Escherichia coli Antimicrobial Resistance Variability in Water Runoff and Soil from a Remnant Native Prairie, an Improved Pasture, and a Cultivated Agricultural Watershed

 ,
,  ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
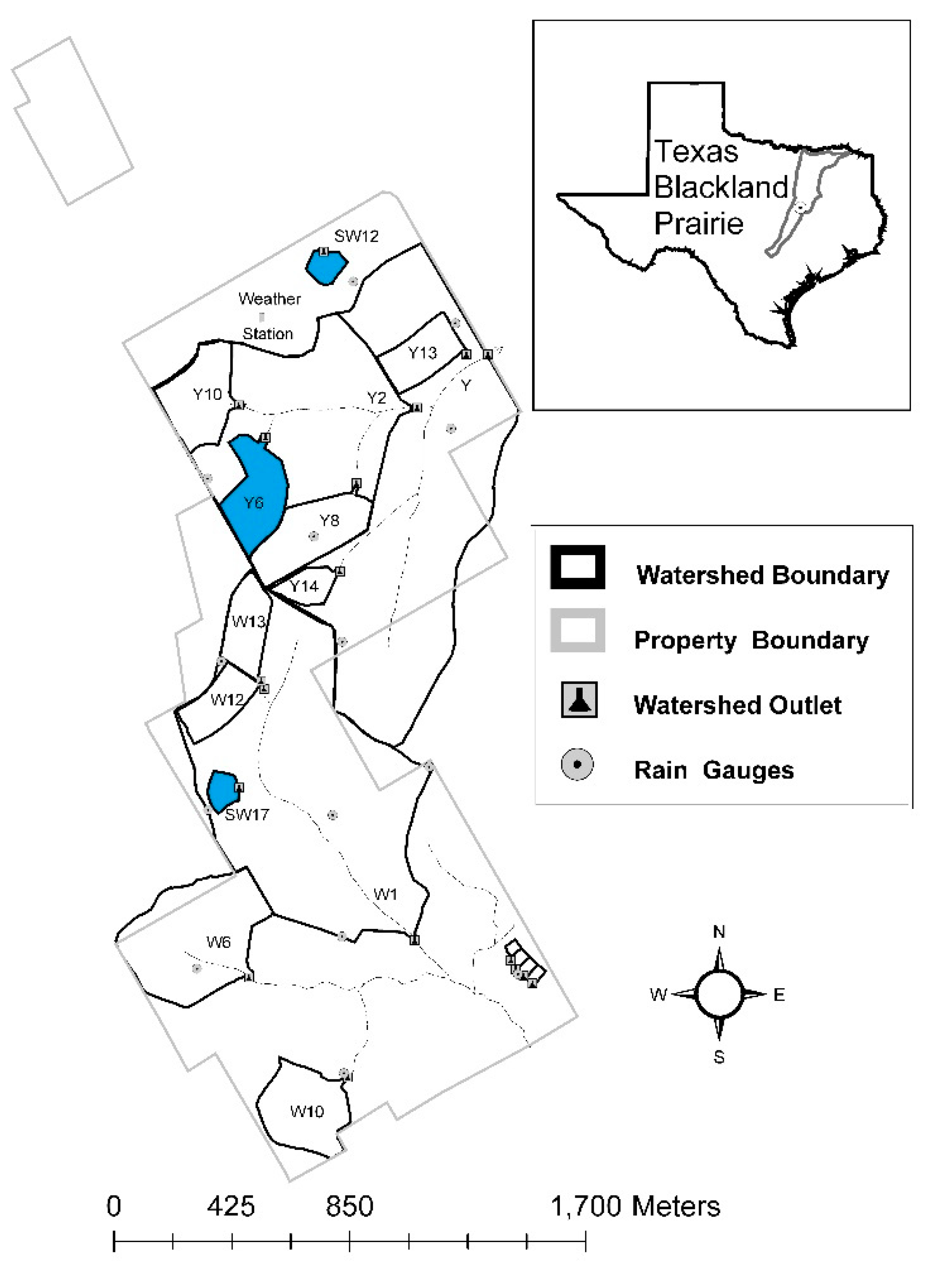
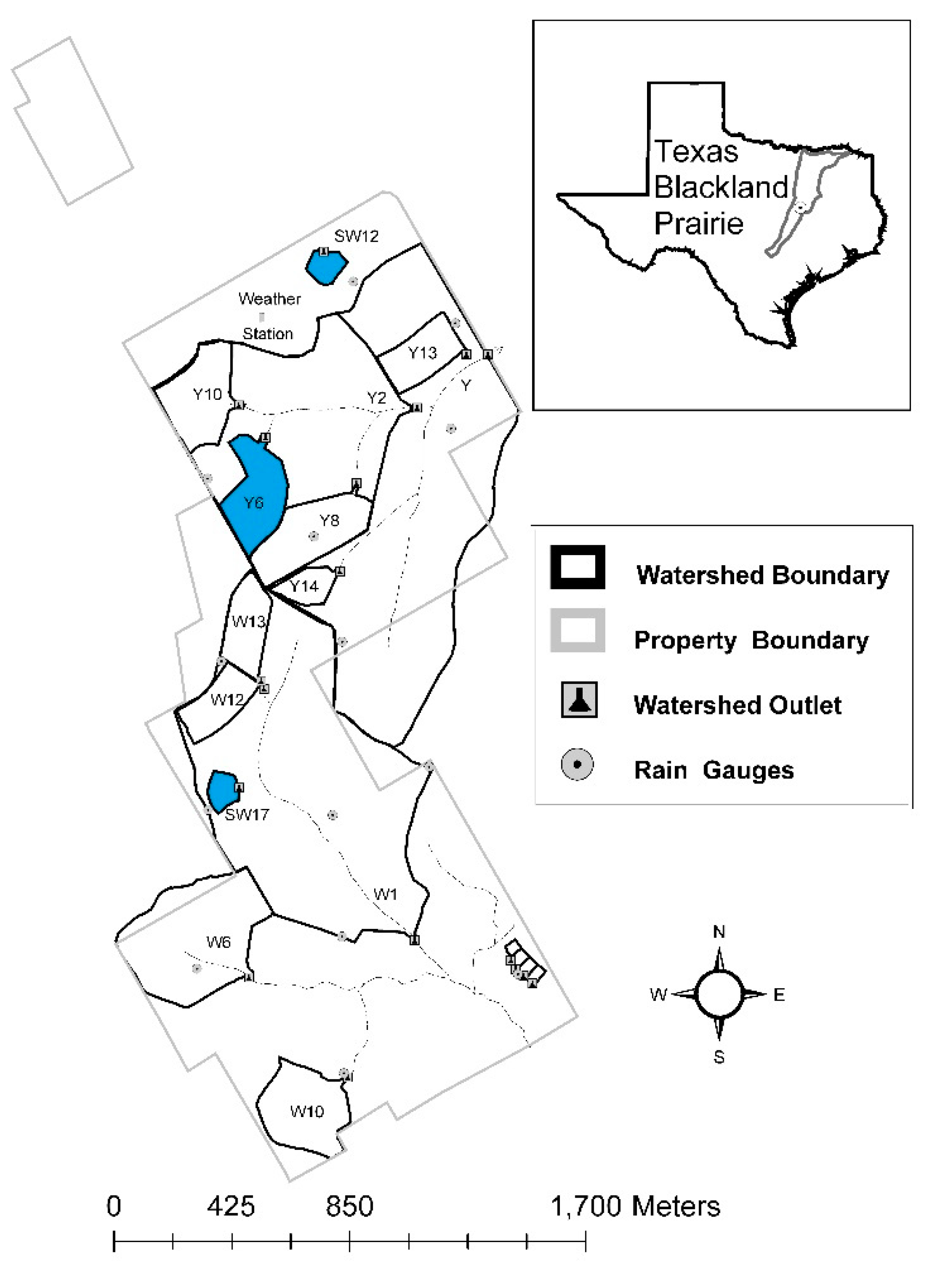
2.1. Site Description
