Wildfires in Grasslands and Shrublands: A Review of Impacts on Vegetation, Soil, Hydrology, and Geomorphology


Abstract
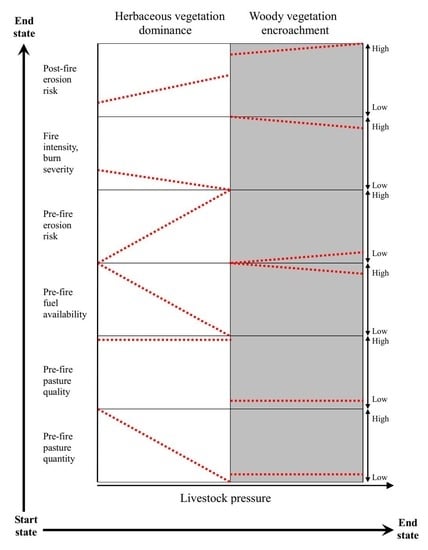
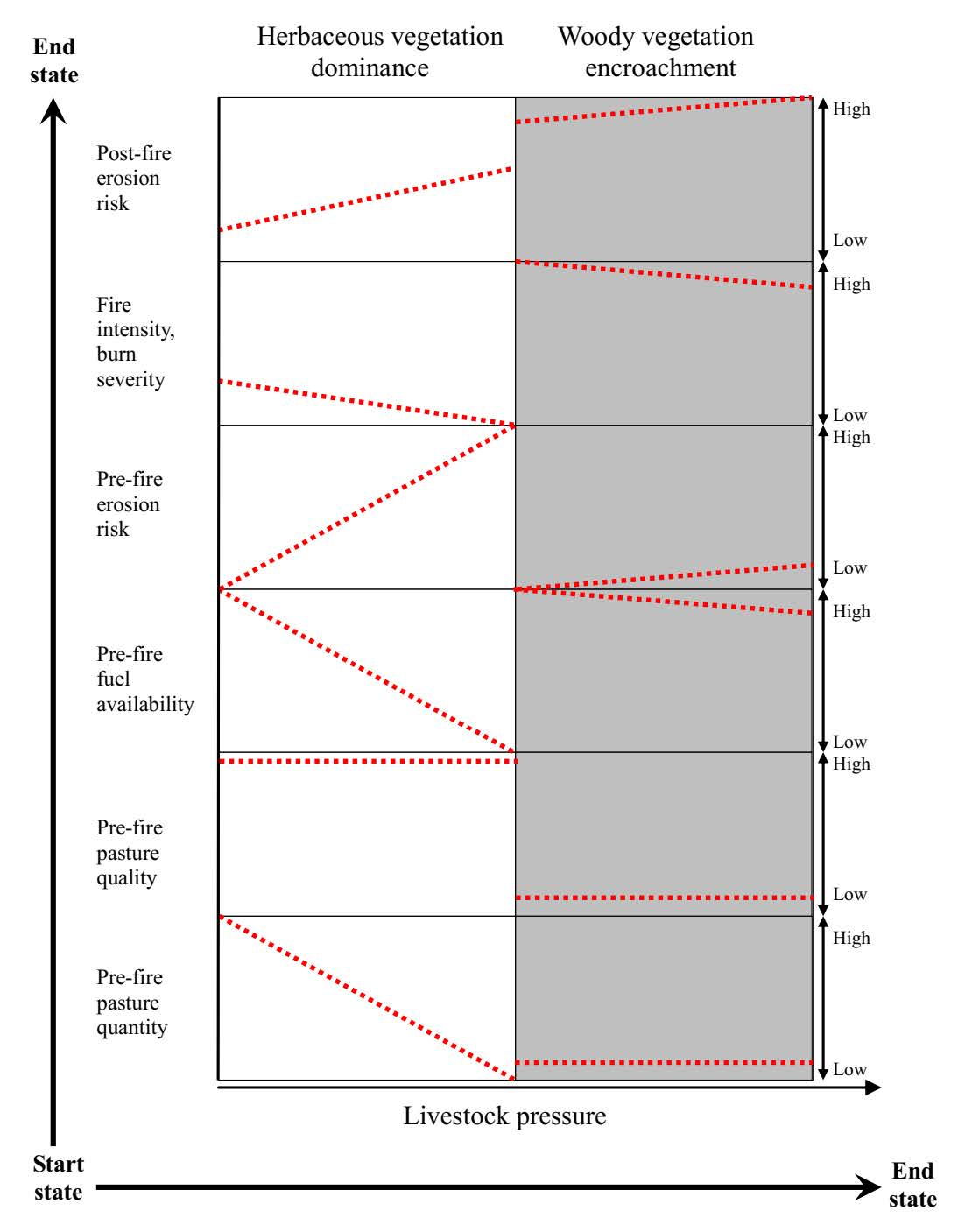{kind=link}
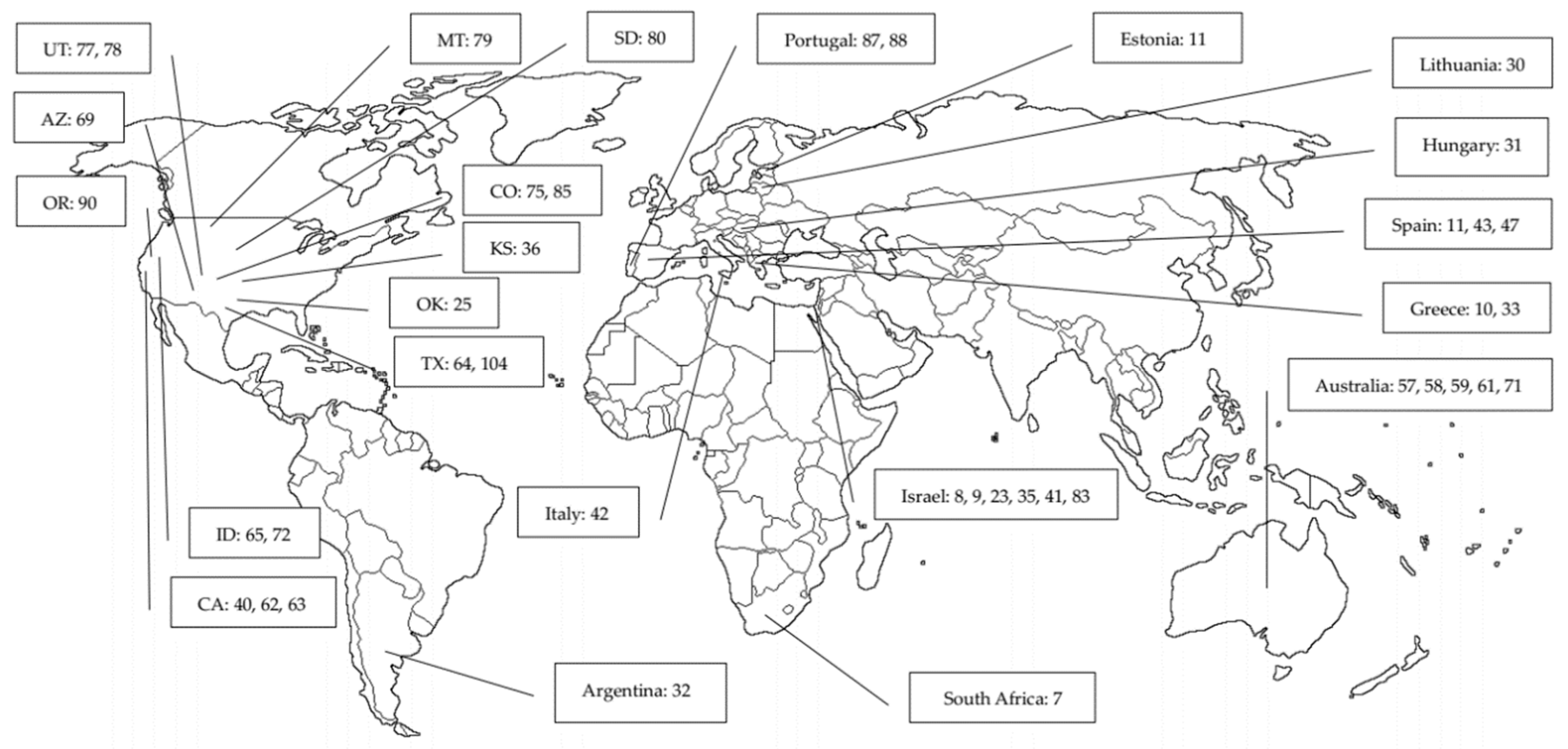{kind=link}
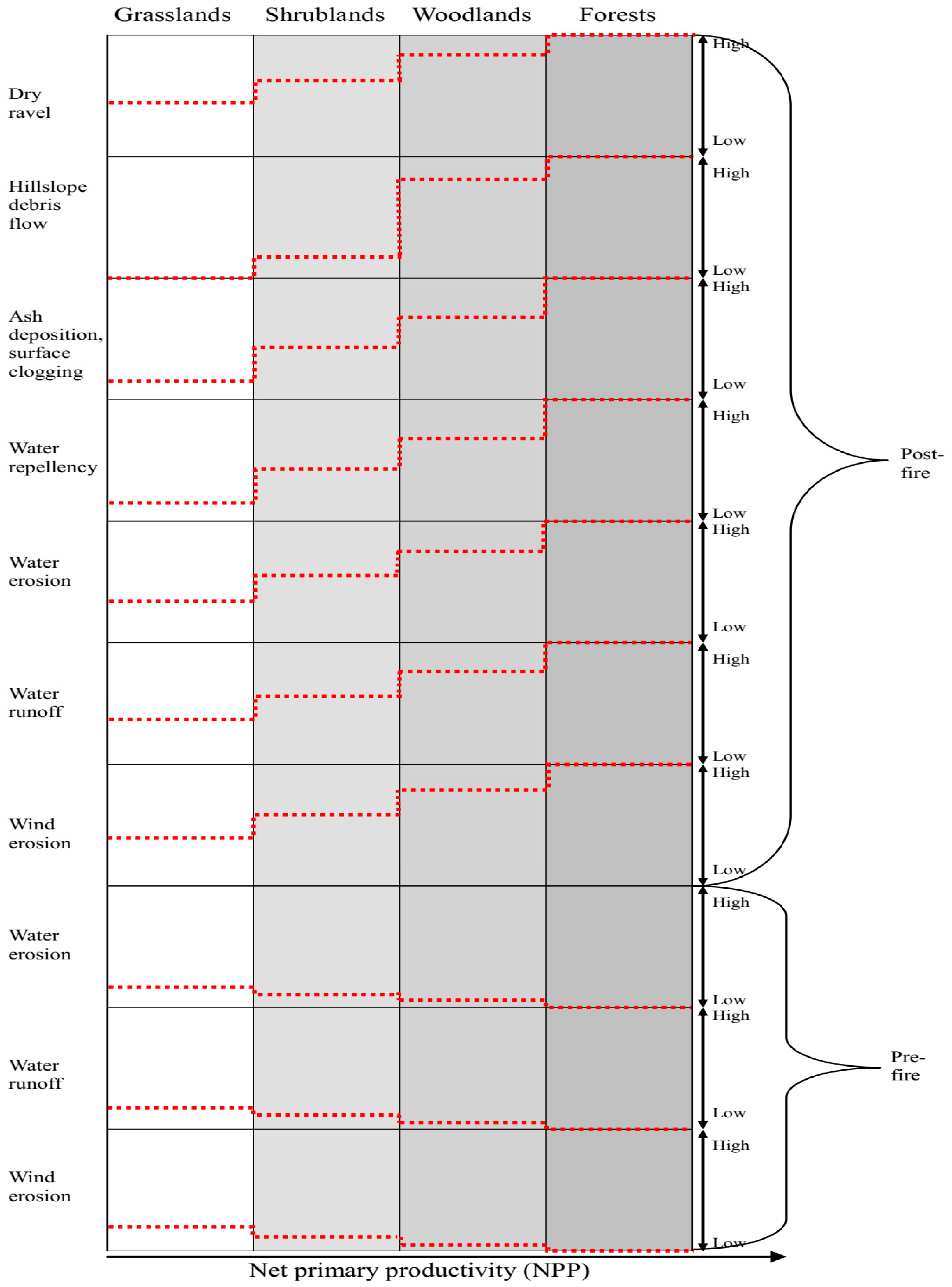{kind=link}
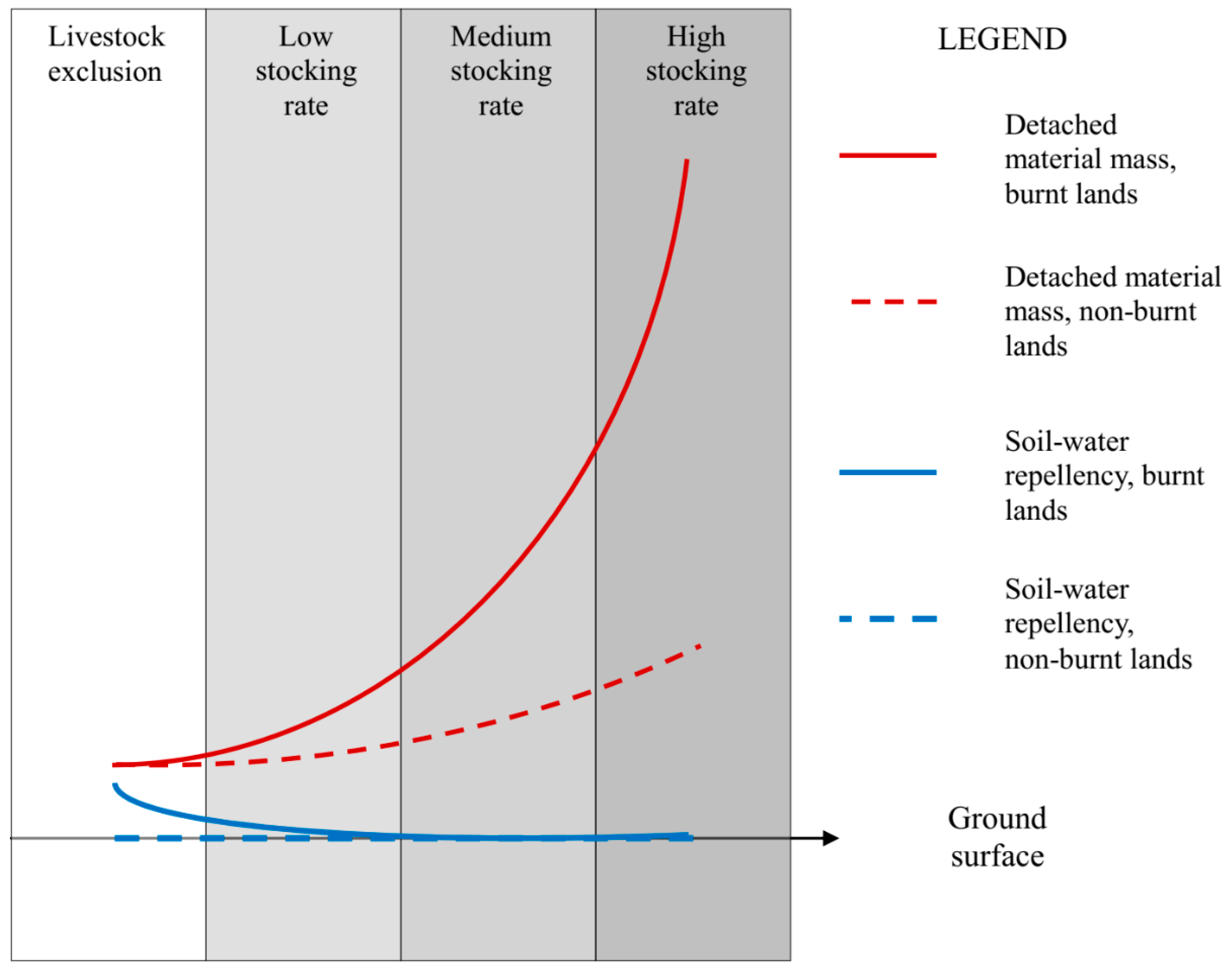{kind=link}
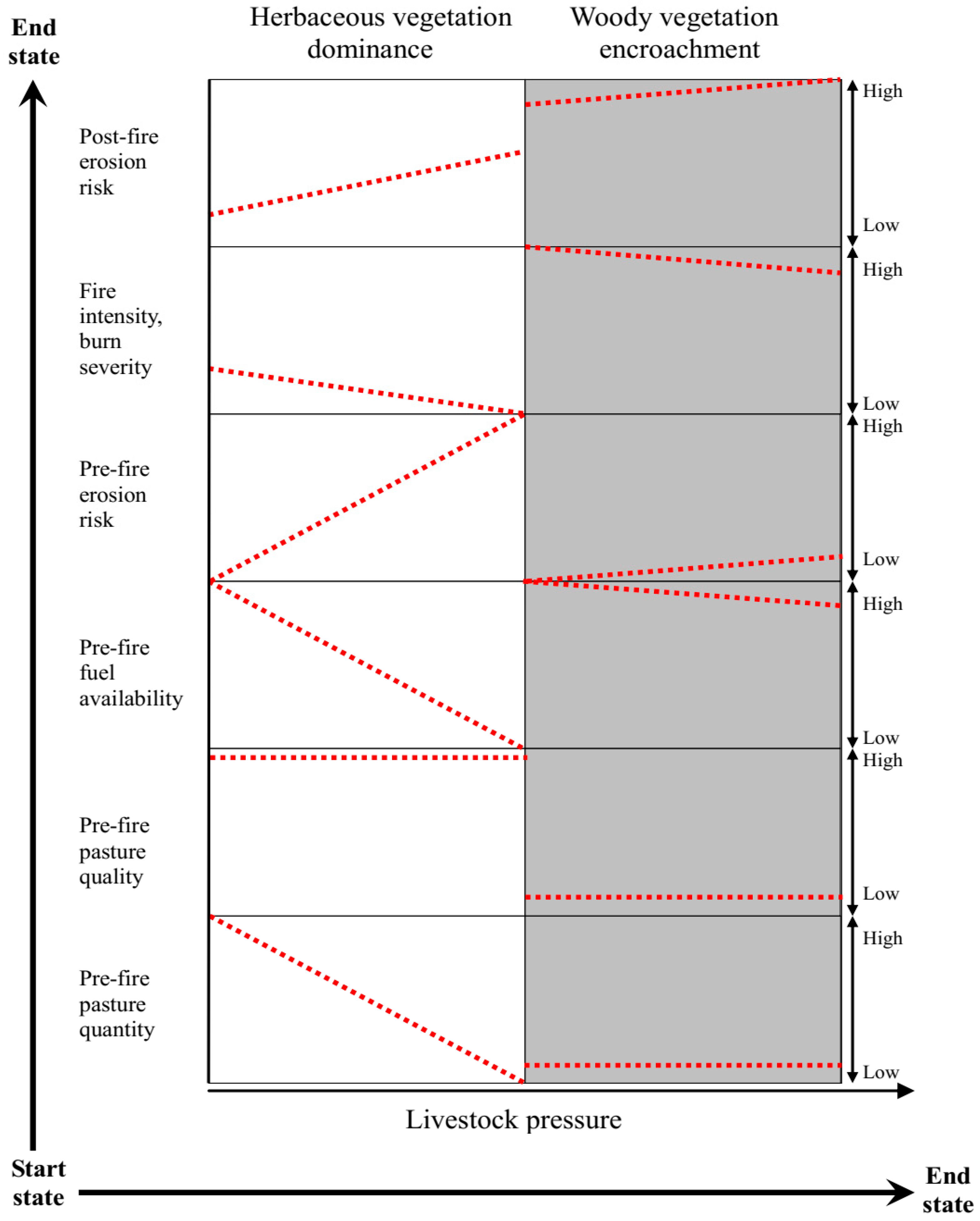{kind=link}
1. Introduction
2. Impacts of Wildfires on Vegetation
3. Impacts of Wildfires on Soil
4. Impacts of Wildfires on Surface Processes
5. Post-fire Ungulate Grazing
6. Environmental Implications, and Recommendations for Land Managers
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hao, R.F.; Yu, D.Y. Optimization schemes for grassland ecosystem services under climate change. Ecol. Indic. 2018, 85, 1158–1169. [Google Scholar] [CrossRef]
- Berg, N.; Perevolotsky, A. National Plan for Monitoring Biological Diversity in the Open Lands of Israel; Israel’s National Nature Assessment Program (HaMaarag): Jerusalem, Israel, 2010; Available online: http://www.hamaarag.org.il/sites/default/files/media/file/report/field_report_report_file/%D7%AA%D7%9B%D7%A0%D7%99%D7%AA%20%D7%9C%D7%90%D7%95%D7%9E%D7%99%D7%AA%20%D7%9C%D7%A0%D7%99%D7%98%D7%95%D7%A8%20%D7%94%D7%9E%D7%92%D7%95%D7%95%D7%9F%20%D7%94%D7%91%D7%99%D7%95%D7%9C%D7%95%D7%92%D7%99.pdf (accessed on 18 May 2019). (In Hebrew)
