 3, 14. [Google Scholar] [CrossRef]
- Schreiber, U.; Schliwa, U.; Bilger, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef]
- Oxborough, K.; Baker, N.R. An instrument capable of imaging chlorophyll a fluorescence from intact leaves at very low irradiance and at cellular and subcellular levels of organization. Plant Cell Environ. 1997, 20, 1473–1483. [Google Scholar] [CrossRef]
- Marques da Silva, J. Monitoring Photosynthesis by In Vivo Chlorophyll Fluorescence: Application to High-Throughput Plant Phenotyping. In Applied Photosynthesis—New Progress; Najafpour, M.M., Ed.; Intechopen: London, UK, 2016; pp. 3–22. [Google Scholar] [CrossRef]
- Bates, B.C.; Kundzewicz, Z.W.; Wu, S.; Palutikof, J.P. (Eds.) Climate Change and Water. Technical Paper of the Intergovernmental Panel on Climate Change; PCC Secretariat: Geneva, Switzerland, 2008; 210p, ISBN 978-92-9169-123-4. [Google Scholar]
- Cruz de Carvalho, R.; Varela, Z.; Paço, T.A.; Branquinho, C. Selecting potential moss species for green roofs in the Mediterranean Basin. Urban Sci. 2019, 3, 57. [Google Scholar] [CrossRef]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulissen, D. Zeigerwerte von Pflanzen in Mitteleuropa Scripta Geobotanica 18, 2nd ed.; Goltze: Göttingen, Germany, 1992; pp. 1–258. ISBN 9783884525180. [Google Scholar]
- Dierssen, K. Distribution, Ecological Amplitude and Phytosociological Characterization of European Bryophytes; Bryophytorum Bibliotheca 56; J. Cramer: Stuttgart, Germany, 2001; pp. 1–289. ISBN 978-3-443-62028-8. [Google Scholar]
- During, H.J. Ecological Classifications of Bryophytes and Lichens. In Bryophytes and Lichens in a Changing Environment; Bates, J.W., Farmer, A.M., Eds.; Clarendon: Oxford, UK, 1992; pp. 1–31. ISBN 978-0198542919. [Google Scholar]
- Kurz-Besson, C.B.; Lousada, J.L.; Gaspar, M.J.; Correia, I.E.; David, T.S.; Soares, P.M.M.; Cardoso, R.M.; Russo, A.; Varino, F.; Mériaux, C.; et al. Effects of recent minimum temperature and water deficit increases on Pinus pinaster radial growth and wood density in southern Portugal. Front. Plant Sci. 2016, 7, 1170. [Google Scholar] [CrossRef] [PubMed]
- British Columbia Ministry of Forests. Techniques and Procedures for Collecting, Preserving, Processing, and Storing Botanical Specimens; Victoria, B.C. Work. Pap. 18/1996; British Columbia Ministry of Forests: Victoria, BC, Canada, 1996; p. 44. Available online: https://www.for.gov.bc.ca/hfd/pubs/Docs/Wp/Wp18.pdf. (accessed on 1 May 2020).
- Baker, N.R.; Oxborough, K. Chlorophyll Fluorescence as a Probe of Photosynthetic Productivity. In Chlorophyll a Fluorescence. Advances in Photosynthesis and Respiration, Volume 19; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 65–82. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Relationships among violaxanthin deepoxidation, thylakoid membrane conformation, and nonphotochemical chlorophyll fluorescence quenching in leaves of cotton (Gossypium hirsutum L.). Planta 1994, 193, 238–246. [Google Scholar] [CrossRef]
- Oxborough, K. Imaging of chlorophyll a fluorescence: Theoretical and practical aspects of an emerging technique for the monitoring of photosynthetic performance. J. Exp. Bot. 2004, 55, 1195–1205. [Google Scholar] [CrossRef]
- Platt, T.; Gallegos, C.L.; Harrison, W.G. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton. J. Mar. Res. 1980, 38, 687–701. Available online: http://images.peabody.yale.edu/publications/jmr/jmr38-04-06.pdf (accessed on 1 May 2020).