2.2. Sampling Duration and Methods
2.3. E. coli Isolation and Archiving from Environmental Samples
2.4. Antimicrobial Resistance Analysis
2.5. ERIC-PCR/Riboprinting Composite Fingerprinting and BioNumerics Analysis
2.6. Statistical Analysis
3. Results
3.1. E. coli Population Within the Three Sites—Water Runoff vs. Soil
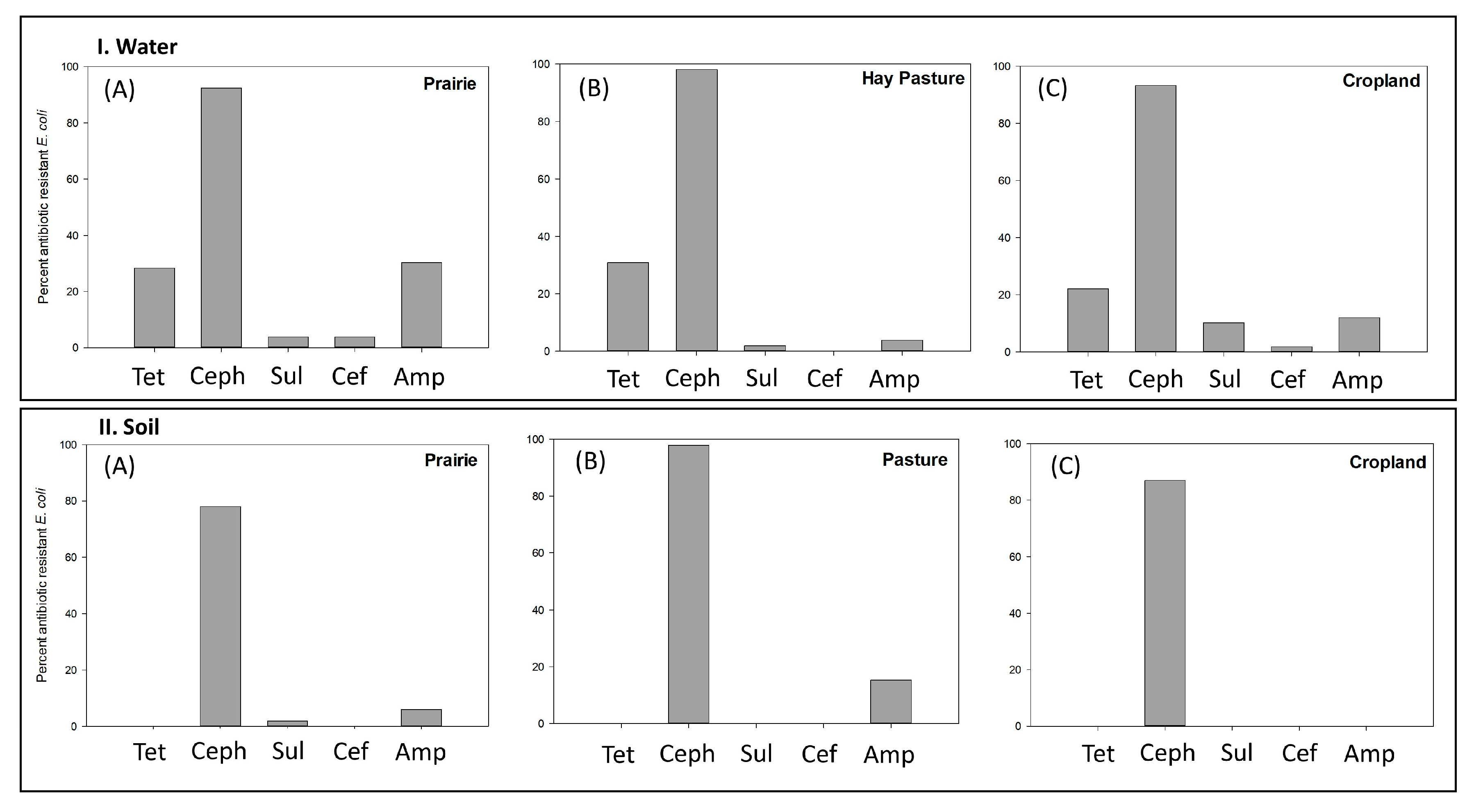
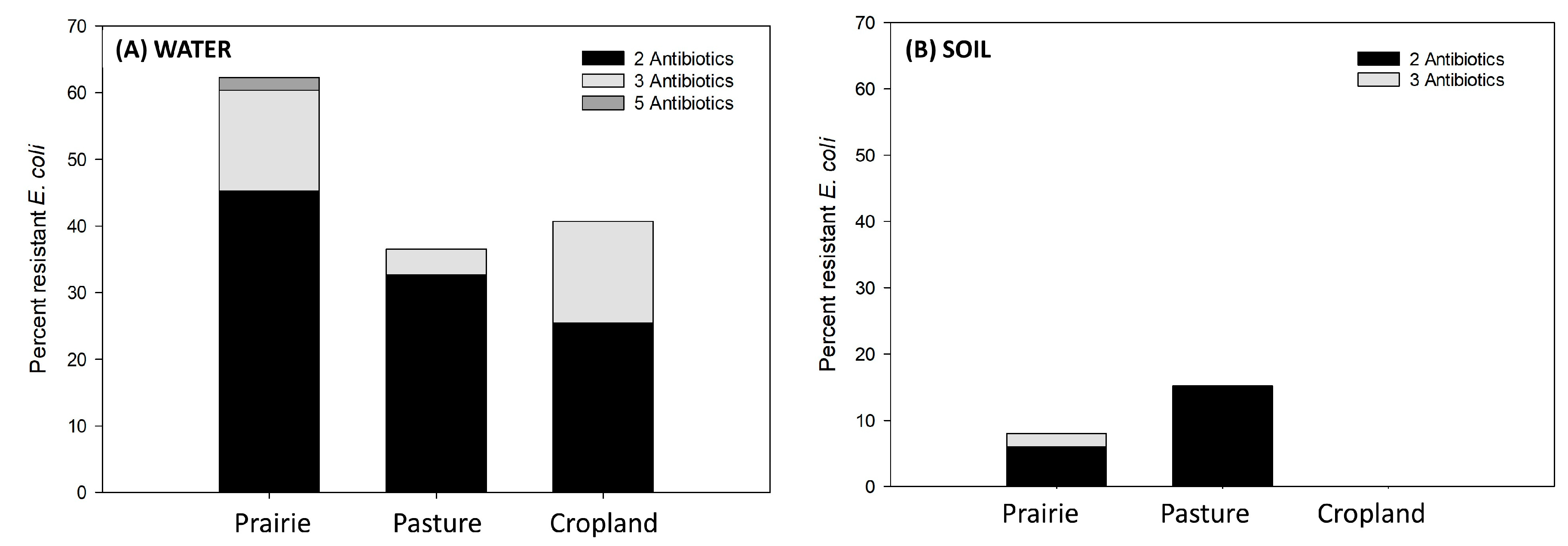
3.2. Percentage of Antimicrobial Resistance within the Three Study Sites—Water Runoff vs. Soil
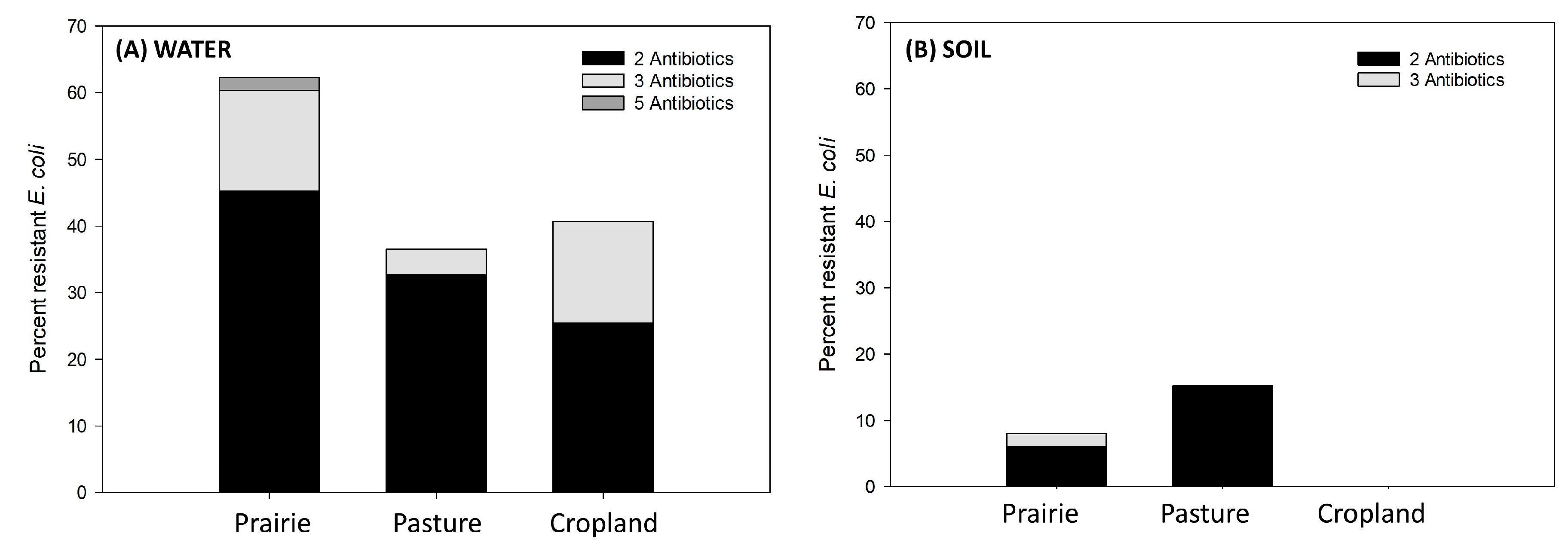
3.3. Percentage of MDR within the Three Study Sites—Water Runoff vs. Soil
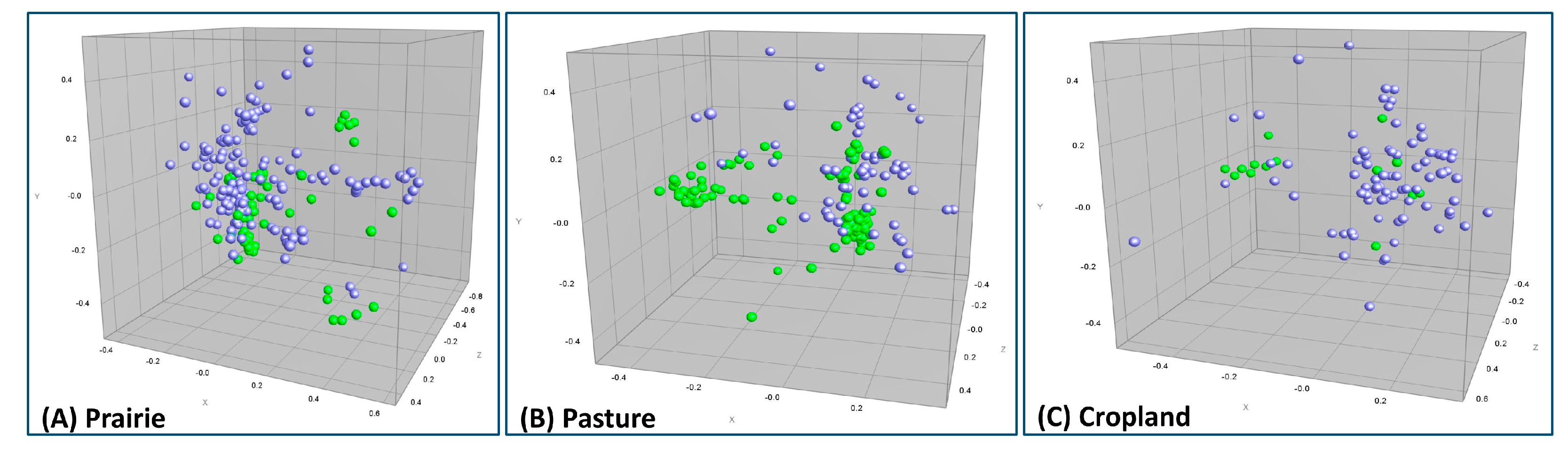
