Impact of Nanoscale Magnetite and Zero Valent Iron on the Batch-Wise Anaerobic Co-Digestion of Food Waste and Waste-Activated Sludge

 , , ,
, , ,
Abstract
1. Introduction
- (i)
- (ii)
- (iii)
- NZVIs serving as a conductive material to promote DIET [21].
2. Materials and Methods
2.1. Substrates and Inoculum
2.2. Preparation and Characteristics of the Nanoparticles
2.3. Anaerobic Co-Digestion Batch Tests
2.4. Analytical Methods
3. Calculations
3.1. Theoretical Biochemical Methane Potential
3.2. Theoretical COD
3.3. Experimental Biochemical Methane Potential
3.4. Biodegradability, Hydrolysis and Acidification Percentages
3.5. Statistical Analysis
3.6. Modeling of Methane Production
4. Results and Discussions
4.1. Characteristics of Substrates
4.2. Effects of Magnetite NPs and NZVIs on Hydrolysis and Acidification
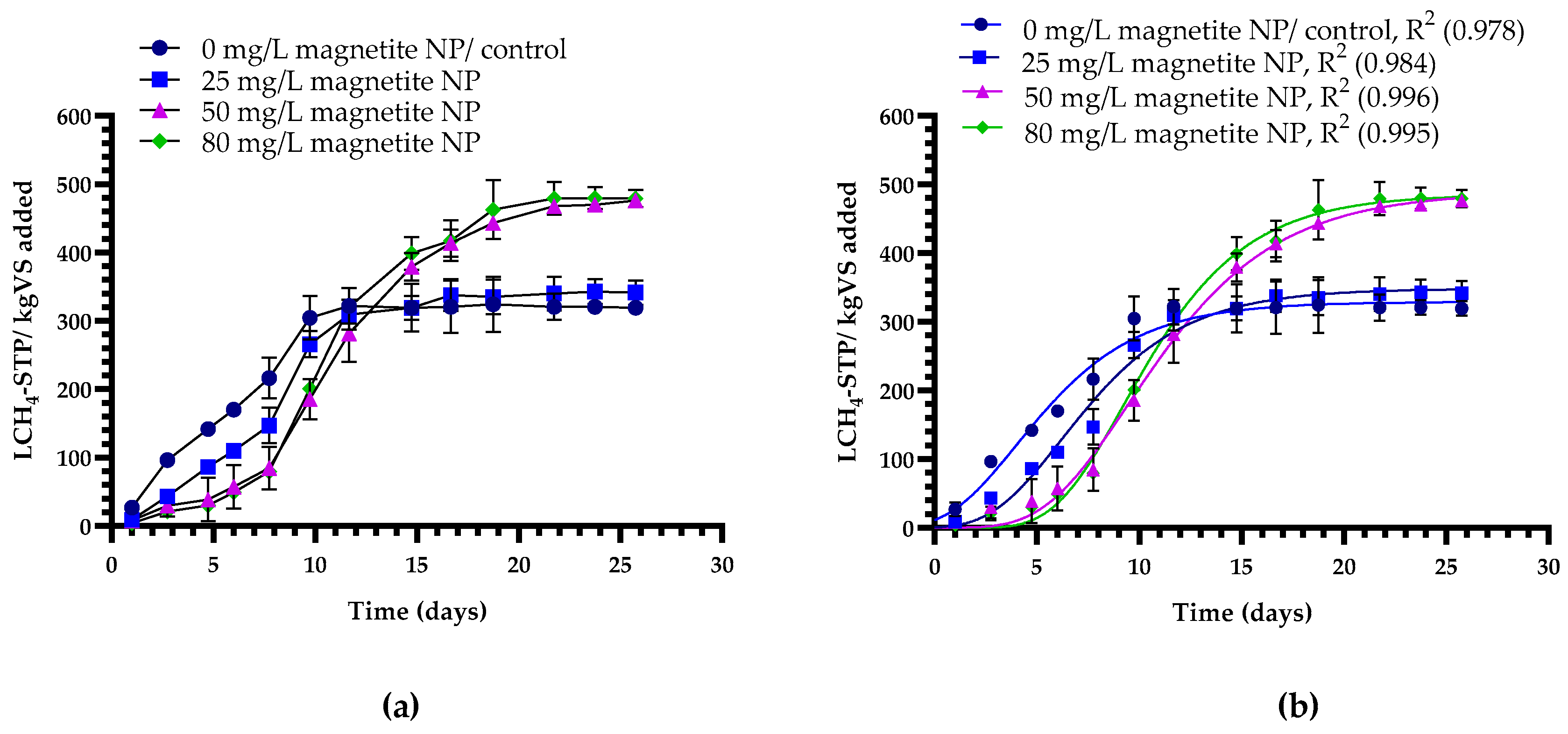
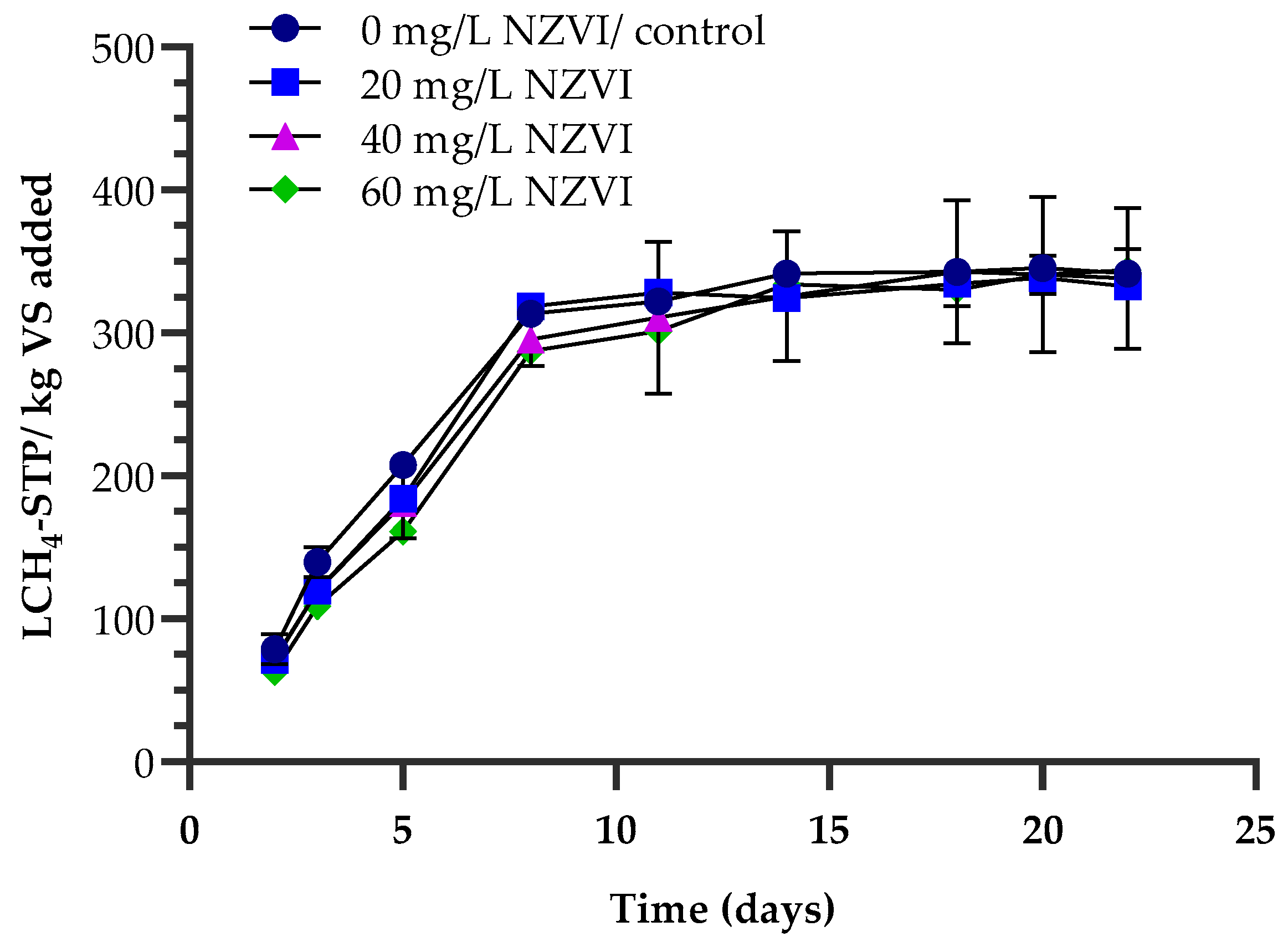
4.3. Effects of Magnetite NPs and NZVIs on Methane Production
5. Economic and Environmental Considerations
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lacovidou, E.; Ohandja, D.; Voulvoulis, N. Food waste co-digestion with sewage sludge-Realizing potential in the UK. J. Environ. Manag. 2012, 112, 267–274. [Google Scholar] [CrossRef]
- Mehariya, S.; Patel, A.; Obulisamy, P.; Punniyakotti, E. Co-digestion of food waste and sewage sludge for methane production: Current status and perspectives. Bioresour. Technol. 2018, 265, 519–531. [Google Scholar] [CrossRef]
- Kim, M.; Chowdhury, M.; Nakhla, G.; Keleman, M. Synergism of co-digestion of food wastes with municipal wastewater treatment biosolids. Waste Manag. 2017, 61, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Frear, C.; Wang, Z.; Yu, L.; Zhao, Q.; Li, X.; Chen, S. A simple methodology for rate limiting step determination for anaerobic digestion of complex substrates and effect of microbial community ratio. Bioresour. Technol. 2013, 134, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, C.; Hu, Z. Impact of metallic and metal oxide nanoparticles on wastewater treatment and anaerobic digestion. Environ. Sci. Process Impacts 2013, 15, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Lee, D. Impact of adding metal nanoparticles on anaerobic digestion performance-A review. Bioresour. Technol. 2019, 121926, 1–9. [Google Scholar] [CrossRef]
- Park, J.; Kang, H.; Park, K.; Park, H. Direct interspecies electron transfer via conductive materials: A perspective for anaerobic digestion applications. Bioresour. Technol. 2018, 254, 300–311. [Google Scholar] [CrossRef]
