Abstract
The most important environmental characteristic in streams is flow. Due to the force of water current, most ecological processes and taxonomic richness in streams mainly occur in the riverbed. Benthic algae (mainly diatoms) and benthic macroinvertebrates (mainly aquatic insects) are among the most important groups in running water biodiversity, but relatively few studies have investigated their complex relationships. Here, we review the multifaceted interactions between these two important groups of lotic organisms. As the consumption of benthic algae, especially diatoms, was one of the earliest and most common trophic habits among aquatic insects, they then had to adapt to the particular habitat occupied by the algae. The environmental needs of diatoms have morphologically and behaviorally shaped their scrapers, leading to impressive evolutionary convergences between even very distant groups. Other less evident interactions are represented by the importance of insects, both in preimaginal and adult stages, in diatom dispersion. In addition, the top-down control of diatoms by their grazers contributes to their spatial organization and functional composition within the periphyton. Indeed, relationships between aquatic insects and diatoms are an important topic of study, scarcely investigated, the onset of which, hundreds of millions of years ago, has profoundly influenced the evolution of stream biological communities.
1. Introduction
“The substratum is the stage upon which the drama of aquatic insect ecology is acted out”: this famous sentence of Minshall [1] is still relevant today, and can be extended to the essence of running water biota. In fact, while in lentic environments the water column hosts a rich and diverse community of producers (i.e., phytoplankton) and consumers (i.e., zooplankton and larger organisms), the effect of flow almost eliminates this habitat in lotic systems. Flow is the environmental factor that conditions life in lotic systems the most, exerting an incessant and unidirectional force on everything exposed to moving water [2]. Therefore, the majority of organisms that inhabit running water habitats belong to the benthic community, i.e., they are strictly associated with the stream bed. In this context, benthic algae and benthic macroinvertebrates represent the most important elements, but there are very few comprehensive studies that consider the different relationships between these two groups.
Almost all surfaces that receive light on river bottoms are covered with aquatic algae, which can be separated into two major non-taxonomic groups, macro- and micro-algae. The first group includes species whose thallus is visible to the naked eye, while the second group includes the smaller species. The latter is the most diverse and rich and, in this context, diatoms represent the ecologically most relevant group, reflecting their widespread occurrence and ecological success [3]. Diatoms are the most diverse group of algae in fresh waters and can colonize almost all aquatic habitats, although green algae and cyanobacteria can be very abundant under certain conditions. Filamentous green algae, for example, tend to prevail in lentic or slow water environments stressed by eutrophication, acidification and metal contamination [3]. The unique life cycle that characterizes diatoms, coupled with their particular wall structure and cell division mechanism [4], has probably played an important role in the evolutionary success of this group of algae. In the benthic habitats, primary producers are the principal component of the complex matrix called periphyton. Periphyton stoichiometry, i.e., C:N:P ratios, greatly influences grazer growth rate and the acquisition of energy reserves. Indeed, benthic algae represent a great energy source and high-quality food for macroinvertebrates due to their low C:N and C:P ratios and high protein and lipid contents [5]. Among the autotrophic groups composing the periphyton, diatoms are considered high quality food for macroinvertebrates compared to green algae [6] and cyanobacteria, due to their high content in long chain polyunsaturated fatty acids [7]. Beside stoichiometry, cyanobacteria represent a low-quality food source for grazers for several other reasons, such as the possession of toxins and secondary metabolites (for example protease inhibitors), their morphology (filamentous and/or colonial) and the lack of essential dietary lipids (see [8] and references therein).
Periphyton can be organized into different layers, whose spatial architecture approximates the vertical organization of terrestrial forests at a microscopic scale. In this context, diatoms are classified into growth forms according to their different positions in the biofilm, which in turn depend on their attachment mode to the substrate. At the basal level, we find adnate forms closely attached to the substrate through the entire valve surface or girdle bands. In the intermediate level, pad attached and stalked forms prevail. They first stick to the substrate by the mucilage excreted through the apical pore field located at one or both poles; the stalked forms produce a mucilage stalk that can be simple or branched. Within the periphyton, another important group of diatoms is represented by motile taxa; these are species able to move both within the biofilm and among cobbles towards the most suitable environmental conditions. Finally, on the upper layer we find overstory diatoms loosely attached, or unattached to the substrate. The growth form determines much of the diatom ecology, including how they interface with the primary consumers, discussed below.
Benthic macroinvertebrates represent a well-known group, widely used in biomonitoring, whose communities consist of species mainly belonging to Tricladida, Annelida, Crustacea, Mollusca, Nematoda, Nematomorpha, and Insecta. Insects represent the dominant component in lotic invertebrate communities because of their species richness, diversity, abundance, distribution and ubiquity [9]; 60.4% of the 126,000 worldwide known freshwater animal species are insects [10]. Likewise, aquatic insects are one of the most important elements of the lotic food webs, acting as primary and secondary consumers, pivotal in the processing and cycling of nutrients, producing a considerable amount of biomass to the ecosystem by means of their high secondary production, i.e., generating biomass [11,12]. They have also an important role linking aquatic food webs with terrestrial food webs by means of their flying activity during the adult stage of life.
Considering that diatoms and insects represent the most taxonomically rich, diverse and abundant groups among freshwater benthic algae and invertebrates, we focused on these two groups. Here, we review the complex interactions between the two most important groups of lotic organisms, namely benthic diatoms and macroinvertebrates, to try to shed light on the importance of these interactions.
The most obvious interaction between stream macroinvertebrates and diatoms is grazing, performed by herbivorous invertebrates on benthic biofilm. According to the classification of macroinvertebrates in functional feeding groups (FFGs; Table 1), the grazers (also called scrapers) include macroinvertebrates that feed on periphyton on hard surfaces [13].

Table 1.
Classification of the functional feeding groups (FFGs).
Although many stream invertebrates have considerable plasticity in the foods they ingest and can consume periphyton at some part of their lives [14], we found specialized scrapers in 6 out of 13 orders of insects with aquatic species not particularly related among them from a phylogenetic point of view. This shows that this trophic strategy has evolved several times, independently, in running water environments.
For those groups that colonized fresh waters originating from saltwater environments (e.g., gastropod molluscs, such as Lymnaeidae, Physidae) scraping is a simple transposition of something that occurred in marine ancestors. Nevertheless, the situation is different for aquatic insects, the dominant group of river macroinvertebrates, in terms of both taxonomic [10] and trophic [15] diversity.