- Clarck, A.N. Longman Dictionary of Geography—Human and Physical; Butler & Tanner: Essex, UK, 1985; p. 724. [Google Scholar]
- Jongman, R.H.G.; Kamphorst, J.D. The effects of infrastructure on nature. In COST 341 Habitat Fragmentation due to Transport Infrastructure—State of the Arts Report; Europen Co-operation in the Field of Scientific and Technical Research; European Commission: Brussels, Belgium, 2013; Available online: http://www.iene.info/wp-content/uploads/COST341_NationalReport_Netherlands_UKdef.pdf (accessed on 18 May 2019).
- Jolly, W.M.; Cochrane, M.A.; Freeborn, P.H.; Holden, Z.A.; Brown, T.J.; Williamson, G.J.; Bowman, D.M.J.S. Climate-induced variations in global wildfire danger from 1979 to 2013. Nat. Commun. 2013, 6, 7537. [Google Scholar] [CrossRef] [PubMed]
- Carroll, J.M.; Hovick, T.J.; Davis, C.A.; Elmore, R.D.; Fuhlendorf, S.D. Reproductive plasticity and landscape heterogeneity benefit a ground-nesting bird in a fire-prone ecosystem. Ecol. Appl. 2017, 27, 2234–2244. [Google Scholar] [CrossRef]