- Sekoai, P.; Ouma, C.; du Preez, S.; Modisha, P.; Engelbrecht, N.; Bessarabov, D.; Ghimire, A. Application of nanoparticles in biofuels: An overview. Fuel 2019, 237, 380–397. [Google Scholar] [CrossRef]
- Baek, G.; Kim, J.; Kim, J.; Lee, C. Role and potential of direct interspecies electron transfer in anaerobic digestion. Energies 2018, 11, 107. [Google Scholar] [CrossRef]
- Xu, H.; Chang, J.; Wang, H.; Liu, Y.; Zhang, X.; Liang, P.; Huang, X. Enhancing direct interspecies electron transfer in syntrophic methanogenic association with semi conductive iron oxides: Effects and mechanisms. Sci. Total Environ. 2019, 695, 133876. [Google Scholar] [CrossRef]
- Jiang, S.; Park, S.; Yoon, Y.; Lee, J.; Wu, W.; Phuoc Dan, N. Methanogenesis facilitated by geo-biochemical iron cycle in a novel syntrophic methanogenic microbial community. Environ. Sci. Technol. 2013, 47, 10078–10084. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Hashimoto, K.; Watanabe, K. Methanogenesis facilitated by electric syntrophy via (semi)conductive iron oxide minerals. Environ. Microbiol. 2012, 14, 1646–1654. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Usman, M.; Tsang, D.; O’thong, S.; Angelidaki, I.; Zhu, X.; Zhang, S.; Luo, G. Hydrochar- facilitated anaerobic digestion: Evidence for direct interspecies electron transfer mediated through surface oxygen-containing functional groups. Environ. Sci. Technol. In press.
- Zhang, J.; Lu, Y. Conductive Fe3O4 nanoparticles accelerate syntrophic methane production from butyrate oxidation in two different lake sediments. Front. Microbiol. 2016, 7, 1316–1325. [Google Scholar] [CrossRef]
- Cruz Viggi, C.; Rossetti, S.; Fazi, S.; Paiano, P.; Majone, M. Magnetite particles triggering a faster and more robust syntrophic pathway of methanogenic propionate degradation. Environ. Sci. Technol. 2014, 48, 7536–7543. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, Y.; Li, Y.; Quan, X.; Zhao, Z. Comparing the mechanisms of ZVI and Fe3O4 for promoting waste activated sludge digestion. Water Res. 2018, 1444, 126–133. [Google Scholar] [CrossRef]
- Pong, H.; Zhang, Y.; Tan, D.; Zhao, Z.; Zhao, H.; Quan, X. Roles of magnetite and granular activated carbon in improvement of anaerobic sludge digestion. Bioresour. Technol. 2018, 249, 666–672. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhaohui, Y.; Xu, R.; Xiang, Y.; Jia, M.; Hu, J.; Zheng, Y.; Xiong, W.; Cao, J. Enhanced mesophilic anaerobic digestion of waste sludge with the iron nanoparticles addition and kinetic analysis. Sci. Total Environ. 2019, 686, 124–133. [Google Scholar] [CrossRef]
- Su, L.; Shi, X.; Gun, G.; Zhano, A.; Zhao, Y. Stabilization of sewage sludge in the presence of nanoscale zero valent iron (nZVI): Abatement of odor and improvement of biogas production. J. Mater. Cycles Waste 2013, 15, 461–468. [Google Scholar] [CrossRef]