It is generally believed that the first insects evolved in terrestrial environments and subsequently colonized inland waters, although some discordant hypotheses have emerged [16]. The first aquatic insects probably date back to the Permian [9]. Several groups of insects independently invaded aquatic habitats [17], so that truly aquatic insects include the most primitive winged forms (Ephemeroptera) and other ancient Paleoptera (Odonata), but also Neoptera Exopterygota (Plecoptera) and Endopterygota (Trichoptera and Megaloptera). In addition to this, typically aquatic taxa are present in about eight other insect orders, such as Coleoptera, Diptera, Lepidoptera and Hemiptera [9]. Most of these groups comprise scrapers. Moreover, even some taxa usually considered as predators can behave as mainly diatom consumers in some particular habitats, such as temporary streams [18].
In this context, two elements emerge as underlining the strict and ancient relationship between aquatic insects and benthic algae.
The first element is antiquity. The first aquatic insects are generally believed to have been predators feeding on a broad range of invertebrates. The second trophic habit that appeared was benthic periphyton-scraping, in the middle of the Permian period [19]. This hypothesis comes from the analysis of trophic resources available in ancient lotic systems (see [20]). Firstly, we have no fossil traces of aquatic macrophytes before the Triassic; moreover, macrophytes do not represent a primary food source for benthic invertebrates and, in particular, for insects [21]. Furthermore, it is well known that an important part of the energy input in current lotic systems derives from allochthonous non-living coarse particulate organic matter (CPOM), mainly terrestrial leaves [22]. However, this resource, consumed by shredders, was probably scarcely available in the past because ancient catchments were barely vegetated, and CPOM only assumed a certain importance in the early Cretaceous period, with the spread of angiosperms [20]. Consequently, the availability of fine organic particles (FPOM), originating from CPOM breakdown, was also probably low, explaining the late advent of filterers [23]. It can therefore be hypothesized that, for a very long period, the direct consumption of periphyton was one of the earliest, most important and common trophic habits among lotic invertebrates, and in particular among insects.
The second element is the selective pressure. As mentioned before, a second interesting aspect underlining the important relationship between benthic algae and aquatic insects is that scraping appeared several times, independently, in different and even very phylogenetically distant insect groups. Scrapers can be found in six orders of insects and for some of these, feeding on algae represents probably one of the most important ways of life [23]. In the most ancient winged group, the Ephemeroptera, grazers are very common in many families, such as Heptageniidae, Siphlonuridae, Leptophlebiidae, Caenidae and Baetidae [24]. In addition, many Plecoptera feed on periphyton, such as some Diamphipnoidae, Gripopterygidae, Notonemouridae, Capniidae and Taeniopterygidae [25,26]. Among Hemiptera, some Corixidae are largely herbivorous, scraping algae from submerged vegetation and stone [9,27]. Among Endopterygota, scrapers are also very common, for example, Coleoptera Hydraenidae, Psephenidae, Helodidae/Scirtidae, Dryopidae and Elmidae feed on algae in the preimaginal and/or adult stage [13]. Several families of Trichoptera are almost exclusively scrapers, such as Glossosomatidae, Helicopsychidae, Apataniidae, Goeridae and Uenoidae; others include many taxa with this trophic habit, such as Hydroptilidae and Leptoceridae, while some other taxa are facultative scrapers, such as Psychomyiidae, Hydropsyche (Hydropsychidae) and Marila (Odontoceridae) [28]. Among Diptera, according to [29], larval Blephariceridae, Deuterophlebiidae, some Psychodidae, Ephydridae and Chironomidae are scrapers. In practice, all these organisms, even very distant from a phylogenetic point of view, have undergone the same evolutionary pressures to adapt to the scraper diet, and have therefore evolved some similar characteristics in a convergent way.
What are these characteristics? Or rather, how did algae shape their consumers? We must first consider that the development of periphyton is not homogeneous in all river microenvironments, being favored by some specific environmental conditions, such as good sunlight exposure (which facilitates the photosynthetic process), medium to high current velocity (which prevents sedimentation and thus protects algae from burial) and, in general, coarse substrata (which ensures better immovability over time and a more stable colonization) [30].
In scrapers belonging to different insect groups, natural selection has favored the development of similar morphological and behavioral adaptations, which can be summarized as follows.
2. Ability to Withstand Elevated Velocity Current Environments
Compared to shredders or predators, lotic grazers are exposed to higher hydraulic stress, as their food sources colonize the top of boulders and pebbles due to their light requirements. Therefore, the general adaptations presented by lotic aquatic insects to the high flow speed must be more developed and more sophisticated in scrapers.
Different convergent strategies have been developed, aimed to minimize the threat of drifting downstream by the current. These essentially include adopting a hydrodynamic profile, having powerful adhesion structures (nails, suction cups, silk) or, in some cases, weighing down the body. Among aquatic insects, the most impressive examples of hydrodynamic shape are found in scrapers. Since the pioneering studies of Statzner and collaborators [31], the dorsoventral flatness of Heptageniidae (Ephemeroptera) is believed to be the key that enabled these animals to colonize even the fastest environments, taking shelter in the lowest, thick and viscous portion of the boundary layer, where flow is slowest and laminar (Figure 1). In addition, immature stages of Diptera Deuterophlebiidae, typical scrapers that inhabit riffle habitats where current velocities usually exceed 1 m/s, are noticeably dorsoventrally flattened [32]. The same adaptation has occurred several times in the evolutionary history of aquatic insects, reaching the most spectacular forms among Coleoptera, where the larvae of Elmidae and Psephenidae have unique dimensional relationships, with very flattened, shield- to disc-shaped body forms [17]. In addition to dorsoventral flattening, mayflies and beetles, despite being distant in the evolutionary tree of hexapods, have adopted the same solutions even in smaller details, namely the general body form (broader at the front and narrow behind), the presence of smooth lateral structures helping the hydrodynamic profile (i.e., femura and headparts in heptagenids, lateral plate-like extensions in Coleoptera larvae) and the existence of marginal hair fringes that increase the adhesion capacity to the substrate.

Figure 1.
Dorsoventral flattening of an Ecdyonurus (Ephemeroptera: Heptageniidae) nymph (Photo Roberto Messori; reproduced with permission from the author).