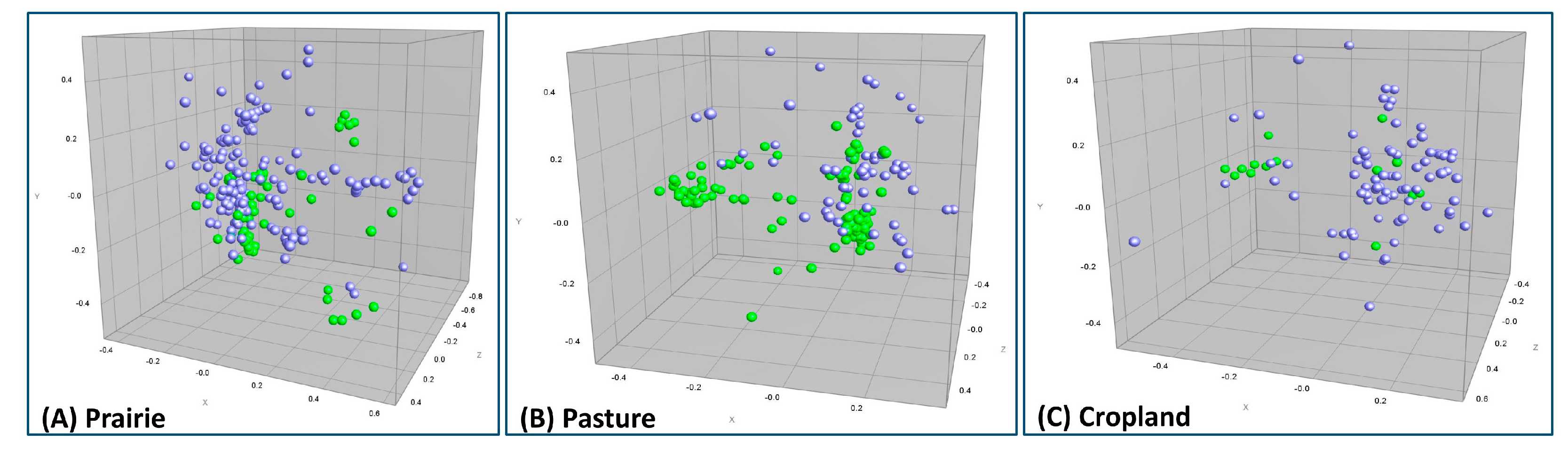
3.4. Composite ERIC-RP Fingerprint Comparison between Runoff and Soil E. coli Isolates
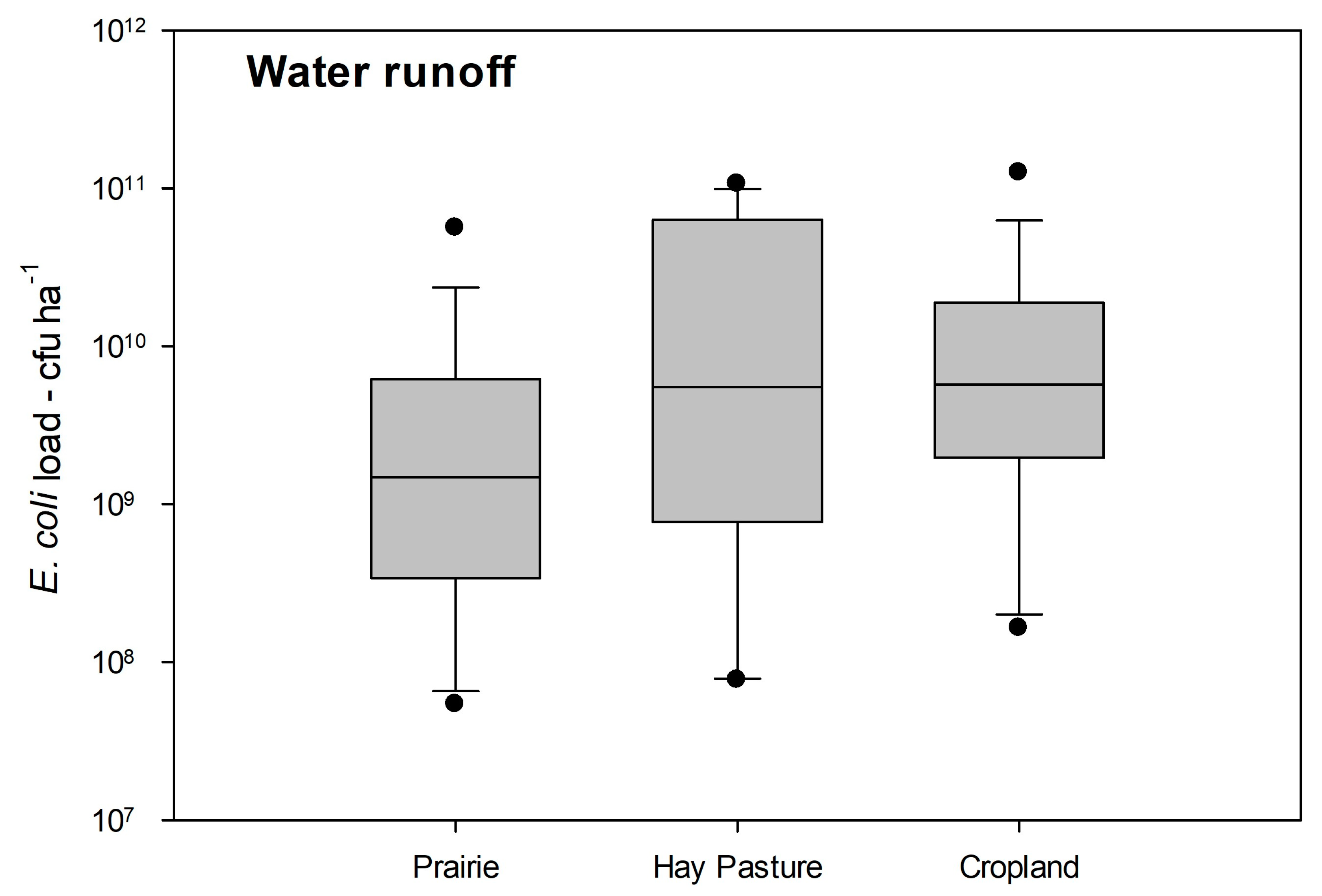
3.5. AMR and MDR within Study Sites per Hectare
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kumar, K.; Gupta, S.C.; Chander, Y.; Singh, A.K. Antibiotic use in agriculture and its impact on the terrestrial environment. Advan. Agronom. 2005, 87, 1–54. [Google Scholar]
- Center for Disease Control—Antibiotic Resistance Threats in the United States. 2019. Available online: https://www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-report-508.pdf (accessed on 27 April 2020).