- Abdelsalam, E.; Samer, M.; Attia, Y.; Abdel Hadi, M.; Hassan, H.; Badr, Y. Influence of zero valent iron nanoparticles and magnetic iron oxide nanoparticles on biogas and methane production from anaerobic digestion of manure. Energy 2017, 120, 842–853. [Google Scholar] [CrossRef]
- Lizama, A.; Figueiras, C.; Pedreguera, A.; Espinoza, J. Enhancing the performance and stability of the anaerobic digestion of sewage sludge by zero valent iron nanoparticles dosage. Bioresour. Technol. 2019, 275, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Zhang, Y.; Quan, X.; Chen, S. Enhanced anaerobic digestion of waste activated sludge digestion by the addition of zero valent iron. Water Res. 2014, 52, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Wei, J.; van Loosdrecht, M.; Cao, D. Analyzing the mechanisms of sludge digestion enhanced by iron. Water Res. 2017, 117, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Baek, G.; Kim, J.; Lee, C. A review of the effects of iron compounds on methanogenesis in anaerobic environments. Renew. Sust. Energy Rev. 2019, 113, 109282. [Google Scholar] [CrossRef]
- Wei, W.; Cai, Z.; Fu, J.; Xie, G.; Li, A.; Zhou, X.; Ni, B. Zero valent iron enhances methane production from primary sludge in anaerobic digestion. Chem. Eng. J. 2018, 351, 1159–1165. [Google Scholar] [CrossRef]
- Yu, B.; Huang, X.; Zhang, D.; Lou, Z.; Yuan, H.; Zhu, N. Response of sludge fermentation liquid and microbial community to nano zero valent iron exposure in a mesophilic anaerobic digestion system. RSC Adv. 2016, 6, 24236–24243. [Google Scholar] [CrossRef]
- Lee, C.; Kim, J.; Lee, W.; Nelson, K.; Yoon, J.; Sedlak, D. Bactericidal effect of zero valent iron nanoparticles on Escherichia coli. Environ. Sci. Technol. 2008, 42, 4927–4933. [Google Scholar] [CrossRef]
- Kim, J.; Park, H.; Lee, C.; Kara, N.; Sedlak, D.; Yoon, J. Inactivation of Escherichia coli by nano particulate zero valent iron and ferrous ion. Appl. Environ. Microbiol. 2010, 76, 7668–7670. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, J.; Hu, Z. Impact of nano zero valent iron (NZVI) on methanogenic activity and population dynamics in anaerobic digestion. Water Res. 2013, 47, 6790–6800. [Google Scholar] [CrossRef]
- Cho, Y.; Young, J.; Jordan, J.; Moon, H. Factors affecting measurement of specific methanogenic activity. Water Sci. Technol. 2005, 52, 435–440. [Google Scholar] [CrossRef]
- Kang, Y.; Risbud, S.; Rabolt, J.; Stroeve, P. Synthesis and characterization of nanometer size Fe3O4 and Fe2O3 particles. Chem. Mater. 1996, 8, 2209–2211. [Google Scholar] [CrossRef]