Moreover, to live in fast-flowing zones with high hydraulic constraints, scrapers have developed very effective adhesion structures. The tarsal claws of rheophilic Ephemeroptera, Plecoptera, Coleoptera, Diptera and Trichoptera are usually extremely robust, sharp and curved to better interlock with the surface irregularities of the substrate [17]. Some Heptageniidae, such as Epeorus assimilis, can cling with their claws to even slightly rough substrates, but Ditsche-Kuru and collaborators [33] found that the presence of biofilm considerably increases the adhesion ability of insects. Other attachment devices are represented by circlets of small hooks in the pseudopoda of Deuterophlebiidae and powerful ventral suckers of Blephariceridae (Figure 2).
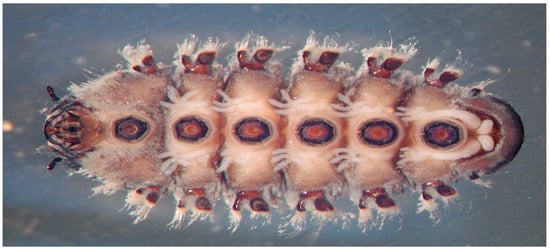
Figure 2.
Ventral suckers of a Blephariceriidae larva (Photo Stefano Fenoglio).
3. Ability to Scrape Algae from the Substrate
Lotic algae, and in particular diatoms, are known to have strong adhesion ability to hard substrates [34,35]. To feed on these organisms, different insect groups have had to adopt similar morphological adaptations, in particular regarding mouthpart morphology. Arens [36] reported that all scrapers had to find a solution to solve four main problems. Firstly, they must be able to detach the algae from the substrate. Secondly, once removed from the substrate, algae must be collected and transferred to the oral cavity. For example, some Heptageniidae mayflies exhibit adapted labial and maxillary palps combining reinforced scraping parts and setose brush parts [37]. Among Plecoptera, Brachypterinae have maxillae combining chisel-like structures in the lacinia and brush-like structures in the galea [17]. Among Trichoptera, the mandibles of Glossosomatidae are elongated with edge and broom structures [28]. In scraping beetles, mandibles can present a sharp, anterior incisor lobe, while galea or lacinia can show a brush apparatus. Thirdly, the diatom siliceous shell can be a mechanical protection against grazing that grazers need to overcome, although its defensive effectiveness is not supported by much experimental evidence [38]. Finally, all this mouthpart activity must be carried out in the shelter of the current. It is impressive how, starting from different morphological bases, all insect scrapers have converged in adopting very similar solutions; for more details, see the comparison made by Arens [39] of scraper mouthparts using SEM images. Lastly, this feeding mode leads to another interesting convergence among aquatic insects, as scraper mouthparts are subjected to strong abrasion and wear out quickly. These organisms have thus had to evolve specific adaptations, or use some previously existing ones, such as the exoskeleton moult in arthropods, to favor renewal of mouthparts and prolong their duration (see details in [36]).
In reviewing the aquatic insects–periphyton relationships, Lamberti and Moore [40] wondered if taxon-specific preferences between grazers and attached algae exist and to what extent this selectivity can be considered as an active or passive selection. After decades of field and mesocosm experiments, the scientific literature shows evidence for a certain degree of selectivity by scrapers but, generally, freshwater ecologists agree upon the fact that this selectivity is mainly a passive consequence of the interplay between the size and morphology of the insect mouthparts and the algal susceptibility to grazing according to their life forms [41,42,43]. Indeed, as already mentioned before, pad attached (such as Meridion) or stalked (such as Cymbella and Gomphonema) diatoms are more susceptible to grazing pressure than small adnate forms (such as Achnanhtidium) [41].
4. Adaptation to Live in The Open
Most benthic invertebrate species live hidden among the elements of the river bottom, between the detritus, under the large boulders, in the interstices between the pebbles, or even burrowed in the sand, where they find food and shelter from predators. However, this strategy cannot be adopted by scrapers, which must spend a lot of time “above” and not “below” the rocks to feed. For this reason, when wading a stream, insect scrapers are the most easily observable: for example, in an alpine creek, the nymphs of Heptageniidae are commonly visible on the upper surface of boulders, while dense populations of Blephariceridae stand out on large rocks with fast flow. It is the very nature of their microhabitat (and therefore their food strategy) that protects them from predators: in fact, predators select the most advantageous prey according to different factors, such as prey density, energy contents, handling time and encounter rate [44], and this last element greatly reduces the vulnerability of scrapers. For example, analyzing the diet of two large-sized predaceous Systellognatha Plecoptera, Bo et al. [45] found that Blephariceridae, even if present in the same river stretch, were never consumed, probably because they were too difficult to reach. Moreover, in another study focused on feeding preferences of predaceous stoneflies, using trophic electivity indices to compare gut contents with natural composition and abundance of macroinvertebrate community in the riverbed, Heptageniidae, as a food source, was always negatively selected [46]. Interestingly, we also report that the few non-scraper insects inhabiting the same microhabitats (upper surface of masses and rocks in very fast-flowing waters), such as Diptera Simuliidae, are only included in the predator diet in exceptional cases [46].
5. Direct and Indirect Effects of Scrapers in Shaping Diatom Communities
The relationship between scrapers and diatoms can also be viewed from the other side: how do scrapers shape the diatom community as an effect of herbivory? Did diatoms evolve defense mechanisms against herbivory? Grazing in freshwater benthic ecosystems was essentially unexplored until the 1980s when Gregory [47] elaborated the theory of a top-down control of primary producers by grazers. In the book Algal Ecology, Stevenson et al. [48] devoted an entire chapter to the regulative action on benthic algae played by grazers. In most cases, periphyton is considered to be affected by both bottom-up (nutrients, light and other abiotic factors) and top-down controls [49], but their respective role can vary upon additional conditions. Graça and collaborators [49] performed an experimental study in a low order tropical stream in enclosures under controlled conditions of nutrients and light. Given the low grazer density, they expected a low grazing pressure. Conversely, they found an intense grazing effect, more evident in semi-shaded areas (53% algal biomass removed) than in fully shaded ones (33%). Moreover, grazing resulted in a decrease of diatoms in favor of cyanobacteria, which are considered more tightly adhered in some cases (e.g., Chamaesiphonales), less palatable and lacking in high-saturated fatty acids.
Herbivores can frequently alter periphyton composition and physiognomy as they more easily consume overstory or loosely-attached algae [50], favoring a shift towards tightly attached prostrate forms. Despite this, a recent study highlighted that herbivorous abundance apparently does not significantly affect diatom ecological guild structure, and that grazing pressure has no effects on diatom ecological guild diversity [51]. Besides possible changes in community composition, the overall biomass can be almost unaltered by grazing pressure. In fact, the ability of grazers to induce a significant biomass loss depends, among other reasons, on their density, as shown in classical mesocosm studies [52].