- Friedman, N.D.; Temkin, E.; Carmeli, Y. The negative impact of antibiotic resistance. Clin. Microbiol. Infect. 2016, 22, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Eliopoulos, G.M.; Cosgrove, S.E.; Carmeli, Y. The impact of antimicrobial resistance on health and economic outcomes. Clin. Infec. Dis. 2003, 36, 1433–1437. [Google Scholar] [CrossRef] [PubMed]
- Allen, H.K.; Donato, J.; Wang, H.H.; Cloud-Hansen, K.A.; Davies, J.; Handelsman, J. Call of the wild: Antimicrobial resistance genes in natural environments. Nat. Rev. Microbiol. 2010, 8, 251–259. [Google Scholar] [CrossRef]
- Wellington, E.M.H.; Boxall, A.B.A.; Cross, P.; Feil, E.J.; Gaze, W.H.; Hawkey, P.M.; Johnson-Rollings, A.S.; Jones, D.L.; Lee, N.M.; Otten, W.; et al. The role of the natural environment in the emergence of antimicrobial resistance in Gram-negative bacteria. Lancet. Infect. Dis. 2013, 13, 155–165. [Google Scholar] [CrossRef]
- Martinez, J.L. Antibiotics and antimicrobial resistance genes in natural environments. Science 2008, 321, 365–367. [Google Scholar] [CrossRef]
- Wright, G.D. Antimicrobial resistance in the environment: A link to the clinic? Curr. Opin. Microbiol. 2010, 13, 589–594. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, T.; Fang, H.H.P. Antimicrobial resistance genes in water environment. Appl. Microbiol. Biotechnol. 2009, 82, 397–414. [Google Scholar] [CrossRef]
- Singer, A.C.; Shaw, H.; Rhodes, V.; Hart, A. Review of antimicrobial resistance in the environment and its relevance to environmental regulators. Front. Microbiol. 2016, 7, 1728. [Google Scholar] [CrossRef] [Green Version]
- D’costa, V.M.; McGrann, K.M.; Hughes, D.W.; Wright, G.D. Sampling the antibiotic resistome. Science 2006, 311, 374–377. [Google Scholar] [CrossRef] [Green Version]
- Allen, H.K.; Moe, L.A.; Rodbumrer, J.; Gaarder, A.; Handelsman, J. Functional metagenomics reveals diverse beta-lactamases in a remote Alaskan soil. ISME J. 2009, 3, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Riesenfeld, C.S.; Goodman, R.M.; Handelsman, J. Uncultured soil bacteria are a reservoir of new antimicrobial resistance genes. Environ. Microbiol. 2004, 6, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Bacterial diversity and antimicrobial resistance in water habitats: Searching the links with the human microbiome. FEMS Microbiol. Rev. 2014. 38, 761–818.
- Sayah, R.S.; Kaneene, J.B.; Johnson, Y.; Miller, R.S. Patterns of antimicrobial resistance observed in Escherichia coli isolates obtained from domestic- and wild-animal fecal samples, human septage, and surface water. Appl. Environ. Microbiol. 2005, 71, 1394–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baquero, F.; Martınez, J.M.; Canto, R. Antibiotics and antimicrobial resistance in water environments. Curr. Opin. Biotech. 2008, 19, 260–265. [Google Scholar] [CrossRef]
- Cho, S.; Hiott, L.M.; McDonald, J.M.; Barrett, J.B.; McMillan, E.A.; House, S.L.; Adams, E.S.; Frye, J.G.; Jackson, C.R. Diversity and antimicrobial resistance of Enterococcus from upper Oconee Watershed, Georgia. J. Appl. Microbiol. 2019, 128, 1221–1233. [Google Scholar] [CrossRef]
- Sullivan, B.A.; Karthikeyan, R. Occurrence and prevalence of tetracycline resistant bacteria in a rapidly urbanizing subtropical watershed. J. Nat. Environ. Sci. 2011, 2, 25–31. [Google Scholar]
- Sullivan, B.A.; Gentry, T.J.; Karthikeyan, R. Characterization of tetracycline-resistant bacteria in an urbanizing subtropical watershed. J. Appl. Microbiol. 2013, 11, 774–785. [Google Scholar] [CrossRef]
- Brooks, J.P.; Adeli, A.; McLaughlin, M.R. Microbial ecology, bacterial pathogens, and antibiotic resistant genes in swine manure wastewater as influenced by three swine management systems. Water Res. 2014, 57, 96–103. [Google Scholar] [CrossRef]
- Zhang, X.; Li, Y.; Liu, B.; Wang, J.; Feng, C.; Gao, M.; Wang, L. Prevalence of veterinary antibiotics and antibiotic-resistant Escherichia coli in the surface water of a livestock production region in northern China. PLoS ONE 2014, 9, e111026. [Google Scholar] [CrossRef]
- Watkinson, A.J.; Murby, E.J.; Kolpin, D.W.; Costanzo, S.D. The occurrence of antibiotics in an urban watershed: From wastewater to drinking water. Sci. Tot. Environ. 2008, 407, 2711–2723. [Google Scholar] [CrossRef] [PubMed]