- He, F.; Zhao, D.; Liu, J.; Roberts, C. Stabilization of Fe-Pd nanoparticles with sodium carboxymethyl cellulose for enhanced transport and dechlorination of trichloroethylene in soil and groundwater. Ind. Eng. Chem. Res. 2007, 46, 29–34. [Google Scholar] [CrossRef]
- Angelidaki, I.; Alves, M.; Bolzonella, D.; Borzacconi, L.; Campos, J.; Guwy, A.; Kalyuzhnyi, S.; Jenicke, P.; van Lier, J. Defining the biomethane potential (BMP) of solid organic wastes and energy crops: A proposed protocol for batch assays. Water Sci. Technol. 2009, 59, 927–934. [Google Scholar] [CrossRef] [PubMed]
- Pabon, P.; Castanares, G.; van Lier, J. An oxiTop protocol for screening plant material for its biochemical methane potential (BMP). Water Sci. Technol. 2012, 66, 1416–1423. [Google Scholar] [CrossRef] [PubMed]
- Baird, R.; Bridgewater, L. Standard Methods for the Examination of Water and Wastewater, 23rd ed.; American Public Health Association: Washington, DC, USA, 2017. [Google Scholar]
- Radojevic, M.; Bashkin, V. Practical Environmental Analysis; RSC Publishing: Cambridge, UK, 2006. [Google Scholar]
- Buswell, A.; Mueller, H. Mechanism of methane fermentation. Ind. Eng. Chem. 1952, 44, 550–552. [Google Scholar] [CrossRef]
- Elbeshbishy, E.; Nakhla, G. Batch anaerobic co-digestion of proteins and carbohydrates. Bioresour. Technol. 2012, 116, 170–178. [Google Scholar] [CrossRef]
- Fisgativa, H.; Tremier, A.; Dabert, P. Characterizing the variability of food waste quality: A need for efficient valorization through anaerobic digestion. Waste Manag. 2016, 50, 264–274. [Google Scholar] [CrossRef]
- Zhang, C.; Su, H.; Baeyens, J.; Tan, T. Reviewing the anaerobic digestion of food waste for biogas production. Renew. Sust. Energy Rev. 2014, 38, 383–392. [Google Scholar] [CrossRef]
- Liu, C.; Li, H.; Zhang, Y.; Liu, C. Improve biogas production from low-organic- content sludge through high-solids anaerobic co-digestion with food waste. Bioresour. Technol. 2016, 219, 252–260. [Google Scholar] [CrossRef]
- Fisgativa, H.; Tremier, A.; Roux, S.; Bureau, C.; Dabert, P. Understanding the anaerobic biodegradability of food waste: Relationship between the typological, biochemical and microbial characteristics. J. Environ. Manag. 2017, 188, 95–107. [Google Scholar] [CrossRef]