- Hájek, T. Physiological Ecology of Peatland Bryophytes. In Photosynthesis in Bryophytes and Early Land Plants. Advances in Photosynthesis and Respiration, Chapter 13; Hanson, D.T., Rice, S.K., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 233–252. [Google Scholar] [CrossRef]
- Garabito, D.; Vallejo, R.; Montero, E.; Garabito, J.; Martínez-Abaigar, J. Green buildings envelopes with bryophytes. A review of the state of the art. Bol. Soc. Españ. Briol. 2017, 48–49, 1–16. Available online: https://www.briologia.es/boletines/vol48-49.pdf (accessed on 1 May 2020).
- Oliver, M.J.; Velten, J.; Mishler, B.D. Desiccation tolerance in bryophytes: A reflection of the primitive strategy for plant survival in dehydrating habitats? Integr. Comp. Biol. 2005, 45, 788–799. [Google Scholar] [CrossRef]
- Wang, X.Q.; Yang, P.F.; Liu, Z.; Liu, W.Z.; Hu, Y.; Chen, H.; Kuang, T.Y.; Pei, Z.M.; Shen, S.H.; He, Y.K. Exploring the mechanism of Physcomitrella patens desiccation tolerance through a proteomic strategy. Plant Physiol. 2009, 149, 1739–1750. [Google Scholar] [CrossRef] [PubMed]
- Stark, L.R. Ecology of desiccation tolerance in bryophytes: A conceptual framework and methodology. Bryologist 2017, 120, 129–164. [Google Scholar] [CrossRef]
- Cruz de Carvalho, R.; Santos, P.; Branquinho, C. Production of moss-dominated biocrusts to enhance the stability and function of the margins of artificial water bodies. Restor. Ecol. 2018, 26, 419–421. [Google Scholar] [CrossRef]
- Francis, L.F.M.; Jensen, M.B. Benefits of green roofs: A systematic review of the evidence for three ecosystem services. Urban Urban Green 2017, 28, 167–176. [Google Scholar] [CrossRef]



{kind=link}
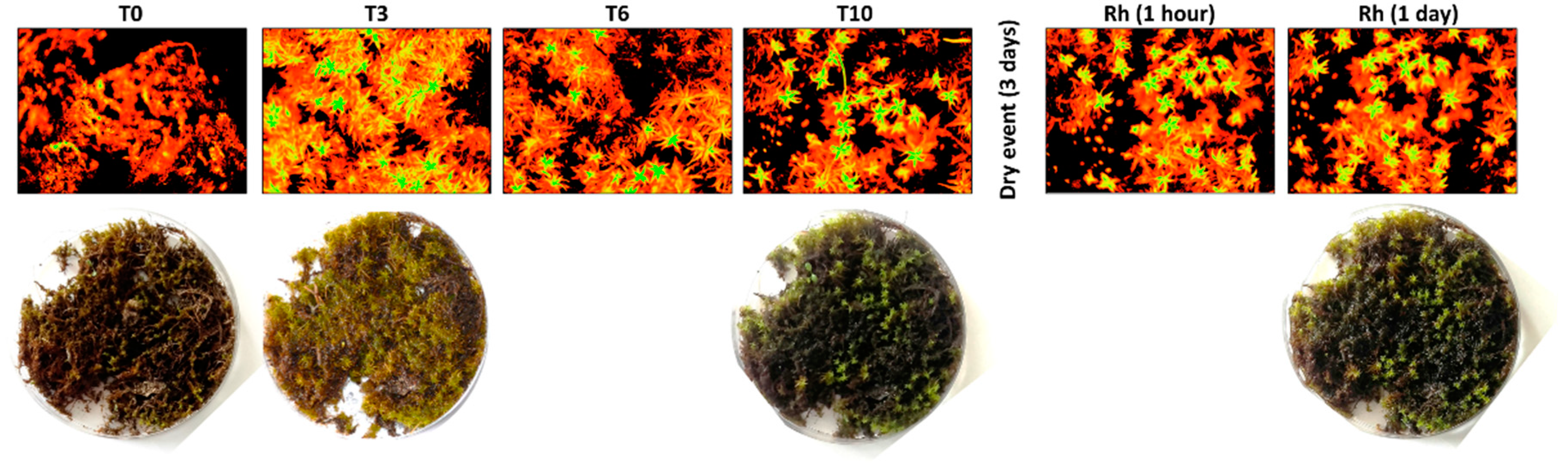{kind=link}
| Species | Plant Clade | Growth Form | Location | A.I. * | N/U | Coordinates |
|---|---|---|---|---|---|---|
| Didymodon fallax (Hedw.) R.H.Zander | Bryophyta (mosses) | Acrocarpous | Ermidas-Sado | Semi-arid | U | 38°00′24.6″ N 8°25′03.2″ W |
| Barreiro | Dry sub-humid | U | 38°39′56.0″ N 9°04′05.9″ W | |||
| Grimmia lisae De Not. | Bryophyta (mosses) | Acrocarpous | Zebreira | Semi-arid | U | 39°50′33.4″ N 7°04′07.2″ W |
| Hypnum cupressiforme Hedw. | Bryophyta (mosses) | Pleurocarpous | Alegrete (Parque Natural de São Mamede) | Dry sub-humid | N | 39°15′14.6″ N 7°18′05.0″ W |