- Servais, P.; Passerat, J. Antimicrobial resistance of fecal bacteria in waters of the Seine river watershed (France). Sci. Tot. Environ. 2009, 408, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Edge, T.A.; Hill, S. Occurrence of antimicrobial resistance in Escherichia coli from surface waters and fecal pollution sources near Hamilton, Ontario. Can. J. Microbiol. 2005, 51, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Hatosy, S.M.; Martiny, A.C. The ocean as a global reservoir of antimicrobial resistance genes. Appl. Environ. Microbiol. 2015, 81, 7593–7599. [Google Scholar] [CrossRef] [Green Version]
- Segawa, T.; Takeuchi, N.; Rivera, A.; Yamada, A.; Yoshimura, Y.; Barcaza, G.; Shinbori, K.; Motoyama, H.; Kohshima, S.; Ushida, K. Distribution of antimicrobial resistance genes in glacier environments. Environ. Microbiol. Rep. 2013, 5, 127–134. [Google Scholar] [CrossRef]
- Miller, R.V.; Gammon, K.; Day, M.J. Antimicrobial resistance among bacteria isolated from seawater and penguin fecal samples collected near Palmer Station, Antarctica. Can. J. Microbiol. 2009, 55, 37–45. [Google Scholar] [CrossRef]
- Martiny, A.C.; Martiny, J.B.H.; Weihe, C.; Field, A.; Ellis, J.C. Functional metagenomics reveals previously unrecognized diversity of antimicrobial resistance genes in gulls. Front. Microbiol. 2011, 2, 238. [Google Scholar] [CrossRef] [Green Version]
- Harwood, V.J.; Whitlock, J.; Withington, V. Classification of antimicrobial resistance patterns of indicator bacteria by discriminant analysis: Use in predicting the source of fecal contamination in subtropical waters. Appl. Environ. Micro. 2000, 66, 3698–3704. [Google Scholar] [CrossRef] [Green Version]
- Price, R.G.; Wideboer, D.E. coli as an indicator of contamination and health risk in environmental waters. In Escherichia coli—Recent Advances on Physiology, Pathogenesis and Biotechnological Applications; IntechOpen: London, UK, 2016. [Google Scholar] [CrossRef] [Green Version]
- Bernhard, A.E.; Field, K.G. Identification of nonpoint sources of fecal pollution in coastal waters by using host-specific 16S ribosomal DNA genetic markers from fecal anaerobes. Appl. Environ. Microbiol. 2000, 66, 1587–1594. [Google Scholar] [CrossRef] [Green Version]
- Guber, A.K.; Fry, J.; Ives, R.L.; Rose, J.B. Escherichia coli survival in, and release from, white-tailed deer feces. Appl. Environ. Microbiol. 2015, 81, 1168–1176. [Google Scholar] [CrossRef]
- Shanks, O.C.; Atikovic, E.; Blackwood, A.D.; Lu, J.; Noble, R.T.; Domingo, J.S.; Seifring, S.; Sivaganesan, M.; Haugland, R.A. Quantitative PCR for detection and enumeration of genetic markers of bovine fecal pollution. Appl. Environ. Microbiol. 2008, 74, 745–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ervin, J.S.; Van De Werfhorst, L.C.; Murray, J.L.; Holden, P.A. Microbial source tracking in coastal California watershed reveals canines as controllable sources of fecal contamination. Environ. Sci. Technol. 2014, 48, 9043–9052. [Google Scholar] [CrossRef] [PubMed]
- Ibekwe, A.M.; Murinda, S.E.; Graves, A.K. Genetic diversity and antimicrobial resistance of Escherichia coli from human and animal sources uncovers multiple resistances from human sources. PLoS ONE 2011, 6, e20819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaak, H.; Lynch, G.; Italiaander, R.; Hamidjaja, R.A.; Schets, F.M.; de Roda Husman, A.M. Multidrug-resistant and extended spectrum beta-lactamase-producing Escherichia coli in Dutch surface water and wastewater. PLoS ONE 2015, 10, e0127752. [Google Scholar] [CrossRef] [Green Version]
- Ramírez Castillo, F.Y.; Avelar González, F.J.; Garneau, P.; Márquez Díaz, F.; Guerrero Barrera, A.L.; Harel, J. Presence of multidrug resistant pathogenic E. coli in the San Pedro River located in the State of Aguascalientes, Mexico. Front. Microbiol. 2013, 4, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappell, A.D.; DeNies, M.S.; Ahuja, N.H.; Ledeboer, N.A.; Newton, R.J.; Hristova, K.R. Detection of multi-drug resistant Escherichia coli in the urban waterways of Milwaukee, WI. Front. Microbiol. 2015, 29, 336. [Google Scholar] [CrossRef] [Green Version]
- Harmel, R.D.; Richardson, C.W.; King, K.W.; Allen, P.M. Runoff and soil loss relationships for the Texas Blackland Prairies ecoregion. J. Hydro. 2006, 331, 471–483. [Google Scholar] [CrossRef]
- Harmel, D.; Wagner, K.; Martin, E.; Smith, D.; Wanjugi, P.; Gentry, T.; Gregory, L.; Hendon, T. Effects of field storage method on E. coli concentrations measured in storm water runoff. Environ. Monit. Assess. 2016, 188, 170. [Google Scholar] [CrossRef]