- Bond, W. Fires, Ecological Effects of. In Encyclopedia of Biodiversity, Volume 2; Academic Press: Cambridge, MA, USA, 2001; pp. 745–753. [Google Scholar]
- KKL. Fire Fighting in Forests, Woodlands, and Open Lands; Keren Kayemet Le’Israel: Jerusalem, Israel, 2013; Available online: http://www.kkl.org.il/files/hebrew_files/michrazim/bid-30-15-reference-document-8.pdf (accessed on 18 May 2019). (In Hebrew)
- Elran, M. Insights from the Wildfires Wave at the National Security Level. Over View 874; The Institute for National Security Studies—Strategic, Innovative, and Policy-Oriented Research; The Institute for National Security Studies: Tel Aviv, Israel, 2016; Available online: https://www.inss.org.il/he/wp-content/uploads/sites/2/systemfiles/874%20M%20E.pdf (accessed on 18 May 2019). (In Hebrew)
- Repousis, S. Pyro terrorism in Greece? Greek forest fires August 2007 and the impact on Greek banks stocks. J. Money Laund. Control 2018, 21, 163–170. [Google Scholar] [CrossRef]
- Deshpande, N. Pyro-terrorism: Recent cases and the potential for proliferation. Stud. Confl. Terror. 2009, 32, 36–44. [Google Scholar] [CrossRef]
- Baird, R.A. Pyro-terrorism–The threat of arson-induced forest fires as a future terrorist weapon of mass destruction. Stud. Confl. Terror. 2006, 29, 415–428. [Google Scholar] [CrossRef]