Given the complex multilayer structure of periphyton and the different feeding strategies adopted by grazers, it is reasonable to hypothesize preferential feeding based on resource partitioning. Resource partitioning is a central concept in community ecology as it may explain the coexistence of species belonging to the same guild. It has been confirmed for terrestrial vertebrates, especially birds [53] but hardly associated with benthic grazers, which are generally considered as largely generalists. In a recent study, Piano et al. [54] examined the distribution of three Heptagenidae taxa, namely Rhitrogena, Epeorus and Ecdyonurus, commonly found in Alpine streams. Their findings suggested that the distribution of the first two genera was strictly linked to diatom biomass, whereas this relationship was not so evident for Ecdyonurus, which has a more generalist diet including allochthonous detritus. Tall et al. [55] analyzed the food preferences of grazers feeding on epiphytic diatoms growing on the moss Fontinalis. Their findings suggest that when available food is reduced there is a resource partitioning within the grazers. Based on their results, they divided the grazers into three categories: (1) true scrapers, such as the coleopteran Promoresia and the caddisfly Hydroptila, which fed selectively on the adnate diatoms; (2) generalists (e.g., Baetis and some Chironomidae), without any clear preference regarding the biofilm layer; (3) surfers (e.g., some Chironomidae), which avoided the adnate diatoms and showed a preference for detached cells. In this repartition, mouthpart morphology has a great importance [56].
In addition to the direct effects of grazing, aquatic insects that feed on attached algae can also affect the composition and biomass of periphyton indirectly. For instance, the action of the grazers may reduce the self-shading of biofilms and, in turn, stimulate the cell growth and species turnover within the periphytic layer [57]. Moreover, grazers can indirectly affect diatom composition by favoring the dispersal of certain taxa. Indeed, Peterson [41] observed that after gut passage, an important percentage of ingested diatom cells stay viable when eliminated through feces (about 40–52%). The physical dislodgement of diatoms by grazers can, therefore, significantly contribute to diatom dispersal, especially in the downstream stretches. Another indirect and beneficial effect attributed to grazers is the nutrient enrichment due to the residuals of ungrazed algae as well as grazers’ feces. In a mesocosm experiment, Herren et al. [58] found that the area-specific primary production of attached algae was 71% higher in the presence of residential Chironomidae larvae than in their absence, probably because of the consumers’ fertilizing action. These findings suggest that the relationship between insect scrapers and attached algae is a complex trade-off between positive (i.e., beneficial) and negative (i.e., adverse) interactions [57].
The occupation of different layers within the periphyton and, in particular, the basal one can be seen as a strategy to resist the action of grazing. Furthermore, the siliceous frustule could have a defensive function, though its anti-grazing role has been questioned in some articles, almost all related to marine planktonic environment. Hamm et al. [59] started from the consideration that its role is not decisive as diatoms are an elective food for a large variety of organisms. Nevertheless, they highlighted that in an environment characterized by a high grazing pressure, such as the planktonic one, the presence of a mechanical protection should play a role in reducing diatom population mortality and in shaping phyto- and zoo-plankton populations. Pančić et al. [38] experimentally demonstrated that the frustule defensive power of marine phytoplanktonic diatoms varies according to different grazing modes. Calanoid copepods have robust mandibles that can crack diatom frustules before ingestion, with a success rate that depends on the silica content of the frustule, which can vary among diatom species. The frustule does not provide any protection against grazing by protozoans, which engulf their prey and digest the cell content without breaking the valves. Despite the paucity of similar studies in freshwater benthic environments, we can confirm that aquatic insects often digest the diatom content without destroying the frustules. In fact, complete diatom frustules are usually found in aquatic insect gut contents, and even intact but empty frustules are found in the insect fecal material, together with other diatom individuals that have survived the passage through the digestive system [18,60].
Producing harmful secondary metabolites as anti-predatory mechanisms is a complex and interesting issue that must still be unraveled. A few diatom taxa are known to produce a toxic amino acid, domoic acid, which causes serious gastrointestinal and neurological consequences in humans and aquatic animals [61]. These are the marine genus Pseudonitzschia [61] and the estuarine species Amphora coffeaeformis [62]. More recently, Violi et al. [63] reported the production of other toxic amino acids, e.g., β-methylamino-L-alanine and its isomer 2,4-diaminobutyric acid, by several freshwater diatoms under certain culture conditions, formerly only attributed to Cyanobacteria. They hypothesized that diatoms could produce these toxins as a response to stress such as nutrient depletion. As suggested by this study, production of these toxins in freshwater environments may become a health issue in freshwater diatom blooms, although the effect on aquatic insects requires further research to be assessed.
On the other hand, the effects that massive blooms of some benthic diatoms have in modifying the habitat for river communities are quite well known. In this regard, an emblematic example is represented by the blooms of Didymosphenia geminata. This is the most studied invasive diatom [64], which under certain environmental conditions produces large quantities of extracellular stalks that can almost completely cover more or less long stretches of river, sometimes reaching an extension of a few kilometers in length [65]. The blooms of D. geminata can profoundly alter the invaded benthic communities, by decreasing β-diversity and increasing taxonomic homogenization in both algal and invertebrate assemblages [66]. The thick filament mats prevent the movements of the larger aquatic insects, such as the Heptageniidae, favoring instead smaller, opportunistic and generally herbivorous organisms, such as the chironomids and oligochaetes [65,66]. When in bloom, the D. geminata filamentous mats cover almost completely hard substrates, disadvantaging crawlers, shredders and scrapers. The chironomids take advantage of the absence of large competitors and predators, and a significant change in top-down control is therefore determined because the chironomids prefer the larger diatoms, favoring in turn the smaller and pioneering taxa. Even though D. geminata stalks can provide a suitable substrate for other diatoms (such as Achnanthidium spp.), contributing to changes in diatom community composition, we found no significant evidence that this could lead to advantages or disadvantages on macroinvertebrate communities. Overall, D. geminata blooms lead to a simplification of the food web structure with a dominance of smaller organisms.
6. Epizoic Relationships
In some instances diatoms take advantage of the relationship with macroinvertebrates, as in the case of the epibiosis. The term “epibiosis” describes a strict association between organisms, in which we can identify an epibiont (the organism that lives attached to the body surface of another organism) and a basibiont (the organism that hosts epibionts [67]). Even if is not parasitic, this relationship is often unbalanced because the epibiont has the greatest advantages. Here we review the relationship between diatoms (as epibionts) and aquatic insects (as basibionts) in running water ecosystems.