- Liu, L.; He, Q.; Ma, Y.; Wang, X.; Peng, X. A mesophilic anaerobic digester for treating food waste: Process stability and microbial community analysis using pyro sequencing. Microb. Cell Factories 2016, 15, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Duan, N.; Dong, B.; Dai, L. High solids anaerobic co-digestion of sewage and food waste in comparison with mono digestion: Stability and performance. Waste Manag. 2013, 33, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Gou, C.; Yang, Z.; Huang, J.; Wang, H.; Xu, H.; Wang, L. Effects of temperature and organic loading rate on the performance and microbial community of anaerobic co-digestion of waste activated sludge and food waste. Chemosphere 2014, 105, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Koch, K.; Plabst, M.; Schmidt, A.; Helmreich, B.; Drewes, J. Co-digestion of food waste in a municipal wastewater treatment plant: Comparison of batch tests and full scale experiences. Waste Manag. 2016, 47, 28–33. [Google Scholar] [CrossRef]
- Zhang, R.; El-Mashad, H.; Hartman, K.; Wang, F.; Liu, G.; Choate, C.; Gamble, P. Characterization of food waste as feedstock for anaerobic digestion. Bioresour. Technol. 2007, 98, 929–935. [Google Scholar] [CrossRef]
- El-Mashad, H.; Zhang, R. Biogas production from co-digestion of dairy manure and food waste. Bioresour. Technol. 2010, 101, 4021–4028. [Google Scholar] [CrossRef]
- Zhang, L.; Lee, Y.; Jahng, D. Anaerobic co-digestion of food waste and piggery wastewater: Focusing on the role of trace elements. Bioresour. Technol. 2011, 102, 5048–5059. [Google Scholar] [CrossRef]
- Agyeman, F.; Tao, W. Anaerobic co-digestion of food waste and dairy manure: Effects of food waste particle size and organic loading rate. J. Environ. Manag. 2014, 133, 268–274. [Google Scholar] [CrossRef]
- Zhang, Y.; Banks, C.; Heaven, S. Co-digestion of source segregated domestic food waste to improve process stability. Bioresour. Technol. 2012, 114, 168–178. [Google Scholar] [CrossRef]
- Von Sperling, M.; Chernicharo, C. Biological Wastewater Treatment in Warm Climate Regions; IWA Publishing: London, UK, 2005. [Google Scholar]
- El-Mashad, H.; Zhang, R. Co-digestion of food waste and dairy maure for biogas production. Tasabe 2007, 50, 1815–1821. [Google Scholar]
- Zhao, Z.; Li, Y.; Yu, Q.; Zhang, Y. Ferroferric oxide triggered possible direct interspecies electron transfer between syntrophomonas and Methanosaeta to enhance waste activated sludge anaerobic digestion. Bioresour. Technol. 2018, 250, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Guo, L.; Wang, Y.; Zhao, Y.; She, Z.; Gao, M.; Guo, Y. Application of iron oxide (Fe3O4) nanoparticles during the two stage anaerobic digestion with waste sludge: Impact on the biogas production and the substrate metabolism. Renew. Energy 2020, 146, 2724–2735. [Google Scholar] [CrossRef]