- Keeley, J.E. Fire intensity, fire severity and burn severity: A brief review and suggested usage. Int. J. Wildland Fire 2009, 18, 116–126. [Google Scholar] [CrossRef]
- Neary, D.G. Impacts of wildfire severity on hydraulic conductivity in forest, woodland, and grassland soils. In Hydraulic Conductivity—Issues, Determination and Applications; Lakshmanan, E., Ed.; InTechOpen: London, UK, 2011; pp. 123–142. [Google Scholar]
- Parsons, A.; Robichaud, P.R.; Lewis, S.A.; Napper, C.; Clark, J.T. Field Guide for Mapping Post-Fire Soil Burn Severity; Gen. Tech. Rep. RMRS-GTR-243; Rocky Mountain Research Station, Forest Service, U.S. Department of Agriculture: Fort Collins, CO, USA, 2010.
- Schmidt, G.A.; Arndt, D. NOAA/NASA Annual Global Analysis for 2016; NASA: Silver Spring, MD, USA, 2017. Available online: https://www.giss.nasa.gov/research/news/20170118/NOAA-NASA_Global_Analysis-2016-FINAL.pdf (accessed on 18 May 2019).
- WMO. WMO Statement on the State of the Global Climate in 2016; WMO No. 1189; World Meteorological Organization: Geneva, Switzerland, 2017; Available online: https://reliefweb.int/sites/reliefweb.int/files/resources/1189_Statement2016_EN.pdf (accessed on 18 May 2019).
