Oxygen Depletion Affects Kinematics and Shoaling Cohesion of Cyprinid Fish




Abstract
:1. Introduction
2. Materials and Methods
3. Results
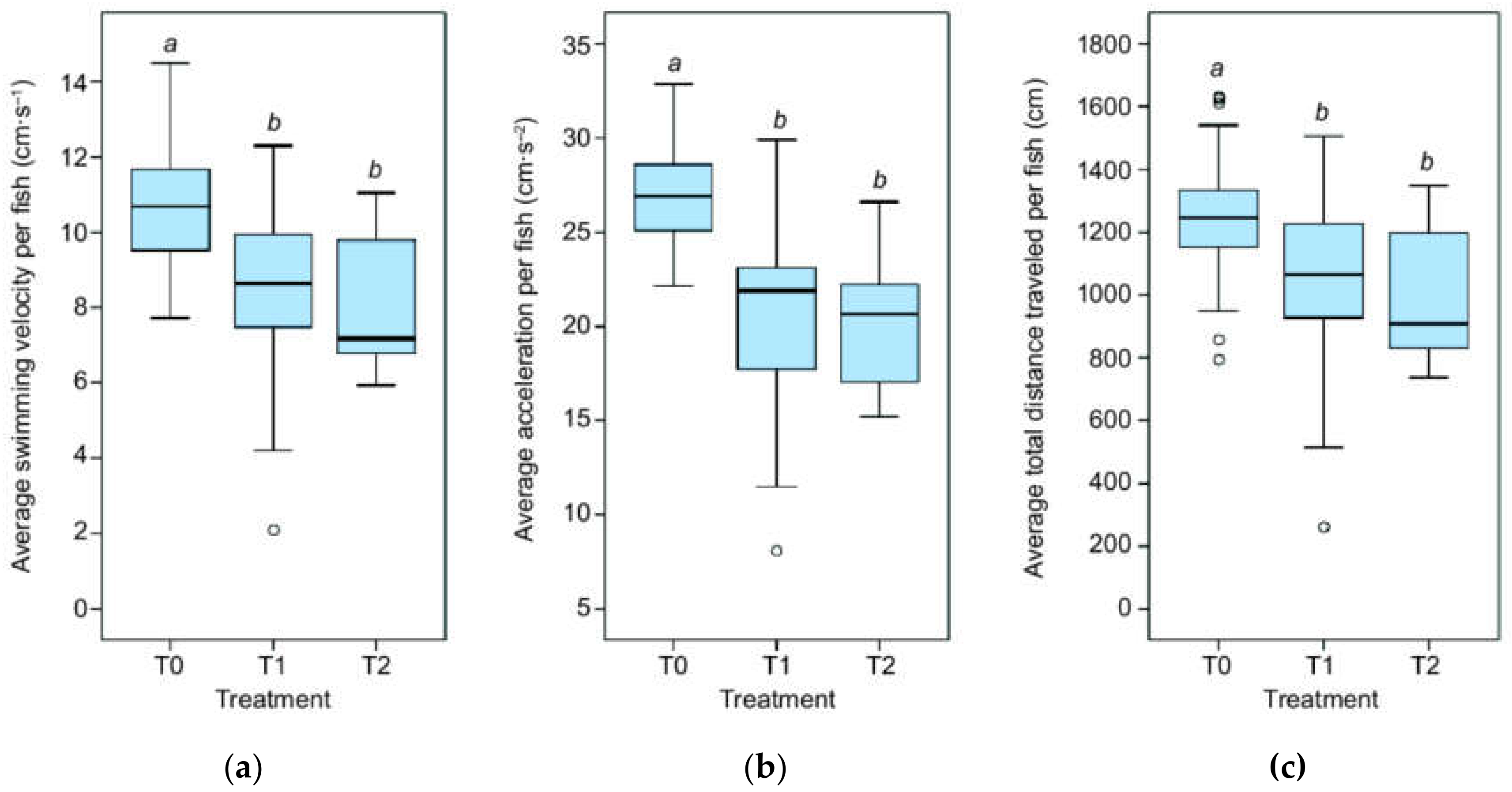
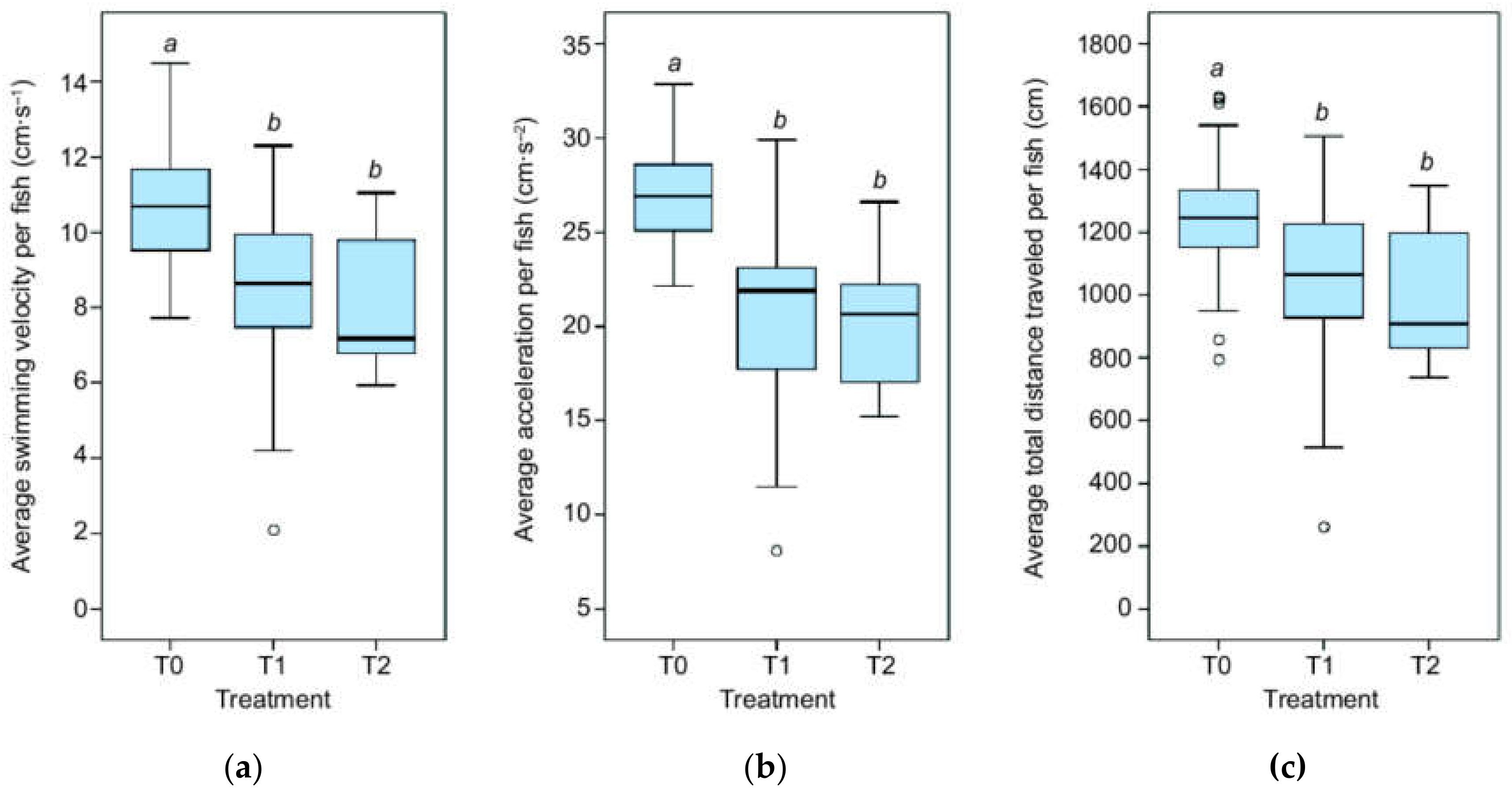
3.1. Kinematics
3.2. Shoaling Cohesion
4. Discussion
4.1. Kinematics
4.2. Shoaling Cohesion
4.3. Ecological Implications
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gasith, A.; Resh, V.H. Streams in Mediterranean climate regions: Abiotic influences and biotic responses to predictable seasonal events. Annu. Rev. Ecol. Syst. 1999, 30, 51–81. [Google Scholar] [CrossRef]
- Lillebø, A.I.; Morais, M.; Guilherme, P.; Fonseca, R.; Serafim, A.; Neves, R. Nutrient dynamics in Mediterranean temporary streams: A case study in Pardiela catchment (Degebe River, Portugal). Limnol. Ecol. Manag. Inl. Waters 2007, 37, 337–348. [Google Scholar] [CrossRef]
- John, S.; Ian, M.; Mike, M.; Ian, S. Effects of short-term oxygen depletion on fish. Environ. Toxicol. Chem. 2000, 19, 2937–2942. [Google Scholar]
- Prat, N.; Munné, A. Water use and quality and stream flow in a Mediterranean stream. Water Res. 2000, 34, 3876–3881. [Google Scholar] [CrossRef]
- Domenici, P.; Herbert, N.A.; Lefrançois, C.; Steffensen, J.F.; McKenzie, D.J. The Effect of Hypoxia on Fish Swimming Performance and Behaviour. In Swimming Physiology of Fish; Palstra, A., Plana, J., Eds.; Springer: Berlin, Heidelberg, 2013; pp. 129–159. [Google Scholar]
- Ekau, W.; Auel, H.; Pörtner, H.O.; Gilbert, D. Impacts of hypoxia on the structure and processes in pelagic communities (zooplankton, macro-invertebrates and fish). Biogeosciences 2010, 7, 1669–1699. [Google Scholar] [CrossRef]
- Herbert, N.A.; Steffensen, J.F. The response of Atlantic cod, Gadus morhua, to progressive hypoxia: Fish swimming speed and physiological stress. Mar. Biol. 2005, 147, 1403–1412. [Google Scholar] [CrossRef]