Epibionts, such as benthic freshwater diatoms, have some obvious advantages in colonizing the surfaces of aquatic insects, including: (i) an enhanced protection against grazing; (ii) an increased accessibility to solar radiation, carbon dioxide and nutrients, because of the basibiont activity and metabolism; (iii) a better ability to avoid burial by fine sediments; and (iv) an important benefit for dispersion [68]. Benefits for basibionts are less obvious; some studies report that the presence of an epibiont biofilm can be useful to increase camouflage and reduce irradiation, while others emphasize how this layer can impair numerous functional processes (such as gas exchange) and limit the possibility of movement [69]. For these reasons, epizoic (i.e., living on an animal) diatoms are probably more common and diverse than previously supposed. To date, most studies on epizoic diatoms have been performed in marine environments [70] and, of the studies carried out in fresh water, many have focused on non-insect taxa. Freshwater diatoms have been found on Testudines [71], Gastropoda Physidae [72], Rotifera [73], Copepoda [74] and, in particular, Crustacea Decapoda. Due to their generally large size and hard exoskeletons, freshwater crabs and crayfish ideally represent optimal basibionts for diatoms and other microalgae [68,75].
Although there are not many studies on the diatom–insect epibiosis, we can still underline how this relationship has a considerable and probably underestimated importance, especially regarding the role of aquatic insects in algal dispersion. Very few studies have focused on diatoms living on preimaginal stages of aquatic insects. Larval cases of some Trichoptera are known to host algae [76], but mechanisms regulating epibiotic associations between diatoms and aquatic stages are still practically unknown; not all aquatic insects nor all diatom species show this kind of association. For example, Wujek [77], using scanning electron microscopy, reported that three species of diatoms, among the numerous present in the substrate of Cedar Creek (MI, USA), lived on the cerci of Caenidae nymphs, while none were found on the nymphs of sympatric Ephemerellidae. Further studies need to be carried out regarding pre-imaginal stages, while we have more information about epizoic diatoms on adult, winged organisms. In fact, most aquatic insects have an “amphibious” life-cycle, characterized by the presence of a pre-imaginal aquatic stage and an imaginal terrestrial stage. Contact between diatoms and adult insects may occur during emergence from nymphal or pupal exuvia, or during oviposition, when females (and occasionally also males, e.g., in some Odonata) come into contact with surface water or even submerge to lay eggs in the vegetation or substrate, or when resting in the wet areas on the banks of the stream or on the boulders reached by the splashes of water. For example, Stewart and Schlichting [78] reported the presence of diatoms on the exoskeleton of some Odonata Zygoptera and Anisoptera, Hemiptera Gelostocoridae, Diptera Chironomidae and Trichoptera Polycentropodidae collected with light-traps at night or picked from riverbanks and vegetation surrounding water. Another study reported that some diatoms (such as Navicula and Nitzschia) were found attached to adult aquatic Diptera belonging to Tipulidae and Ptychopteridae families [79].
This is interesting, because adult stages may represent an important element in the dispersion of diatoms, allowing algae to colonize new environments and to pass from river to river across land barriers.
7. Conclusions
In conclusion, relationships between aquatic insects and diatoms is a subject of great interest, which, over hundreds of millions of years, has profoundly marked the evolutionary path of biological communities in streams. In running water environments, these groups are truly among the most important organisms from different points of view, whose multiple and often still unknown relationships form the substrate at the base of entire communities and trophic chains. This review shows that the direct and indirect grazing behavior of aquatic insects acts as an important and multifaceted mechanism that affects the diversity, composition, growth rate and biomass of attached algae, along with the bottom-up processes. However, the effects of these biotic interactions in shaping benthic algae communities and ecosystem functionality have not been studied in-depth (but see [80,81]). A novel and challenging approach could be adopting the recent advances of the metacommunity theory [82], as biological communities are simultaneously shaped by the pure and combined effect of environmental filters, spatial variables and biotic interactions. Although the former two categories of predictors have been investigated a lot, few studies have attempted to quantify the latter category. In light of their strong evolutive and trophic relationship, we believe that attached algae and aquatic insect scrapers represent ideal organisms to better investigate the role of biotic interactions in shaping benthic metacommunities. Similarly, we believe that all the natural and human-induced variations in the distribution and density of scrapers, such as flow and thermal and sedimentary alterations, deserve greater attention for future studies because of their cascade effects on the attached algae.