- Schlaepfer, D.R.; Bradford, J.B.; Lauenroth, W.K.; Munson, S.M.; Tietjen, B.; Hall, S.A.; Wilson, S.D.; Duniway, M.C.; Jia, G.; Pyke, D.A.; et al. Climate change reduces extent of temperate drylands and intensifies drought in deep soils. Nat. Commun. 2017, 8, 14196. [Google Scholar] [CrossRef]
- Polade, S.D.; Gershunov, A.; Cayan, D.R.; Dettinger, M.D.; Pierce, D.W. Precipitation in a warming world: Assessing projected hydro-climate changes in California and other Mediterranean climate regions. Sci. Rep. 2017, 7, 10783. [Google Scholar] [CrossRef]
- Sommers, W.T.; Loehman, R.A.; Hardy, C.C. Wildland fire emissions, carbon, and climate: Science overview and knowledge needs. For. Ecol. Manag. 2014, 317, 1–8. [Google Scholar] [CrossRef]
- Zavala, L.M.; de Celis, R.; Jordán, A. How wildfires affect soil properties. A brief review. Cuadernos Investig. Geogr. 2014, 40, 311–331. [Google Scholar] [CrossRef]
- Ruckman, E.M.; Schwinning, S.; Lyons, K.G. Effects of phenology at burn time on post-fire recovery in an invasive C4 grass. Restor. Ecol. 2012, 20, 756–763. [Google Scholar] [CrossRef]
- Malkinson, D.; Wittenberg, L. Recurring Wildfires—Geomorphic and Ecological Effects; A Report to the Ministry of Science, Technology, and Space; University of Haifa: Haifa, Israel, 2015. [Google Scholar]
- Wester, D.B.; Rideout-Hanzak, S.; Britton, C.M.; Whitlaw, H. Plant community response to the East Amarillo Complex wildfires in the Southern High Plains, USA. Community Ecol. 2014, 15, 222–234. [Google Scholar] [CrossRef]
- Weir, J.R.; Fuhlendorf, S.D.; Engle, D.M.; Bidwell, T.G.; Cummings, D.C.; Elmore, D.; Limb, R.F.; Allred, B.W.; Scasta, J.D.; Winter, S.L. Patch Burning: Integrating Fire and Grazing to Promote Heterogeneity. Publication No. E-998; Department of Natural Resource Ecology and Management, Oklahoma Cooperative Extension Service, Oklahoma State University: Stillwater, OK, USA, 2013; Available online: http://pods.dasnr.okstate.edu/docushare/dsweb/Get/Document-4677/E-998.pdf (accessed on 18 May 2019).
- Rasmussen, G.A.; Wright, H.A. Germination requirements of flameleaf sumac. J. Range Manag. 1988, 41, 48–52. [Google Scholar] [CrossRef]
- Wright, H.A. Range burning. J. Range Manag. 1974, 27, 5–11. [Google Scholar] [CrossRef]
- Daubenmire, R. Ecology of fire in grasslands. Adv. Ecol. Res. 1968, 5, 209–266. [Google Scholar]
- Ansley, R.J.; Castellano, M.J. Effects of summer fires on woody, succulent, and graminoid vegetation in southern mixedprairie ecosystems: A review. In Proceedings of the 23rd Tall Timbers Fire Ecology Conference: Fire in Grassland and Shrubland Ecosystems; Masters, R.E., Galley, K.E.M., Eds.; Tall Timbers Research Station: Tallahassee, FL, USA, 2007; pp. 63–70. [Google Scholar]
- Pereira, P.; Cerda, A.; Jordan, A.; Bolutiene, V.; Pranskevicius, M.; Úbeda, X.; Mataix-Solera, J. Spatio-temporal vegetation recuperation after a grassland fire in Lithuania. Procedia Environ. Sci. 2013, 19, 856–864. [Google Scholar] [CrossRef]
- Deák, B.; Valkó, O.; Török, P.; Végvári, Z.; Hartel, T.; Schmotzer, A.; Kapocsi, I.; Tóthmérész, B. Grassland fires in Hungary—Experiences of nature conservationists on the effects of fire on biodiversity. Appl. Ecol. Env. Res. 2014, 12, 267–283. [Google Scholar] [CrossRef]
- Guevara, J.C.; Stasi, C.R.; Wuilloud, C.F.; Estevez, O.R. Effects of fire on rangeland vegetation in south-western Mendoza plains (Argentina): Composition, frequency, biomass, productivity and carrying capacity. J. Arid Environ. 1999, 41, 27–35. [Google Scholar] [CrossRef]
- Papanastasis, V.P. Effects of season and frequency of burning on a phryganic rangeland in Greece. J. Range Manag. 1980, 34, 251–255. [Google Scholar] [CrossRef]
- Reeves, M.C.; Manning, M.E.; DiBenedetto, J.P.; Palmquist, K.A.; Lauenroth, W.K.; Bradford, J.B.; Schlaepfer, D.R. Effects of Climate Change on Rangeland Vegetation in the Northern Rockies. In Climate Change and Rocky Mountain Ecosystems; Advances in Global Change Research; Halofsky, J.E., Peterson, D.L., Eds.; Springer: Berlin, Germany, 2016; pp. 97–114. [Google Scholar]
- Perevolotsky, A.; Lahman, A.; Polak, G. Mediterranean Woodlands—Wildfire Impacts; The Society for the Protection of Nature in Israel: Tel Aviv, Israel, 1992; Available online: https://www.teva.org.il/GetFile.asp?CategoryID=11708&ArticleID=23793&ID=10710 (accessed on 18 May 2019).
- Johnson, L.C.; Matchett, J.R. Fire and grazing regulate belowground processes in tallgrass prairie. Ecology 2001, 82, 3377–3389. [Google Scholar] [CrossRef]
- Doerr, S.H.; Shakesby, R.A.; Walsh, R.P.D. Soil water repellency: Its causes, characteristics and hydro-geomorphological significance. Earth-Sci. Rev. 2000, 51, 33–65. [Google Scholar] [CrossRef]
- Letey, J. Causes and consequences of fire-induced soil water repellency. Hydrol. Process. 2001, 15, 2867–2875. [Google Scholar] [CrossRef]
- DeBano, L.F. Water Repellent Soils: A State-of-the-Art; General Technical Report PSW-46; United States Department of Agriculture: Berkeley, CA, USA, 1981.