- Kramer, D.L. Dissolved oxygen and fish behavior. Environ. Biol. Fishes 1987, 18, 81–92. [Google Scholar] [CrossRef]
- Chapman, L.J.; McKenzie, D.J. Behavioral Responses and Ecological Consequences. In Fish Physiology; Richards, J.G., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Burlington, MA, USA, 2009; pp. 25–77. ISBN 978-0-12-374632-0. [Google Scholar]
- Killen, S.S.; Marras, S.; Ryan, M.R.; Domenici, P.; Mckenzie, D.J. A relationship between metabolic rate and risk-taking behaviour is revealed during hypoxia in juvenile European sea bass. Funct. Ecol. 2012, 26, 134–143. [Google Scholar] [CrossRef]
- Domenici, P.; Steffensen, J.F.; Marras, S. The effect of hypoxia on fish schooling. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160236. [Google Scholar] [CrossRef] [PubMed]
- Landman, M.J.; Van Den Heuvel, M.R.; Ling, N. Relative sensitivities of common freshwater fish and invertebrates to acute hypoxia. N. Z. J. Mar. Freshw. Res. 2005, 39, 1061–1067. [Google Scholar] [CrossRef]
- Giorgi, F. Climate change hot-spots. Geophys. Res. Lett. 2006, 33, L08707. [Google Scholar] [CrossRef]
- European Environment Agency (EEA). Impacts of Europe’s Changing Climate–Indicator Based Assessment; EEA: Copenhagen, Denmark, 2008. [Google Scholar]
- Da Cunha, L.V.; Da Oliveira, R.P.; Nascimento, J.; Ribeiro, L. Impacts of climate change on water resources: A case-study for Portugal. In Water in Celtic Countries: Quantity, Quality and Climate Variability, Proceedings of the Fourth InterCeltic Colloquium on Hydrology and Management of Water Resources; IAHS Publ.: Guimarães, Portugal, 2007; pp. 37–48. [Google Scholar]
- Ficke, A.D.; Myrick, C.A.; Hansen, L.J. Potential impacts of global climate change on freshwater fisheries. Rev. Fish Biol. Fish. 2007, 17, 581–613. [Google Scholar] [CrossRef]