Author Contributions
All authors contributed equally to this paper. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
This work was realized within the framework of the activities of ALPSTREAM, a research center financed by FESR, Interreg Alcotra 2014–2020, EcO Project of the Piter Terres Monviso. English language review supplied by Rhadika Srinivasan.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Minshall, G.W. Aquatic insect-substratum relationships. In The Ecology of Aquatic Insects; Resh, V.H., Rosenberg, D.M., Eds.; Praeger Publishers: New York, NY, USA, 1984; pp. 358–400. [Google Scholar]
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The natural flow regime. BioScience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Bellinger, E.G.; Sigee, D.C. Freshwater Algae: Identification, Enumeration and Use as Bioindicators; John Wiley Sons: London, UK, 2015; p. 290. [Google Scholar]
- Chepurnov, V.A.; Mann, D.G.; Sabbe., K.; Vyverman, W. Experimental studies on sexual reproduction in diatoms. Int. Rev. Cytol. 2004, 237, 91–154. [Google Scholar] [PubMed]
- Torres-Ruiz M., J.D.; Wehr, J.D.; Perrone, A.A. Trophic relations in a stream food web: Importance of fatty acids for macroinvertebrate consumers. J. North. Am. Benthol. Soc. 2007, 26, 509–522. [Google Scholar] [CrossRef]
- Guo, F.; Kainz, M.J.; Sheldon, F.; Bunn, S.E. The importance of high-quality algal food sources in stream food webs–current status and future perspectives. Freshw. Biol. 2016, 61, 815–831. [Google Scholar] [CrossRef]
- Guo, F.; Bunn, S.E.; Brett, M.T.; Fry, B.; Hager, H.; Ouyang, X.; Kainz, M.J. Feeding strategies for the acquisition of high-quality food sources in stream macroinvertebrates: Collecting, integrating, and mixed feeding. Limnol. Oceanogr. 2018, 63, 1964–1978. [Google Scholar] [CrossRef]
- Groendahl, S.; Fink, P. High dietary quality of non-toxic cyanobacteria for a benthic grazer and its implications for the control of cyanobacterial biofilms. BMC Ecol. 2017, 17, 20. [Google Scholar] [CrossRef]
- Merritt, R.W.; Wallace, J.B. Aquatic habitats. In Encyclopedia of Insects; Resh, V.H., Cardi, R.T., Eds.; Academic Press: Burlington, MA, USA, 2009; pp. 38–48. [Google Scholar]
- Balian, E.V.; Segers, H.; Martens, K.; Lévéque, C. The freshwater animal diversity assessment: An overview of the results. Hydrobiologia 2008, 595, 627–637. [Google Scholar] [CrossRef]
- Resh, V.H.; Rosenberg, D.M. The Ecology of Aquatic Insects; Praeger Publisher: New York, NY, USA, 1984; p. 625. [Google Scholar]
- Cummins, K.W.; Merritt, R.W. Ecology and distribution of aquatic insects. In An Introduction to the Aquatic Insects of North. America; Merritt, R.W., Cummins, K.W., Eds.; Kendall Hunt Publishing Company: Dubuque, IA, USA, 1996; pp. 74–86. [Google Scholar]
- Cummins, K.W. Trophic relations of aquatic insects. Annu. Rev. Entomol. 1973, 18, 183–206. [Google Scholar] [CrossRef]
- Hauer, F.R.; Lamberti, G.A. Methods in Stream Ecology; Academic Press: New York, NY, USA, 2011; p. 506. [Google Scholar]
- Fenoglio, S.; Merritt., R.W.; Cummins, K.W. Why do no specialized necrophagous species exist among aquatic insects? Freshw. Sci. 2014, 33, 711–715. [Google Scholar] [CrossRef]
- Pritchard, G.; McKee, M.H.; Pike, E.M.; Scrimgeour, G.J.; Zloty, J. Did the first insects live in water or in air? Biol. J. Linn. Soc. 1993, 49, 31–44. [Google Scholar] [CrossRef]
- Wichard, W.; Arens, W.; Eisenbeis, G. Biological Atlas of Aquatic Insects; Apollo Books: Aamosen, Denmark, 2002; p. 339. [Google Scholar]
- López-Rodríguez, M.J.; Tierno de Figueroa, J.M.; Fenoglio, S.; Bo, T.; Alba-Tercedor, J. Life strategies of 3 Perlodidae species (Plecoptera) in a Mediterranean seasonal stream in southern Europe. J. North. Am. Benthol. Soc. 2009, 28, 611–625. [Google Scholar] [CrossRef]
- Wooton, R.J. The historical ecology of aquatic insects: An overview. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1988, 62, 477–492. [Google Scholar] [CrossRef]
- Lancaster, J.; Downes, B.J. Aquatic Entomology; Oxford University Press: Oxford, UK, 2013; p. 296. [Google Scholar]
- Allan, J.D.; Castillo, M.M. Stream Ecology: Structure and Function of Running Waters, 2nd ed.; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Huryn, A.D. Aquatic insects—Ecology, feeding, and life history. In Encyclopedia of Inland Waters; Likens, G., Benbow, M.E., Burton, T.M., Van Donk, E., Downing, J.A., Gulati, R.D., Eds.; Academic Press: New York, NY, USA, 2009; pp. 25–50. [Google Scholar]
- Brittain, J.E.; Sartori, M. Ephemeroptera. In Encyclopedia of Insects; Resh, V.H., Cardé, R.T., Eds.; Academic Press: Burlington, MA, USA, 2009; pp. 328–333. [Google Scholar]
- Fenoglio, S.; Bo, T.; López-Rodríguez, M.J.; Tierno de Figueroa, J.M. Nymphal biology of Brachyptera risi (Morton, 1896) (Plecoptera: Taeniopterygidae) in a North Apennine stream (Italy). Entomol. Fenn. 2008, 19, 228–231. [Google Scholar] [CrossRef]
- Tierno de Figueroa, J.M.; López-Rodríguez, M.J. Trophic ecology of Plecoptera (Insecta): A review. Eur. Zool. J. 2019, 86, 79–102. [Google Scholar] [CrossRef]
- Haedicke, C.W.; Redei, D.; Kment, P. The diversity of feeding habits recorded for water boatmen (Heteroptera: Corixoidea) world-wide with implications for evaluating information on the diet of aquatic insects. Eur. J. Entomol. 2017, 114, 147–159. [Google Scholar] [CrossRef]
- Holzenthal, R.W.; Thomson, R.E.; Ríos-Touma, B. Order Trichoptera. In Thorp and Covich’s Freshwater Invertebrates: Ecology and General Biology; Thorp, J.H., Rogers, D.C., Eds.; Elsevier: London, UK, 2015; Volume 1, pp. 965–1002. [Google Scholar]