- Hubbert, K.R.; Wohlgemuth, P.M.; Beyers, J.L.; Narog, M.G.; Gerrard, R. Post-fire soil water repellency, hydrologic response, and sediment yield compared between grass-converted and chaparral watersheds. Fire Ecol. 2012, 8, 143–162. [Google Scholar] [CrossRef]
- Stavi, I.; Barkai, D.; Knoll, Y.M.; Abu Glion, H.; Katra, I.; Brook, A.; Zaady, E. Fire impact on soil-water repellency and functioning of semi-arid croplands and rangelands: Implications for prescribed burnings and wildfires. Geomorphology 2017, 280, 67–75. [Google Scholar] [CrossRef]
- Novara, A.; Gristina, L.; Rühl, J.; Pasta, S.; D’Angelo, G.; La Mantia, T.; Pereira, P. Grassland fire effect on soil organic carbon reservoirs in a semiarid environment. Solid Earth 2013, 4, 381–385. [Google Scholar] [CrossRef]
- Varela, M.E.; Benito, E.; Keizer, J.J. Wildfires effects on soil erodibility of woodlands in NW Spain. Land Degrad. Dev. 2010, 21, 75–82. [Google Scholar] [CrossRef]
- Fultz, L.M.; Moore-Kucera, J.; Dathe, J.; Davinic, M.; Perry, G.; Wester, D.; Schwilk, D.; Rideout-Hanzak, S. Forest wildfire and grassland prescribed fire effects on soil biogeochemical processes and microbial communities: Two case studies in the semiarid Southwest. Appl. Soil Ecol. 2016, 99, 118–128. [Google Scholar] [CrossRef]
- Dooley, S.R.; Treseder, K.K. The effect of fire on microbial biomass: A meta-analysis of field studies. Biogeochemistry 2012, 109, 49–61. [Google Scholar] [CrossRef]
- Docherty, K.M.; Balser, T.C.; Bohannan, B.J.M.; Gutknecht, J.L.M. Soil microbial responses to fire and interacting global change factors in a California annual grassland. Biogeochemistry 2012, 109, 63–83. [Google Scholar] [CrossRef]
- Úbeda, X.; Lorca, M.; Outeiro, L.R.; Bernia, S.; Castellnou, M. Effects of prescribed fire on soil quality in Mediterranean grassland (Prades Mountains, north-east Spain). Int. J. Wildland Fire 2005, 14, 379–384. [Google Scholar] [CrossRef]
- Pereira, P.; Úbeda, X.; Mataix-Solera, J.; Oliva, M.; Novara, A. Short-term changes in soil Munsell colour value, organic matter content and soil water repellency after a spring grassland fire in Lithuania. Solid Earth 2014, 5, 209–225. [Google Scholar] [CrossRef]
- Rundel, P.W. Fire as an ecological factor. In Physiological Plant Ecology I—Responses to the Physical Environment; Lange, O.L., Nobel, P.S., Osmond, C.B., Ziegler, H., Eds.; Springer: Berlin, Germany, 2012; pp. 501–538. [Google Scholar]
- Song, Y.T.; Zhou, D.W.; Zhang, H.X.; Li, G.D.; Jin, Y.H.; Li, Q. Effects of vegetation height and density on soil temperature variations. Chin. Sci. Bull. 2013, 58, 907–912. [Google Scholar] [CrossRef]
- Sharrow, S.H.; Wright, H.A. Effects of fire, ash, and litter on soil Nitrate, temperature, moisture and Tobosagrass production in the Rolling Plains. J. Range Manag. 1977, 4, 197–270. [Google Scholar] [CrossRef]
- Terefe, T.; Mariscal-Sancho, I.; Peregrina, F.; Espejo, R. Influence of heating on various properties of six Mediterranean soils. A laboratory Study. Geoderma 2008, 143, 273–280. [Google Scholar] [CrossRef]
- Lu, J.; Zheng, F.; Li, G.; Bian, F.; An, J. The effects of raindrop impact and runoff detachment on hillslope soil erosion and soil aggregate loss in the Mollisol region of Northeast China. Soil Tillage Res. 2016, 161, 79–85. [Google Scholar] [CrossRef]
- Tongway, D.J.; Ludwig, J.A. The nature of landscape dysfunction in rangelands. In Landscape Ecology Function and Management; Ludwig, J.A., Tongway, D.J., Freudenberger, D., Noble, J., Hodgkinson, K., Eds.; CSIRO Publishing: Canbbera, Australia, 2003; pp. 49–61. [Google Scholar]
- DeBano, L.F. The role of fire and soil heating on water repellency in wildland environments: A review. J. Hydrol. 2000, 231–232, 195–206. [Google Scholar] [CrossRef]
- Johansen, M.P.; Hakonson, T.E.; Breshears, D.D. Post-fire runoff and erosion from rainfall simulation: Contrasting forests with shrublands and grasslands. Hydrol. Process. 2001, 15, 2953–2965. [Google Scholar] [CrossRef]
- Nyman, P.; Smith, H.G.; Sherwin, C.B.; Langhans, C.; Lane, P.N.J.; Sheridan, G.J. Predicting sediment delivery from debris flows after wildfire. Geomorphology 2015, 250, 173–186. [Google Scholar] [CrossRef]
- Langhans, C.; Nyman, P.; Noske, P.J.; Van der Sant, R.E.; Lane, P.N.J.; Sheridan, G.J. Post-fire hillslope debris flows: Evidence of a distinct erosion process. Geomorphology 2017, 295, 55–75. [Google Scholar] [CrossRef]
- Nyman, P.; Sheridan, G.J.; Smith, H.G.; Lane, P.N.J. Evidence of debris flow occurrence after wildfire in upland catchments of south-east Australia. Geomorphology 2011, 125, 383–401. [Google Scholar] [CrossRef]
- Sheridan, G.J.; Nyman, P.; Langhans, C.; Cawson, J.; Noske, P.J.; Oono, A.; Van der Sant, R.; Lane, P.N.J. Is aridity a high-order control on the hydro–geomorphic response of burned landscapes? Int. J. Wildland Fire 2016, 25, 262–267. [Google Scholar] [CrossRef]
- Sheridan, G.J.; Lane, P.N.J.; Noske, P.J. Quantification of hillslope runoff and erosion processes before and after wildfire in a wet Eucalyptus forest. J. Hydrol. 2007, 343, 12–28. [Google Scholar] [CrossRef]
- Gabet, E.J. Sediment transport by dry ravel. J. Geophys. Res.-Solid Earth 2003, 108, 2049. [Google Scholar] [CrossRef]
- Florsheim, J.L.; Chin, A.; O’Hirok, L.S.; Storesund, R. Short-term post-wildfire dry-ravel processes in a chaparral fluvial system. Geomorphology 2016, 252, 32–39. [Google Scholar] [CrossRef]
- Stout, J.E. A field study of wind erosion following a grass fire on the Llano Estacado of North America. J. Arid Environ. 2012, 82, 165–174. [Google Scholar] [CrossRef][Green Version]
- Wagenbrenner, N.S.; Germino, M.J.; Lamb, B.K.; Robichaud, P.R.; Foltz, R.B. Wind erosion from a sagebrush steppe burned by wildfire: Measurements of PM10 and total horizontal sediment flux. Aeolian Res. 2013, 10, 25–36. [Google Scholar] [CrossRef]
- Germino, M.J. Wind Erosion Following Wildfire in Great Basin Ecosystems; Graet Basin Factsheet Series—Information and Tools to Conserve and Restore Great Basin Ecosystems, 2015. Available online: https://www.sagegrouseinitiative.com/wp-content/uploads/2015/10/Wind-Erosion-Following-Wildfire-Grt-Basin-fs-6.pdf (accessed on 18 May 2019).