- Branco, P.; Santos, J.M.; Katopodis, C.; Pinheiro, A.; Ferreira, M.T. Pool-Type Fishways: Two Different Morpho-Ecological Cyprinid Species Facing Plunging and Streaming Flows. PLoS ONE 2013, 8, e65089. [Google Scholar] [CrossRef] [PubMed]
- Branco, P.; Santos, J.M.; Amaral, S.; Romão, F.; Pinheiro, A.N.; Ferreira, M.T. Potamodromous fish movements under multiple stressors: Connectivity reduction and oxygen depletion. Sci. Total Environ. 2016, 572, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.M.; Silva, A.; Katopodis, C.; Pinheiro, P.; Pinheiro, A.; Bochechas, J.; Ferreira, M.T. Ecohydraulics of pool-type fishways: Getting past the barriers. Ecol. Eng. 2012, 48, 38–50. [Google Scholar] [CrossRef]
- Fu, S.; Nie, L.; Wu, H.; Kuang, L.; Huang, Q. The effect of group size on school structure in juvenile black carp. Acta Ecol. Sin. 2016, 36, 6062–6067. [Google Scholar]
- Schurmann, H.; Steffensen, J.F. Spontaneous swimming activity of Atlantic Cod Gadus Morhua exposed to graded hypoxia at three temperatures. J. Exp. Biol. 1994, 197, 129–142. [Google Scholar] [PubMed]
- Small, K.; Kopf, R.K.; Watts, R.J.; Howitt, J. Hypoxia, Blackwater and Fish Kills: Experimental Lethal Oxygen Thresholds in Juvenile Predatory Lowland River Fishes. PLoS ONE 2014, 9, e94524. [Google Scholar] [CrossRef]
- Lewis, W.M., Jr. Morphological adaptations of cyprinodontoids for inhabiting oxygen deficient waters. Copeia 1970, 2, 319–326. [Google Scholar] [CrossRef]
- Crampton, W.G.R. Effects of anoxia on the distribution, respiratory strategies and electric signal diversity of gymnotiform fishes. J. Fish Biol. 1998, 53, 307–330. [Google Scholar] [CrossRef]
- Calapez, A.R.; Branco, P.; Santos, J.M.; Ferreira, T.; Hein, T.; Brito, A.G.; Feio, M.J. Macroinvertebrate short-term responses to flow variation and oxygen depletion: A mesocosm approach. Sci. Total Environ. 2017, 599–600, 1202–1212. [Google Scholar] [CrossRef]