- Yin, Q.; He, K.; Echigo, S.; Wu, G.; Zhan, X.; Hu, H. Ferroferric oxide significantly affected production soluble microbial products and extracellular polymeric substances in anaerobic methangenesis reactors. Front. Microbiol. 2018, 9, 2376. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Sun, F.; Liu, J.; Zhao, X. Enhanced anaerobic phenol degradation by conductive materials via EPS and microbial community alteration. Chem. Eng. J. 2018, 352, 1–9. [Google Scholar] [CrossRef]
- Oh, S.; Kang, S.; Azizi, A. Electrochemical communication in anaerobic digestion. Chem. Eng. J. 2018, 353, 878–889. [Google Scholar] [CrossRef]
- Hoffman, M.; Decho, A. Extracellular enzymes within microbial biofilms and the role of the extracellular plymer matrix. In Microbial Extracellular Polymeric Substances: Characterization, Structure and Function, 1st ed.; Wingender, J., Neu, T., Flemming, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 217–227. [Google Scholar]
- Baek, G.; Kim, J.; Lee, C. A long study on the effect of magnetite supplementation in continuous anaerobic digestion of dairy effluent-Enhancement in process performance and stability. Bioresour. Technol. 2016, 222, 344–354. [Google Scholar] [CrossRef]
- Baek, G.; Jung, H.; Kim, J.; Lee, C. A long-term study on the effect of magnetite supplementation in continuous anaerobic digestion of dairy effluent. Bioresour. Technol. 2017, 241, 830–840. [Google Scholar] [CrossRef]
- Li, H.; Chang, J.; Liu, P.; Fu, L.; Ding, D.; Lu, Y. Direct interspecies electron transfer accelerates syntrophic oxidation of butyrate in paddy soil enrichments. Environ. Microbiol. 2014, 17, 1533–1547. [Google Scholar] [CrossRef]
- He, C.; He, P.; Yang, H.; Li, L.; Lin, Y.; Mu, Y. Impact of zero-valent iron nanoparticles on the activity of anaerobic granular sludge: From macroscopic to microcosmic investigation. Water Res. 2017, 127, 32–40. [Google Scholar] [CrossRef]
- Jia, T.; Wang, Z.; Shan, H.; Liu, Y.; Lei, G. Effect of nanoscale zero-valent iron on sludge anaerobic digestion. Resour. Conserv. Recycl. 2017, 127, 190–195. [Google Scholar] [CrossRef]
- Suanon, F.; Sun, Q.; Mama, D.; Li, J.; Dimon, B. Effect of nanoscale zero- valent iron and magnetite (Fe3O4) on the fate of metals during anaerobic digestion of sludge. Water Res. 2016, 88, 897–903. [Google Scholar] [CrossRef]