- Courtney, G.W.; Cranston, P.S. Diptera. In Thorp and Covich’s Freshwater Invertebrates: Ecology and General Biology; Thorp, J.H., Rogers, D.C., Eds.; Elsevier: London, UK, 2015; Volume 1, pp. 1043–1058. [Google Scholar]
- Biggs, B.J.F. Patterns in benthic algae of streams. In Algal Ecology: Freshwater Benthic Ecosystems; Stevenson, R.J., Bothwell, M.L., Lowe, R.L., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 31–56. [Google Scholar]
- Statzner, B.; Holm, T.F. Morphological adaptations of benthic invertebrates to stream flow—An old question studied by means of a new technique (Laser Doppler Anemometry). Oecologia 1982, 53, 290–292. [Google Scholar] [CrossRef]
- Courtney, G.W. Life history patterns of Nearctic mountain midges (Diptera: Deuterophlebiidae). J. North. Am. Benthol. Soc. 1991, 10, 177–197. [Google Scholar] [CrossRef]
- Ditsche-Kuru, P.; Barthlott, W.; Koop, J.H. At which surface roughness do claws cling? Investigations with larvae of the running water mayfly Epeorus assimilis (Heptageniidae, Ephemeroptera). Zoology 2012, 115, 379–388. [Google Scholar] [CrossRef]
- Higgins, M.J.; Crawford, S.A.; Mulvaney, P.; Wetherbee, R. Characterization of the adhesive mucilages secreted by live diatom cells using atomic force microscopy. Protist 2002, 153, 25–38. [Google Scholar] [CrossRef]
- Gebeshuber, I.C.; Thompson, J.B.; Del Amo, Y.; Stachelberger, H.; Kindt, J.H. In vivo nanoscale atomic force microscopy investigation of diatom adhesion properties. Mater. Sci. Tech. Lond. 2002, 18, 763–766. [Google Scholar] [CrossRef]
- Arens, W. Wear and tear of mouthparts: A critical problem in stream animals feeding on epilithic algae. Can. J. Zool. 1990, 68, 1896–1914. [Google Scholar] [CrossRef]
- McShaffrey, D.; McCafferty, W.P. Feeding behavior of Rhithrogena pellucida (Ephemeroptera: Heptageniidae). J. North. Am. Benthol. Soc. 1988, 7, 87–99. [Google Scholar] [CrossRef]
- Pančić, M.; Torres, R.R.; Almeda, R.; Kiørboe, T. Silicified cell walls as a defensive trait in diatoms. Proc. R. Soc. B Biol. Sci. 2019, 286, 20190184. [Google Scholar]
- Arens, W. Striking convergence in the mouthpart evolution of stream-living algae grazers. J. Zool. Syst. Evol. Res. 1994, 32, 319–343. [Google Scholar] [CrossRef]
- Lamberti, G.A.; Moore, J.W. Aquatic insects as primary consumers. In The Ecology of Aquatic Insects; Resh, V.H., Rosenberg, D.M., Eds.; Praeger: New Yok, NY, USA, 1984; pp. 164–195. [Google Scholar]
- Peterson, C.G. Gut passage and insect grazer selectivity of lotic diatoms. Freshwat. Biol. 1987, 18, 455–460. [Google Scholar] [CrossRef]
- Díaz Villanueva, V.; Albariño, R. Algal Ingestion and Digestion by Two Ephemeropteran Larvae from a Patagonian Andean Stream; Research update on Ephemeroptera & Plecoptera; University of Perugia: Perugia, Italy, 2003; pp. 468–475. [Google Scholar]
- Schmid-Araya, J.M.; Figueroa Hernández, D.; Schmid, P.E.; Drouot, C. Algivory in food webs of three temperate Andean rivers. Austral. Ecol. 2012, 37, 440–451. [Google Scholar] [CrossRef]
- Krebs, J.R. Optimal foraging: Decision rules for predators. In Behavioural Ecology: An Evolutionary Approach; Krebs, J.R., Davies, N.B., Eds.; Blackwell: Oxford, UK, 1978; pp. 23–63. [Google Scholar]
- Bo, T.; Fenoglio, S.; López-Rodríguez, M.J.; Tierno de Figueroa, J.M. Trophic behavior of two Perlidae species (Insecta, Plecoptera) in a river in southern Spain. Int. Rev. Hydrobiol. 2008, 93, 167–174. [Google Scholar] [CrossRef]
- Bo, T.; Fenoglio, S.; Malacarne, G. Diet of Dinocras cephalotes and Perla marginata (Plecoptera: Perlidae) in an Apennine stream (northwestern Italy). Can. Entomol. 2007, 139, 358–364. [Google Scholar] [CrossRef]
- Gregory, S.V. Plant-herbivore interactions in stream systems. In Stream Ecology. Application and Testing of General Ecological Theory; Barnes, J.R., Minshall, G.W., Eds.; Springer: Boston, MA, USA, 1983; pp. 157–189. [Google Scholar]
- Stevenson, R.J.; Bothwell, M.L.; Lowe, R.L.; Thorp, J.H. Algal Ecology: Freshwater Benthic Ecosystem; Academic press: Dubuque, MA, USA, 1996; p. 753. [Google Scholar]
- Graça, M.A.; Callisto, M.; Barbosa, J.E.L.; Firmiano, K.R.; França, J.; Júnior, J.F.G. Top-down and bottom-up control of epilithic periphyton in a tropical stream. Freshwat. Sci. 2018, 37, 857–869. [Google Scholar] [CrossRef]
- Holomuzki, J.R.; Feminella, J.W.; Power, M.E. Biotic interactions in freshwater benthic habitats. J. North. Am. Benthol. Soc. 2010, 29, 220–244. [Google Scholar] [CrossRef]
- Vilmi, A.; Tolonen, K.T.; Karjalainen, S.M.; Heino, J. Metacommunity structuring in a highly-connected aquatic system: Effects of dispersal, abiotic environment and grazing pressure on microalgal guilds. Hydrobiologia 2017, 790, 125–140. [Google Scholar] [CrossRef]
- Steinman, A.D.; McIntire, C.D.; Gregory, S.V.; Lamberti, G.A. Effects of irradiance and grazing on lotic algal assemblages. J. Phycol. 1989, 25, 478–485. [Google Scholar] [CrossRef]
- MacArthur, R.H. Population ecology of some warblers of north eastern coniferous forests. Ecology 1958, 39, 599–619. [Google Scholar] [CrossRef]
- Piano, E.; Doretto, A.; Falasco, E.; Gruppuso, L.; Fenoglio, S.; Bona, F. The role of recurrent dewatering events in shaping ecological niches of scrapers in intermittent Alpine streams. Hydrobiologia 2019, 841, 177–189. [Google Scholar] [CrossRef]
- Tall, L.; Cattaneo, A.; Cloutier, L.; Dray, S.; Legendre, P. Resource partitioning in a grazer guild feeding on a multilayer diatom mat. J. North. Am. Benthol. Soc. 2006, 25, 800–810. [Google Scholar] [CrossRef]
- Merritt, R.; Cummins, K.; Berg, M.N. An Introduction to the Aquatic Insects of North. America, 5th ed; Kendall Hunt: Dubuque, IA, USA, 2019; p. 498. [Google Scholar]