| Pleurochaete squarrosa (Brid.) Lindb. | Bryophyta (mosses) | Acrocarpous | Zebreira | Semi-arid | U | 39°51′06.9″ N 7°04′22.9″ W |
| Targionia hypophylla L. | Marchantiophyta (liverworts) | – | Barreiro | Dry sub-humid | U | 38°39′56.0″ N 9°04′05.9″ W |
| Tortella nitida (Lindb.) Broth. | Bryophyta (mosses) | Acrocarpous | Alegrete (Parque Natural de São Mamede) | Dry sub-humid | N | 39°15′14.6″ N 7°18′05.0″ W |
| Estremoz | Semi-arid | U | 38°48′01.8″ N 7°39′41.9″ W |
| A | |||||||
| Fv/Fm | T0 | T3 | T6 | T10 | T13 | T13 + 1h | T14 |
| Didymodon fallax (E-S) | No photosynthetic activity measured by chlorophyll a fluorescence | 0.570 ± 0.026 a | 0.605 ± 0.016 a | 0.680 ± 0.017 b | No photosynthetic activity measured by chlorophyll a fluorescence | 0.589 ± 0.025 a | 0.569 ± 0.026 a |
| Didymodon fallax (Brr) | 0.626 ± 0.058 a | 0.628 ± 0.016 a | 0.545 ± 0.062 a | 0.367 ± 0.101 b | 0.565 ± 0.036 a | ||
| Grimmia lisae (Z) | 0.583 ± 0.051 ac | 0.665 ± 0.013 b | 0.625 ± 0.026 abc | 0.606 ± 0.018 c | 0.623 ± 0.022 abc | ||
| Hypnum cupressiforme (SM) | 0.281 ± 0.051 a | 0.592 ± 0.048 b | 0.655 ± 0.014 b | 0.450 ± 0.048 c | 0.620 ± 0.022 b | ||
| Pleurochaete squarrosa (Z) | 0.618 ± 0.015 a | 0.641 ± 0.019 b | 0.690 ± 0.002 c | 0.527 ± 0.004 d | 0.633 ± 0.004 ab | ||
| Targionia hypophylla (Brr) | 0.716 ± 0.008 ac | 0.744 ± 0.007 a | 0.747 ± 0.002 a | 0.666 ± 0.047 bc | 0.700 ± 0.015 c | ||
| Tortella nitida (SM) | 0.520 ± 0.030 ac | 0.613 ± 0.014 b | 0.615 ± 0.025 b | 0.491 ± 0.040 c | 0.584 ± 0.013 b | ||
| Tortella nitida (E) | 0.352 ± 0.145 a | 0.645 ± 0.047 b | 0.714 ± 0.017 b | 0.000 ± 0.000 c | 0.572 ± 0.016 d | ||
| B | |||||||
| ETR (µmol m−2 s−1) | T0 | T3 | T6 | T10 | T13 | T13 + 1h | T14 |
| Didymodon fallax (E-S) | No photosynthetic activity measured by chlorophyll a fluorescence | 18.5 ± 1.7 a | 20.2 ± 1.4 a | 17.3 ± 1.9 ab | No photosynthetic activity measured by chlorophyll a fluorescence | 13.9 ± 2.5 b | 14.0 ± 1.8 b |
| Didymodon fallax (Brr) | 20.3 ± 3.6 a | 18.2 ± 2.7 ab | 13.2 ± 1.7 bc | 8.0 ± 5.3 c | 18.7 ± 1.2 ab | ||
| Grimmia lisae (Z) | 21.1 ± 2.5 ab | 20.9 ± 1.4 a | 14.7 ± 1.2 b | 19.1 ± 3.5 a | 19.7 ± 1.7 a | ||
| Hypnum cupressiforme (SM) | 3.0 ± 2.3 a | 13.1 ± 2.8 bc | 16.1 ± 3.2 b | 10.5 ± 1.7 c | 16.6 ± 1.9 b | ||
| Pleurochaete squarrosa (Z) | 15.5 ± 1.2 a | 19.2 ± 1.0 b | 17.6 ± 0.8 c | 13.7 ± 0.3 d | 14.5 ± 0.7 ad | ||