- Peay, S.; Hiley, P.D.; Collen, P.; Martin, I. Biocide treatment of ponds in Scotland to eradicate signal crayfish. Bull. Fr. Pêche Piscic. 2006, 380–381, 1363–1379. [Google Scholar] [CrossRef]
- Miller, N.; Gerlai, R. Automated tracking of zebrafish shoals and the analysis of shoaling behavior. In Zebrafish Protocols for Neurobehavioral Research; Kalueff, A.V., Stewart, A.M., Eds.; Humana Press: New York, NY, USA, 2012; pp. 217–230. [Google Scholar]
- Delcourt, J.; Poncin, P. Shoals and schools: Back to the heuristic definitions and quantitative references. Rev. Fish Biol. Fish. 2012, 22, 595–619. [Google Scholar] [CrossRef]
- Barceló, D.; Sabater, S. Water quality and assessment under scarcity: Prospects and challenges in Mediterranean watersheds. J. Hydrol. 2010, 383, 1–4. [Google Scholar] [CrossRef]
- Mandic, M.; Regan, M.D. Can variation among hypoxic environments explain why different fish species use different hypoxic survival strategies? J. Exp. Biol. 2018, 221, jeb161349. [Google Scholar] [CrossRef] [PubMed]
- Díaz, R.J.; Rosenberg, R. Introduction to Environmental and Economic Consequences of Hypoxia. Int. J. Water Resour. Dev. 2011, 27, 71–82. [Google Scholar] [CrossRef]
- Behrens, J.W.; Steffensen, J.F. The effect of hypoxia on behavioural and physiological aspects of lesser sandeel, Ammodytes tobianus (Linnaeus, 1785). Mar. Biol. 2007, 150, 1365–1377. [Google Scholar] [CrossRef]
- Fischer, P.; Rademacher, K.; Kils, U. In situ investigations on the respiration and behaviour of the eelpout Zoarces viviparus under short-term hypoxia. Mar. Ecol. Prog. Ser. 1992, 88, 181–184. [Google Scholar] [CrossRef]
- Nilsson, G.E.; Rosen, P.R.; Johansson, D. Anoxic depression of spontaneous locomotor activity in crucian carp quantified by a computerized imaging technique. J. Exp. Biol. 1993, 180, 153–162. [Google Scholar]