- Gonzalez-Estrella, J.; Sierra-Alvarez, R.; Field, J. Toxicity assessment of inorganic nanoparticles to acetoclastic and hydrogentrophic methanogenic activity in anaerobic granular sludge. J. Hazard. Mater. 2013, 260, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Rosicka, D.; Sembera, J. Assessment of influence of magnetic forces on aggregation of zero valent iron nanoparticles. Nanoscale Res. Lett. 2011, 6, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Lo, I. Magnetic nanoparticles: Essential factors for sustainable environmental application. Water Res. 2013, 47, 2613–2632. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Zeng, G.; Huang, D.; Feng, C.; Hu, S.; Zhao, M.; Lai, C.; Wei, Z.; Huang, C.; Xie, G.; et al. Use of iron oxide nanomaterials in wastewater treatment: A review. Sci. Total Environ. 2012, 424, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Wu, Z.; Yu, T.; Jiang, C.; Kim, W. Recent progress on magnetic iron oxide nanoparticles: Synthesis, surface functional strategies and biomedical application. Sci. Technol. Adv. Mater. 2015, 16, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Hutchins, D.; Downey, J. Effective separation of magnetite nanoparticles within an industrial scale pipeline reactor. Sep. Sci. Technol. 2019, 1–8. [Google Scholar] [CrossRef]
- Rui, M.; Ma, C.; Hao, Y.; Guo, J.; Rui, Y.; Tang, X.; Qi, Z.; Fan, X.; Zetian, Z.; Hou, T.; et al. Iron oxide nanoparticles as a potential iron fertilizer for peanut (Arachis hypogaea). Front. Plant Sci. 2016, 7, 815. [Google Scholar] [CrossRef]
- Li, J.; Chang, P.; Huang, J.; Wang, Y.; Yuan, H.; Ren, H. Physiological effects of magnetic iron oxide nanoparticles towards watermelon. J. Nanosci. Nanotechnol. 2013, 13, 5561–5567. [Google Scholar] [CrossRef]
- Boutchuen, A.; Zimmerman, D.; Aich, N.; Masud, A.; Arabshahi, A.; Palchoundhury, S. Increased plant growth with haematite nanoparticles fertilizer drop and determining nanoparticles uptake in plants using multimodal approach. J. Nanomater. 2019, 2019, 6890572. [Google Scholar] [CrossRef]
- Elfeky, S.; Mohammed, M.; Khater, M.; Osman, Y.; Elsherbini, E. Effect of magnetite nano-fertilizer on growth and yield of Ocimum basilicum L. Int. J. Indig. Med. Plants 2013, 46, 1286–1293. [Google Scholar]
- Plaksenkova, I.; Jermalonoka, M.; Bankovska, L.; Gavarane, I.; Gerbreders, V.; Sledevskis, E.; Snikeris, J.; Kokina, I. Effects of Fe3O4 nanoparticles stress on the growth and development of rocket Eruca sativa. J. Nanomater. 2019, 2019, 2678247. [Google Scholar] [CrossRef]
- Abou El-Nasr, M.; El-Hennawy, H.; El-Kereamy, A.; Abou-El-Yazied, A.; Salah Eldin, T. Effect of magnetite nanoparticles (Fe3O4) as nutritive supplement on pear saplings. Middle East J. Appl. Sci. 2015, 5, 777–785. [Google Scholar]
- Govea-Alcaide, E.; Masunaga, S.; De Souza, A.; Fajardo-Rosabal, L.; Effenberger, F.; Rossi, L.; Jardim, R. Tracking iron oxide nanoparticles in plant organs using magnetic measurements. J. Nanoparticle Res. 2016, 18, 305. [Google Scholar] [CrossRef]