| Targionia hypophylla (Brr) | 18.6 ± 2.3 a | 36.0 ± 0.6 b | 16.1 ± 0.4 ac | 14.3 ± 1.9 c | 17.8 ± 0.9 a | ||
| Tortella nitida (SM) | 16.1 ± 1.0 a | 14.1 ± 0.6 a | 15.1 ± 1.6 a | 12.2 ± 2.9 b | 17.0 ± 1.1 a | ||
| Tortella nitida (E) | 17.2 ± 1.5 a | 18.7 ± 2.2 a | 18.8 ± 2.8 a | 0.0 ± 0.0 b | 14.0 ± 0.7 a | ||
| C | |||||||
| NPQ/4 | T0 | T3 | T6 | T10 | T13 | T13 + 1h | T14 |
| Didymodon fallax (E-S) | No photosynthetic activity measured by chlorophyll a fluorescence | 0.132 ± 0.027 a | 0.091 ± 0.024 a | 0.236 ± 0.056 b | No photosynthetic activity measured by chlorophyll a fluorescence | 0.402 ± 0.081 c | 0.265 ± 0.037 b |
| Didymodon fallax (Brr) | 0.145 ± 0.032 ac | 0.118 ± 0.014 a | 0.121 ± 0.027 a | 0.315 ± 0.062 b | 0.194 ± 0.033 c | ||
| Grimmia lisae (Z) | 0.141 ± 0.018 ab | 0.168 ± 0.021 ab | 0.108 ± 0.034 a | 0.200 ± 0.051 b | 0.198 ± 0.028 b | ||
| Hypnum cupressiforme (SM) | 0.060 ± 0.012 a | 0.383 ± 0.096 b | 0.383 ± 0.148 b | 0.186 ± 0.037 a | 0.398 ± 0.096 b | ||
| Pleurochaete squarrosa (Z) | 0.439 ± 0.057 ac | 0.278 ± 0.053 b | 0.309 ± 0.037 bc | 0.252 ± 0.022 b | 0.376 ± 0.024 c | ||
| Targionia hypophylla (Brr) | 0.663 ± 0.098 a | 0.073 ± 0.002 b | 0.490 ± 0.059 cd | 0.508 ± 0.110 c | 0.366 ± 0.032 d | ||
| Tortella nitida (SM) | 0.147 ± 0.011 a | 0.365 ± 0.031 b | 0.296 ± 0.039 b | 0.297 ± 0.065 b | 0.318 ± 0.079 b | ||
| Tortella nitida (E) | 0.071 ± 0.049 a | 0.182 ± 0.046 b | 0.248 ± 0.089 b | 0.000 ± 0.000 c | 0.202 ± 0.023 b | ||
| A | |||||||
| α | T0 | T3 | T6 | T10 | T13 | T13 + 1h | T14 |
| Didymodon fallax (E-S) | No photosynthetic activity measured by chlorophyll a fluorescence | 0.205 ± 0.014 a | 0.246 ± 0.016 b | 0.255 ± 0.024 b | No photosynthetic activity measured by chlorophyll a fluorescence | 0.249 ± 0.023 b | 0.241 ± 0.022 ab |
| Didymodon fallax (Brr) | 0.243 ± 0.034 ac | 0.218 ± 0.012 ac | 0.170 ± 0.017 ab | 0.160 ± 0.022 b | 0.261 ± 0.037 c | ||
| Grimmia lisae (Z) | 0.241 ± 0.022 a | 0.287 ± 0.017 b | 0.244 ± 0.009 a | 0.272 ± 0.030 ab | 0.251 ± 0.014 ab | ||
| Hypnum cupressiforme (SM) | 0.104 ± 0.014 a | 0.230 ± 0.027 bc | 0.244 ± 0.049 bc | 0.194 ± 0.036 b | 0.282 ± 0.024 c | ||
| Pleurochaete squarrosa (Z) | 0.206 ± 0.025 a | 0.255 ± 0.020 b | 0.259 ± 0.020 b | 0.200 ± 0.007 a | 0.183 ± 0.002 a | ||
| Targionia hypophylla (Brr) | 0.284 ± 0.014 a | 0.285 ± 0.021 a | 0.257 ± 0.013 ac | 0.150 ± 0.006 b | 0.247 ± 0.016 c | ||