- Vadeboncoeur, Y.; Power, M.E. Attached algae: The cryptic base of inverted trophic pyramids in freshwaters. Annu. Rev. Ecol. Evol. S. 2017, 48, 255–279. [Google Scholar] [CrossRef]
- Herren, C.M.; Webert, K.C.; Drake, M.D.; Jake Vander Zanden, M.; Einarsson, Á.; Ives, A.R.; Gratton, C. Positive feedback between chironomids and algae creates net mutualism between benthic primary consumers and producers. Ecology 2017, 98, 447–455. [Google Scholar] [CrossRef]
- Hamm, C.E.; Merkel, R.; Springer, O.; Jurkojc, P.; Maier, C.; Prechtel, K.; Smetacek, V. Architecture and material properties of diatom shells provide effective mechanical protection. Nature 2003, 421, 841–843. [Google Scholar] [CrossRef]
- Fenoglio, S.; Bo, T.; Tierno de Figueroa, J.M.; Cucco, M. Nymphal growth, life cycle, and feeding habits of Potamanthus luteus (Linnaeus, 1767) (Insecta: Ephemeroptera) in the Bormida River, Northwestern Italy. Zool. Stud. 2008, 47, 185–190. [Google Scholar]
- Botana, L.M. Seafood and Freshwater Toxins. Pharmacology, Physiology and Detection; CRC Press: New York, NY, USA, 2000; p. 1215. [Google Scholar]
- Dhar, B.C.; Cimarelli, L.; Singh, K.S.; Brandi, L.; Brandi, A.; Puccinelli, C.; Marcheggiani, S.; Spurio, R. Molecular detection of a potentially toxic diatom species. Int. J. Environ. Res. Public Health 2015, 12, 4921–4941. [Google Scholar] [CrossRef]
- Violi, J.P.; Facey, J.A.; Mitrovic, S.M.; Colville, A.; Rodgers, K.J. Production of β-methylamino-L-alanine (BMAA) and its isomers by freshwater diatoms. Toxins 2019, 11, 512. [Google Scholar] [CrossRef]
- Falasco, E.; Bona, F. Recent findings regarding non-native or poorly known diatom taxa in north-western Italian rivers. J. Limn. 2013, 201, 35–51. [Google Scholar] [CrossRef]
- Ladrera, R.; Goma, J.; Prat, N. Effects of Didymosphenia geminata massive growth on stream communities: Smaller organisms and simplified food web structure. PLoS ONE 2018, 13, e0193545. [Google Scholar] [CrossRef]
- Bray, J.P.; Kilroy, C.; Gerbeaux, P.; Burdon, F.J.; Harding, J.S. Ecological processes mediate the effects of the invasive bloom-forming diatom Didymosphenia geminata on stream algal and invertebrate assemblages. Hydrobiologia 2020, 847, 177–190. [Google Scholar] [CrossRef]
- Wahl, M. Epibiosis. In Marine Hard Bottom Communities; Wahl, L., Ed.; Springer: Boston, MA, USA, 2009; pp. 61–72. [Google Scholar]
- Falasco, E.; Bo, T.; Ghia, D.; Gruppuso, L.; Bona, F.; Fenoglio, S. Diatoms prefer strangers: Non-indigenous crayfish host completely different epizoic algal diatom communities from sympatric native species. Biol. Inv. 2018, 20, 2767–2776. [Google Scholar] [CrossRef]
- Wahl, M.; Goecke, F.; Labes, A.; Dobretsov, S.; Weinberger, F. The second skin: Ecological role of epibiotic biofilms on marine organisms. Front. Microbiol. 2012, 3, 292. [Google Scholar] [CrossRef]
- Robinson, N.J.; Majewska, R.; Lazo-Wasem, E.A.; Nel, R.; Paladino, F.V.; Rojas, L.; Zardus, J.D.; Pinou, T. Epibiotic diatoms are universally present on all sea turtle species. PLoS ONE 2016, 11, e0157011. [Google Scholar] [CrossRef]
- Wetzel, C.E.; Van de Vijver, B.; Cox, E.J.; Bicudo, D.D.C.; Ector, L. Tursiocola podocnemicola sp. nov., a new epizoic freshwater diatom species from the Rio Negro in the Brazilian Amazon Basin. Diatom Res. 2012, 27, 1–8. [Google Scholar] [CrossRef]
- Abbott, L.L.; Bergey, E.A. Why are there few algae on snail shells? The effects of grazing, nutrients and shell chemistry on the algae on shells of Helisoma trivolvis. Freshwat. Biol. 2007, 52, 2112–2120. [Google Scholar] [CrossRef]
- Wujek, D.E. The first occurrence of the chrysophyte alga Amphirhiza epizootica from North America. Mich. Bot. 2006, 45, 197–200. [Google Scholar]
- Winemiller, K.; Winsborough, B. Occurrence of epizoic communities on the parasitic copepod Lernaea carassii (Lernaeidae). Southwest. Nat. 1990, 35, 206–210. [Google Scholar] [CrossRef]
- Fuelling, L.J.; Adams, J.A.; Badik, K.J.; Bixby, R.J.; Caprette, C.L.; Caprette, H.E.; Hall, M.M. An unusual occurrence of Thorea hispida (Thore) Desvaux chantransia on rusty crayfish in West Central Ohio. Nova Hedwig. 2012, 94, 355–366. [Google Scholar] [CrossRef]
- Bergey, E.A.; Resh, V.H. Interactions between a stream caddisfly and the algae on its case: Factors affecting algal quantity. Freshwat. Biol. 1994, 31, 153–163. [Google Scholar] [CrossRef]
- Wujek, D.E. Epizooic diatoms on the cerci of Ephemeroptera (Caenidae) naiads. Great Lakes Entomol. 2013, 46, 117–120. [Google Scholar]
- Stewart, K.W.; Schlichting, H.E., Jr. Dispersal of algae and protozoa by selected aquatic insects. J. Ecol. 1966, 54, 551–562. [Google Scholar] [CrossRef]
- Revill, D.L.; Stewart, K.W.; Schlichting, H.E. Passive dispersal of viable algae and protozoa by certain craneflies and midges. Ecology 1967, 48, 1023–1027. [Google Scholar] [CrossRef]
- Göthe, E.; Angeler, D.G.; Gottschalk, S.; Löfgren, S.; Sandin, L. The influence of environmental, biotic and spatial factors on diatom metacommunity structure in Swedish headwater streams. PLoS ONE 2013, 8, e72237. [Google Scholar]
- Burgazzi, G.; Bolpagni, R.; Laini, A.; Racchetti, E.; Viaroli, P. Algal biomass and macroinvertebrate dynamics in intermittent braided rivers: New perspectives from instream pools. River Res. Appl. 2020. [Google Scholar] [CrossRef]
- Wisz, M.S.; Pottier, J.; Kissling, W.D.; Pellissier, L.; Lenoir, J.; Damgaard, C.F.; Dormann, C.F.; Forchhammer, M.C.; Grytnes, J.-A.; Guisan, A.; et al. The role of biotic interactions in shaping distributions and realised assemblages of species: Implications for species distribution modelling. Biol. Rev. 2013, 88, 15–30. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).