- Kanjana, D. Foliar study on effect of iron oxide nanoparticles as an alternate source of iron fertilizer to cotton. Int. J. Chem. Stud. 2019, 7, 4374–4379. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
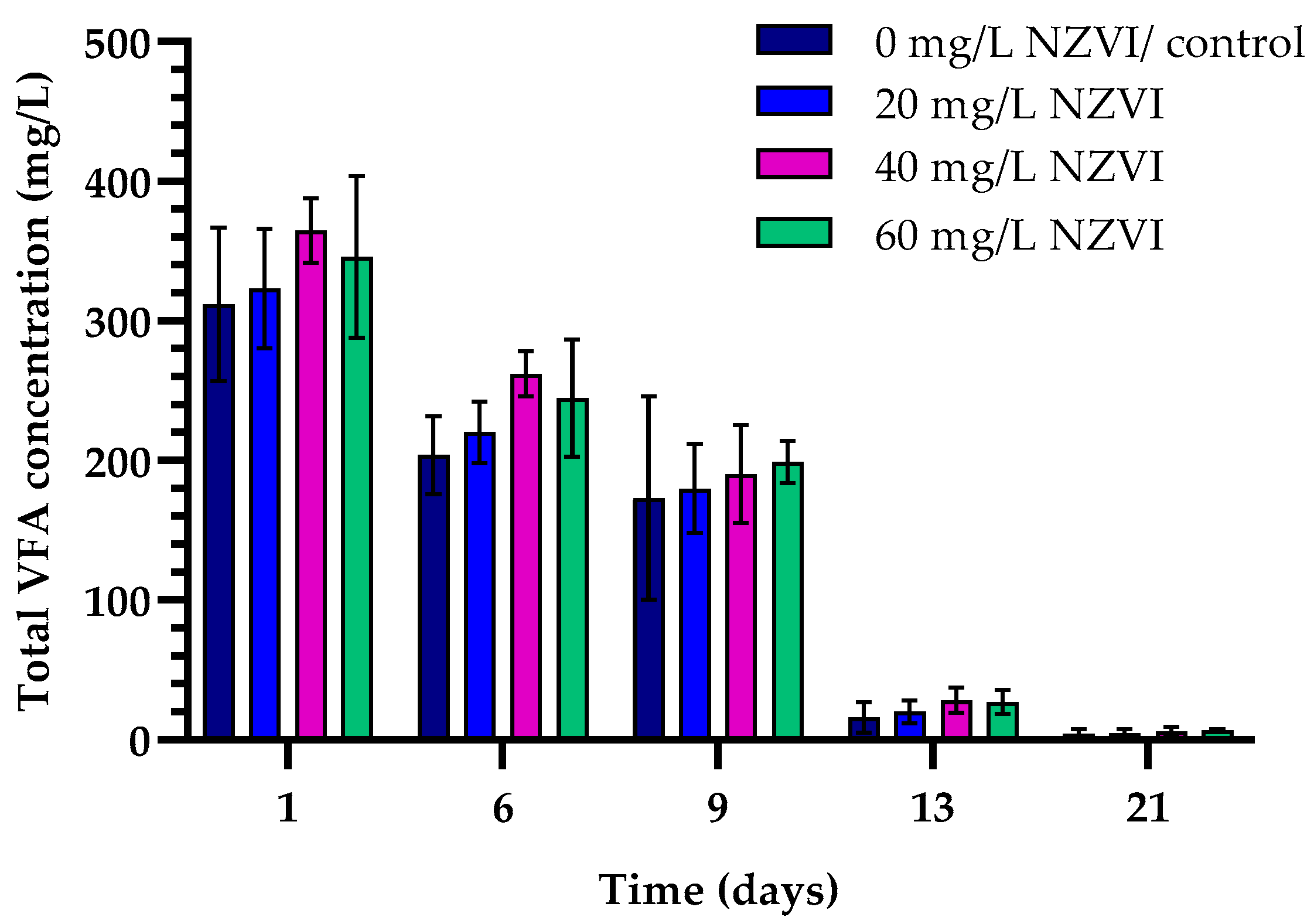{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | n | FW | WAS | ||
|---|---|---|---|---|---|
| (σ) | CV (%) | (σ) | CV (%) | ||
| Physicochemical characteristics | |||||
| pH | 8 | 4.1 (0.5) | 12 | 7.4 (0.7) | 10 |
| TS (gTS/kg wet weight) | 10 | 298.8 (21.6) | 7 | ||
| (gTS/L) | 25.2 (3.4) | 13 | |||
| VS (gVS/kg wet weight) | 10 | 282.1 (24.6) | 9 | ||
| (gVS/kgTS) | 956.0 (10.9) | 1 | |||
| (gVS/L) | 21.1 (3.1) | 15 | |||
| Soluble COD (gO2/kgVS) (mg/L) | 8 | 311.2 (61.1) | 20 | 654 (51) | 8 |
| Total Kjeldahl nitrogen (gN/kgVS) | 8 | 35.1 (2.4) | 7 | 107.7 (5.9) | 5 |
| Total ammonia nitrogen (gN/kg VS ) | 8 | 2.30 (0.5) | 23 | 20 (3.0) | 15 |
| Total phosphorous (gP/kg VS) | 8 | 2.6 (0.3) | 13 | 20.9 (3.40) | 16 |
| Volatile fatty acids (g COD/kg VS) | 8 | 3.7 (0.50) | 12 | 13.10 (3.16) | 24 |
| C/N (%) | 17.6 | 5.5 | |||
| Elementary analysis | |||||
| Carbon (%DM) | 8 | 52.3 (4.7) | 9 | 42.71 (3.14) | 7 |
| Hydrogen (%DM) | 8 | 7.2 (0.8) | 11 | 6.89 (0.6) | 9 |
| Oxygen (%DM) | 8 | 37.1 (6.1) | 16 | 42.5 (4.23) | 10 |
| Nitrogen (%DM) | 8 | 3.4 (0.7) | 20 | 7.9 (1.1) | 14 |
| Cl− (mg/kg DM) | 8 | 11029 (1376.8) | 12 | 6813 (991.6) | 15 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kassab, G.; Khater, D.; Odeh, F.; Shatanawi, K.; Halalsheh, M.; Arafah, M.; van Lier, J.B. Impact of Nanoscale Magnetite and Zero Valent Iron on the Batch-Wise Anaerobic Co-Digestion of Food Waste and Waste-Activated Sludge. Water 2020, 12, 1283. https://doi.org/10.3390/w12051283
Kassab G, Khater D, Odeh F, Shatanawi K, Halalsheh M, Arafah M, van Lier JB. Impact of Nanoscale Magnetite and Zero Valent Iron on the Batch-Wise Anaerobic Co-Digestion of Food Waste and Waste-Activated Sludge. Water. 2020; 12(5):1283. https://doi.org/10.3390/w12051283
Chicago/Turabian StyleKassab, Ghada, Dima Khater, Fadwa Odeh, Khaldoun Shatanawi, Maha Halalsheh, Mazen Arafah, and Jules B. van Lier. 2020. "Impact of Nanoscale Magnetite and Zero Valent Iron on the Batch-Wise Anaerobic Co-Digestion of Food Waste and Waste-Activated Sludge" Water 12, no. 5: 1283. https://doi.org/10.3390/w12051283
APA StyleKassab, G., Khater, D., Odeh, F., Shatanawi, K., Halalsheh, M., Arafah, M., & van Lier, J. B. (2020). Impact of Nanoscale Magnetite and Zero Valent Iron on the Batch-Wise Anaerobic Co-Digestion of Food Waste and Waste-Activated Sludge. Water, 12(5), 1283. https://doi.org/10.3390/w12051283