- Danin, A. Dust trapping by Anabasis Articulata in the Extreme Desert. Israel Vegetation Online, 2012. Available online: http://flora.org.il/books/vegetation-of-israel-and-neighboring-countries/chapter-g/local_veg_g5/ (accessed on 18 May 2019).
- McFadden, L.D. Strongly dust-influenced soils and what they tell us about landscape dynamics in vegetated aridlands of the southwestern United States. In The Web of Geological Sciences: Advances, Impacts, and Interactions; Bickford, M.E., Ed.; Geological Society of America: Boulder, CO, USA, 2013; pp. 501–532. [Google Scholar]
- Paige, G.B.; Stone, J.J.; Guertin, P. Evaluation of Post-Wildfire Runoff and Erosion on Semiarid Ecological Sites; USDA Forest Service Proceedings RMRS-P-36; USDA Forest Service: Washington, DC, UAS, 2005. Available online: https://www.fs.fed.us/rm/pubs/rmrs_p036/rmrs_p036_536_538.pdf (accessed on 18 May 2019).
- Morris, R.H.; Bradstock, R.A.; Dragovich, D.; Henderson, M.K.; Penman, T.D.; Ostendorf, B. Environmental assessment of erosion following prescribed burning in the Mount Lofty Ranges, Australia. Int. J. Wildland Fire 2014, 23, 104–116. [Google Scholar] [CrossRef]
- Smith, H.G.; Sheridan, G.J.; Lane, P.N.J.; Noske, P.J.; Heijnis, H. Changes to sediment sources following wildfire in a forested upland catchment, southeastern Australia. Hydrol. Process. 2011, 25, 2878–2889. [Google Scholar] [CrossRef]
- Clark, P.E.; Nielson, R.M.; Lee, J.; Ko, K.; Johnson, D.E.; Ganskopp, D.C.; Chigbrow, J.; Pierson, F.B.; Hardegree, S.P. Prescribed fire effects on activity and movement of cattle in mesic Sagebrush steppe. Rangel. Ecol. Manag. 2017, 70, 437–447. [Google Scholar] [CrossRef]
- Clark, P.E.; Lee, J.; Ko, K.; Nielson, R.M.; Johnson, D.E.; Ganskopp, D.C.; Pierson, F.B.; Hardegree, S.P. Prescribed fire effects on resource selection by cattle in mesic sagebrush steppe. Part 2: Mid-summer grazing. J. Arid Environ. 2016, 124, 398–412. [Google Scholar] [CrossRef]
- Pearson, S.M.; Turner, M.G.; Wallace, L.L.; Romme, W.H. Winter habitat use by large ungulates following fire in northern Yellowstone National Park. Ecol. Appl. 1995, 5, 744–755. [Google Scholar] [CrossRef]
- Hobbs, N.T.; Spowart, R.A. Effects of prescribed fire on nutrition of mountain sheep and mule deer during winter and spring. J. Wildl. Manag. 1984, 48, 551–560. [Google Scholar] [CrossRef]
- Tracy, B.F.; McNaughton, S.J. Elk grazing and vegetation responses following a late season fire in Yellowstone National Park. Plant Ecol. 1997, 130, 111–119. [Google Scholar] [CrossRef]
- Pickford, G.D. The influence of continued heavy grazing and of promiscuous burning on spring-fall ranges in Utah. Ecology 1932, 13, 159–171. [Google Scholar] [CrossRef]
- West, N.E.; Yorks, T.P. Vegetation responses following wildfire on grazed and ungrazed sagebrush semi-desert. J. Range Manag. 2002, 55, 171–181. [Google Scholar] [CrossRef]
- Vermeire, L.T.; Crowder, J.L.; Wester, D.B. Semiarid rangeland Is resilient to summer fire and postfire grazing utilization. Rangel. Ecol. Manag. 2014, 67, 52–60. [Google Scholar] [CrossRef]
- Gates, E.A.; Vermeire, L.T.; Marlow, C.B.; Waterman, R.C. Reconsidering rest following fire: Northern mixed-grass prairie is resilient to grazing following spring wildfire. Agric. Ecosyst. Environ. 2017, 237, 258–264. [Google Scholar] [CrossRef]
- Miller, R.F.; Chambers, J.C.; Pyke, D.A.; Pierson, F.B.; Williams, C.J. A Review of Fire Effects on Vegetation and Soils in the Great Basin Region: Response and Ecological Site Characteristics; Gen. Tech. Rep. RMRS-GTR-308; Rocky Mountain Research Station, Forest Service, U.S. Department of Agriculture: Fort Collins, CO, USA, 2013; p. 126. Available online: http://sagestep.org/pdfs/rmrs_gtr308.pdf (accessed on 18 May 2019).