| Tortella nitida (SM) | 0.182 ± 0.014 ab | 0.185 ± 0.006 ab | 0.213 ± 0.027 a | 0.175 ± 0.015 b | 0.254 ± 0.019 c | ||
| Tortella nitida (E) | 0.178 ± 0.058 a | 0.240 ± 0.020 b | 0.251 ± 0.045 b | 0.000 ± 0.000 c | 0.192 ± 0.013 ab | ||
| B | |||||||
| β | T0 | T3 | T6 | T10 | T13 | T13 + 1h | T14 |
| Didymodon fallax (E-S) | No photosynthetic activity measured by chlorophyll a fluorescence | 57.9 ± 13.4 a | 59.1 ± 9.5 a | 102.1 ± 20.9 a | No photosynthetic activity measured by chlorophyll a fluorescence | 205.1 ± 82.6 b | 106.7 ± 17.4 a |
| Didymodon fallax (Brr) | 60.0 ± 18.6 a | 64.0 ± 16.2 a | 86.1 ± 13.3 a | 121.0 ± 42.2 a | 65.9 ± 8.3 a | ||
| Grimmia lisae (Z) | 46.9 ± 4.3 ac | 71.0 ± 16.5 abc | 89.5 ± 3.4 b | 89.9 ± 27.3 b | 62.4 ± 1.8 c | ||
| Hypnum cupressiforme (SM) | 448.6 ± 296.2 a | 155.1 ± 69.0 b | 123.6 ± 15.0 b | 195.6 ± 33.4 ab | 158.4 ± 29.8 b | ||
| Pleurochaete squarrosa (Z) | 86.4 ± 3.6 a | 88.0 ± 10.1 a | 120.5 ± 15.2 b | 128.6 ± 1.0 b | 113.3 ± 1.0 b | ||
| Targionia hypophylla (Brr) | 98.8 ± 19.2 a | 79.7 ± 11.9 a | 123.2 ± 13.2 a | 262.0 ± 100.9 b | 116.1 ± 17.0 a | ||
| Tortella nitida (SM) | 79.6 ± 2.7 a | 97.3 ± 0.7 ab | 98.3 ± 14.2 ab | 185.8 ± 120.3 b | 112.4 ± 18.9 ab | ||
| Tortella nitida (E) | 86.1 ± 28.6 a | 143.6 ± 15.0 b | 143.6 ± 15.0 b | 0.0 ± 0.0 c | 126.5 ± 4.5 b | ||
| C | |||||||
| rETRmax (µmol m−2 s−1) | T0 | T3 | T6 | T10 | T13 | T13 + 1h | T14 |
| Didymodon fallax (E-S) | No photosynthetic activity measured by chlorophyll a fluorescence | 37.6 ± 9.0 ab | 43.1 ± 6.4 a | 26.5 ± 6.7 b | No photosynthetic activity measured by chlorophyll a fluorescence | 14.0 ± 5.9 b | 23.6 ± 4.7 b |
| Didymodon fallax (Brr) | 44.9 ± 15.8 a | 37.5 ± 14.2 ab | 20.7 ± 5.1 bc | 14.9 ± 6.7 c | 40.5 ± 4.9 a | ||
| Grimmia lisae (Z) | 53.0 ± 9.2 a | 42.9 ± 9.4 ab | 27.8 ± 0.6 b | 33.9 ± 13.4 b | 40.9 ± 2.4 ab | ||
| Hypnum cupressiforme (SM) | 3.2 ± 1.8 a | 18.2 ± 9.4 bc | 20.7 ± 6.5 b | 10.3 ± 2.3 c | 18.6 ± 3.1 bc | ||
| Pleurochaete squarrosa (Z) | 24.5 ± 3.9 a | 29.7 ± 3.0 b | 22.2 ± 2.9 a | 15.9 ± 0.7 c | 16.4 ± 0.3 c | ||
| Targionia hypophylla (Brr) | 30.1 ± 5.4 ad | 37.0 ± 6.4 a | 21.5 ± 2.0 b | 6.5 ± 2.1 c | 22.1 ± 4.1 bd | ||
| Tortella nitida (SM) | 23.3 ± 1.9 a | 19.4 ± 0.7 a | 22.6 ± 5.0 a | 11.9 ± 4.6 b | 23.6 ± 4.3 a | ||