- Harmel, R.D.; Haney, R.L.; Smith, D.R.; White, M.; King, K.W. USDA-ARS Riesel Watersheds, Riesel, Texas, USA: Water quality research database. Water Resour. Res. 2014, 50, 8374–8382. [Google Scholar] [CrossRef]
- USEPA (2005). Method 1603: Escherichia coli (E. coli) in Water by Membrane Filtration Using Modified Membrane-Thermotolerant Escherichia Coli Agar (Modified mTEC); Environmental Protection Agency: Washington, DC, USA, 2005. [Google Scholar]
- Bauer, A.W.; Kirby, W.M.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Path. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Meth. 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Versalovic, J.; Koeuth, T.; Lupski, J.R. Distribution of repetitive DNA-sequences in eubacteria and application to fingerprinting of bacterial genomes. Nucl. Acids Res. 1991, 19, 6823–6831. [Google Scholar] [CrossRef] [PubMed]
- Casarez, E.A.; Pillai, S.D.; Mott, J.B.; Vargas, M.; Dean, K.E.; Di Giovanni, G.D. Direct comparison of four bacterial source tracking methods and use of composite data sets. J. Appl. Microbiol. 2007, 103, 350–364. [Google Scholar] [CrossRef] [PubMed]
- Brooks, J.P.; McLaughlin, M.R. Antibiotic resistant bacterial profiles of anaerobic swine lagoon effluent. J. Environ. Qual. 2009, 38, 2431–2437. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Gonzalez, N.; Porrero, M.C.; Mentaberre, G.; Serrano, E.; Mateos, A.; Domínguez, L.; Lavín, S. Antimicrobial resistance in indicator Escherichia coli isolates from free ranging livestock and sympatric wild ungulates in a natural environment (Northeastern Spain). Appl. Environ. Microbiol. 2013, 79, 6184–6186. [Google Scholar] [CrossRef] [Green Version]
- Janezic, K.J.; Ferry, B.; Hendricks, E.W. Phenotypic and genotypic characterization of Escherichia coli isolated from untreated surface waters. Open Microbiol. J. 2013, 7, 9–19. [Google Scholar] [CrossRef]
- Laird, D. Characterization of Antimicrobial Resistance Profiles of Surface Water Bacteria in an Urbanizing Watershed. Mastre’s Thesis, Texas A&M University, College Station, TX, USA, 2016. Available online: https://oaktrust.library.tamu.edu/handle/1969.1/157788 (accessed on 27 April 2020).
- Holvoet, K.; Sampers, I.; Callens, B.; Dewulf, J.; Uyttendaele, M. Moderate prevalence of antimicrobial resistance in Escherichia coli isolates from lettuce, irrigation water, and soil. Appl. Environ. Microbiol. 2013, 79, 6677–6683. [Google Scholar] [CrossRef] [Green Version]
- Nicolau, D.P.; Carmeli, Y.; Crank, C.W.; Goff, D.A.; Graber, C.J.; Lima, A.L.L.; Goldstein, E.J.C. Carbapenem stewardship: Does ertapenem affect Pseudomonas susceptibility to other carbapenems? A review of the evidence. Int. J. Antimicrob. Agents 2012, 39, 11–15. [Google Scholar] [CrossRef]
- Maal-Bared, R.; Bartlett, K.H.; Bowie, W.R.; Hall, E.R. Phenotypic antimicrobial resistance of Escherichia coli and E. coli O157 isolated from water, sediment, and biofilms in an agricultural watershed in British Columbia. Sci. Tot. Environ. 2013, 443, 315–323. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mukherjee, M.; Gentry, T.; Mjelde, H.; Brooks, J.P.; Harmel, D.; Gregory, L.; Wagner, K. Escherichia coli Antimicrobial Resistance Variability in Water Runoff and Soil from a Remnant Native Prairie, an Improved Pasture, and a Cultivated Agricultural Watershed. Water 2020, 12, 1251. https://doi.org/10.3390/w12051251
Mukherjee M, Gentry T, Mjelde H, Brooks JP, Harmel D, Gregory L, Wagner K. Escherichia coli Antimicrobial Resistance Variability in Water Runoff and Soil from a Remnant Native Prairie, an Improved Pasture, and a Cultivated Agricultural Watershed. Water. 2020; 12(5):1251. https://doi.org/10.3390/w12051251
Chicago/Turabian StyleMukherjee, Maitreyee, Terry Gentry, Heidi Mjelde, John P. Brooks, Daren Harmel, Lucas Gregory, and Kevin Wagner. 2020. "Escherichia coli Antimicrobial Resistance Variability in Water Runoff and Soil from a Remnant Native Prairie, an Improved Pasture, and a Cultivated Agricultural Watershed" Water 12, no. 5: 1251. https://doi.org/10.3390/w12051251
APA StyleMukherjee, M., Gentry, T., Mjelde, H., Brooks, J. P., Harmel, D., Gregory, L., & Wagner, K. (2020). Escherichia coli Antimicrobial Resistance Variability in Water Runoff and Soil from a Remnant Native Prairie, an Improved Pasture, and a Cultivated Agricultural Watershed. Water, 12(5), 1251. https://doi.org/10.3390/w12051251