- Stritar, M.L.; Schweitzer, J.A.; Hart, S.C.; Bailey, J.K. Introduced ungulate herbivore alters soil processes after fire. Biol. Invasions 2010, 12, 313–324. [Google Scholar] [CrossRef]
- Stavi, I.; Barkai, D.; Knoll, Y.M.; Zaady, E. Livestock grazing impact on soil wettability and erosion risk in post-fire agricultural lands. Sci. Total Environ. 2016, 573, 1203–1208. [Google Scholar] [CrossRef]
- Smith, H.G.; Sheridan, G.J.; Lane, P.N.J.; Nyman, P.; Haydon, S. Wildfire effects on water quality in forest catchments: A review with implications for water supply. J. Hydrol. 2011, 396, 170–192. [Google Scholar] [CrossRef]
- Mast, M.A.; Murphy, S.F.; Clow, D.W.; Penn, C.A.; Sexstone, G.A. Water-quality response to a high-elevation wildfire in the Colorado Front Range. Hydrol. Process. 2016, 30, 1811–1823. [Google Scholar] [CrossRef]
- Son, J.H.; Kim, S.; Carlson, K.H. Effects of wildfire on river water quality and riverbed sediment phosphorus. Water Air Soil Pollut 2015, 226, 26. [Google Scholar] [CrossRef]
- Mansilha, C.; Duarte, C.G.; Melo, A.; Ribeiro, J.; Flores, D.; Marques, J.E. Impact of wildfire on water quality in Caramulo Mountain ridge (Central Portugal). Sustain. Water Resour. Manag. 2019, 5, 319–331. [Google Scholar] [CrossRef]
- Mansilha, C.; Carvalho, A.; Guimarães, P.; Marques, J.E. Water quality concerns due to forest fires: Polycyclic Aromatic Hydrocarbons (PAH) contamination of groundwater from mountain areas. J. Toxicol. Environ. Health Part A 2014, 77, 806–815. [Google Scholar] [CrossRef]
- FAO. Wildfre Prevention in the Mediterranean. A Key Issue to Reduce the Increasing Risks of Mediterranean Wildfres in the Context of Climate Changes—Position Paper. In Proceedings of the 5th International Wildland Fire Conference, Avignon, France, 5–8 April 2011; Available online: http://www.fao.org/forestry/40319-06791969d1427714a896b8faeee2aa501.pdf (accessed on 18 May 2019).
- Davies, K.W.; Boyd, C.S.; Bates, J.D.; Hulet, A. Winter grazing can reduce wildfire size, intensity and behaviour in a shrub-grassland. Int. J. Wildland Fire 2015, 25, 191–199. [Google Scholar] [CrossRef]
- Scasta, J.D.; Weir, J.R.; Stambaugh, M.C. Droughts and wildfires in western U.S. rangelands. Rangelands 2016, 38, 197–203. [Google Scholar] [CrossRef]
- Moritz, M.A.; Parisien, M.A.; Batllori, E.; Krawchuk, M.A.; Van Dorn, J.; Ganz, D.J.; Hayhoe, K. Climate change and disruptions to global fire activity. Ecosphere 2012, 3, 49. [Google Scholar] [CrossRef]
- Mills, A.J.; O’Connor, T.G.; Donaldson, J.S.; Fey, M.V.; Skowno, A.L.; Sigwela, A.M.; Lechmere-Oertel, R.G.; Bosenberg, J.D. Ecosystem carbon storage under different land uses in three semi-arid shrublands and a mesic grassland in South Africa. S. Afr. J. Plant Soil 2005, 22, 183–190. [Google Scholar] [CrossRef]
- Urbanski, S. Wildland fire emissions, carbon, and climate: Emission factors. Forest Ecol. Manag. 2014, 317, 51–60. [Google Scholar] [CrossRef]
- Myhre, G.; Shindell, D.; Bréon, F.M.; Collins, W.; Fuglestvedt, J.; Huang, J.; Koch, D.; Lamarque, J.F.; Lee, D.; Mendoza, B.; et al. Anthropogenic and natural radiative forcing. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Vončina, A.; Ferlan, M.; Eler, K.; Batič, F.; Vodnik, D. Effects of fire on carbon fluxes of a calcareous grassland. Int. J. Wildland Fire 2014, 23, 425–434. [Google Scholar] [CrossRef]
- Singh, A.; Agrawal, M. Acid rain and its ecological consequences. J. Environ. Biol. 2008, 29, 15–24. [Google Scholar]
- Strand, T.; Gullett, B.; Urbanski, S.; O’Neill, S.; Potter, B.; Aurell, J.; Holder, A.; Larkin, N.; Moore, M.; Rorig, M. Grassland and forest understorey biomass emissions from prescribed fires in the southeastern United States—RxCADRE 2012. Int. J. Wildland Fire 2016, 25, 102–113. [Google Scholar] [CrossRef]
- Luvuno, L.; Biggs, R.; Stevens, N.; Esler, K. Woody encroachment as a social-ecological regime shift. Sustainability 2018, 10, 2221. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Reynolds, J.F.; Cunningham, G.L.; Huenneke, L.F.; Jarrell, W.M.; Virginia, R.A.; Whitford, W.G. Biological feedbacks in global desertification. Science 1990, 247, 1043–1048. [Google Scholar] [CrossRef]
- Mata-González, R.; Figueroa-Sandoval, B.; Clemente, F.; Manzano, M. Vegetation changes after livestock grazing excludion and shrub control in the southern Chihuahuan Desert. West. N. Am. Nat. 2007, 67, 63–70. [Google Scholar] [CrossRef]
- Kosmas, C.; Detsis, V.; Karamesouti, M.; Kounalaki, K.; Vassiliou, P.; Salvati, L. Exploring long-term impact of grazing management on land degradation in the socio-ecological system of Asteroussia Mountains, Greece. Land 2015, 4, 541–559. [Google Scholar] [CrossRef]
- Twidwell, D.; Fuhlendorf, S.D.; Taylor, C.A., Jr.; Rogers, W.E. Refining thresholds in coupled fire–vegetation models to improve management of encroaching woody plants in grasslands. J. Appl. Ecol. 2013, 50, 603–613. [Google Scholar] [CrossRef]
- Wester, D.B.; Bryant, F.C.; Tjelmeland, A.D.; Grace, J.L.; Mitchell, S.L.; Edwards, J.T.; Hernández, F.; Lyons, R.K.; Clayton, M.K.; Rideout-Hanzak, S.; et al. Tanglehead in southern Texas: A native grass with an invasive behavior. Rangelands 2018, 37–44. [Google Scholar] [CrossRef]




© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stavi, I. Wildfires in Grasslands and Shrublands: A Review of Impacts on Vegetation, Soil, Hydrology, and Geomorphology. Water 2019, 11, 1042. https://doi.org/10.3390/w11051042
Stavi I. Wildfires in Grasslands and Shrublands: A Review of Impacts on Vegetation, Soil, Hydrology, and Geomorphology. Water. 2019; 11(5):1042. https://doi.org/10.3390/w11051042
Chicago/Turabian StyleStavi, Ilan. 2019. "Wildfires in Grasslands and Shrublands: A Review of Impacts on Vegetation, Soil, Hydrology, and Geomorphology" Water 11, no. 5: 1042. https://doi.org/10.3390/w11051042
APA StyleStavi, I. (2019). Wildfires in Grasslands and Shrublands: A Review of Impacts on Vegetation, Soil, Hydrology, and Geomorphology. Water, 11(5), 1042. https://doi.org/10.3390/w11051042