| Tortella nitida (E) | 21.3 ± 5.0 a | 17.3 ± 3.4 a | 19.7 ± 8.6 a | 0.0 ± 0.0 b | 15.5 ± 1.4 a | ||
| D | |||||||
| Ek | T0 | T3 | T6 | T10 | T13 | T13+1h | T14 |
| Didymodon fallax (E-S) | No photosynthetic activity measured by chlorophyll a fluorescence | 183.5 ± 39.4 a | 175.7 ± 26.2 a | 103.1 ± 19.8 bc | No photosynthetic activity measured by chlorophyll a fluorescence | 55.9 ± 19.9 c | 97.7 ± 16.9 c |
| Didymodon fallax (Brr) | 182.2 ± 50.6 a | 171.0 ± 59.9 a | 120.8 ± 19.1 ab | 91.1 ± 29.4 b | 156.3 ± 17.2 a | ||
| Grimmia lisae (Z) | 218.5 ± 19.2 a | 151.3 ± 43.0 bc | 114.0 ± 4.2 c | 121.8 ± 35.8 bc | 163.3 ± 4.6 b | ||
| Hypnum cupressiforme (SM) | 31.3 ± 18.8 a | 76.5 ± 31.4 b | 83.7 ± 11.8 b | 53.2 ±7.7 ab | 66.9 ± 16.8 ab | ||
| Pleurochaete squarrosa (Z) | 118.2 ± 5.1 a | 117.1 ± 13.0 a | 85.8 ± 11.4 bc | 79.4 ± 0.6 c | 90.0 ± 0.8 c | ||
| Targionia hypophylla (Brr) | 105.8 ± 16.8 ab | 129.8 ± 17.0 a | 83.5 ± 8.6 b | 43.0 ± 13.8 c | 89.3 ± 12.8 b | ||
| Tortella nitida (SM) | 128.2 ± 4.2 a | 104.8 ± 0.8 ac | 105.2 ± 12.9 ac | 67.0 ± 23.4 b | 93.0 ± 16.6 bc | ||
| Tortella nitida (E) | 128.7 ± 42.2 a | 71.7 ± 8.1 a | 75.5 ± 20.7 a | 0.0 ± 0.0 b | 80.7 ± 2.8 a | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz de Carvalho, R.; Afonso do Paço, T.; Branquinho, C.; Marques da Silva, J. Using Chlorophyll a Fluorescence Imaging to Select Desiccation-Tolerant Native Moss Species for Water-Sustainable Green Roofs. Water 2020, 12, 1748. https://doi.org/10.3390/w12061748
Cruz de Carvalho R, Afonso do Paço T, Branquinho C, Marques da Silva J. Using Chlorophyll a Fluorescence Imaging to Select Desiccation-Tolerant Native Moss Species for Water-Sustainable Green Roofs. Water. 2020; 12(6):1748. https://doi.org/10.3390/w12061748
Chicago/Turabian StyleCruz de Carvalho, Ricardo, Teresa Afonso do Paço, Cristina Branquinho, and Jorge Marques da Silva. 2020. "Using Chlorophyll a Fluorescence Imaging to Select Desiccation-Tolerant Native Moss Species for Water-Sustainable Green Roofs" Water 12, no. 6: 1748. https://doi.org/10.3390/w12061748
APA StyleCruz de Carvalho, R., Afonso do Paço, T., Branquinho, C., & Marques da Silva, J. (2020). Using Chlorophyll a Fluorescence Imaging to Select Desiccation-Tolerant Native Moss Species for Water-Sustainable Green Roofs. Water, 12(6), 1748. https://doi.org/10.3390/w12061748