- Domenici, P.; Silvana Ferrari, R.; Steffensen, J.F.; Batty, R.S. The effect of progressive hypoxia on school structure and dynamics in Atlantic herring Clupea harengus. Proc. R. Soc. B Biol. Sci. 2002, 269, 2103–2111. [Google Scholar] [CrossRef]
- Schurmann, H.; Steffensen, J.F.; Lomholt, J.P. The influence of hypoxia on the preferred temperature of rainbow trout Oncorhynchus mykiss. J. Exp. Biol. 1991, 157, 75–86. [Google Scholar]

{kind=link}
| Variable | Treatments | |||
|---|---|---|---|---|
| Control | Mild Depletion | High Depletion | ||
| Water quality | O2 (%) | (Normoxia) | 48.4 ± 1.9 | 16.5 ± 1.2 |
| O2 (mg∙L−1) | (Normoxia) | 4.4 ± 0.2 | 1.4 ± 0.1 | |
| Temperature (°C) | 19.4 ± 0.4 | 19.6 ± 0.3 | 22.7 ± 0.1 | |
| Conductivity (μS∙cm−1) | 152 ± 4.5 | 492 ± 16.4 | 738 ± 33.4 | |
| pH | 8.1 ± 0.1 | 8.2 ± 0.1 | 8.4 ± 0.1 | |
| Fish | Total length (mm) | 202 ± 23.9 | 213 ± 29.2 | 191 ± 33.0 |
| Body mass (g) | 81 ± 27.5 | 94 ± 37.5 | 70 ± 37.7 | |
| Video | Analyzed videos | 5 | 5 | 4 1 |
| Analyzed frames | 30,025 | 30,025 | 24,020 1 | |
| Distance (cm ± SE) | |||
|---|---|---|---|
| Treatments | Minimum | Mean | Maximum |
| T0—Control (normoxia) | 14.4 ± 2.7 a | 48.3 ± 8.5 a | 82.2 ± 16.0 a |
| T1—Mild depletion (48.4% DO) | 8.5 ± 0.9 a | 46.6 ± 6.7 a | 88.4 ± 13.0 a |
| T2—High depletion (16.5% DO) | 10.5 ± 0.4 a | 67.8 ± 15.4 a | 124.5 ± 26.0 b |
| Kruskal-Wallis test | χ2 = 1.16, p = 0.56 | χ2 = 4.77, p = 0.09 | χ2 = 8.89, p < 0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayes, D.S.; Branco, P.; Santos, J.M.; Ferreira, T. Oxygen Depletion Affects Kinematics and Shoaling Cohesion of Cyprinid Fish. Water 2019, 11, 642. https://doi.org/10.3390/w11040642
Hayes DS, Branco P, Santos JM, Ferreira T. Oxygen Depletion Affects Kinematics and Shoaling Cohesion of Cyprinid Fish. Water. 2019; 11(4):642. https://doi.org/10.3390/w11040642
Chicago/Turabian StyleHayes, Daniel S., Paulo Branco, José Maria Santos, and Teresa Ferreira. 2019. "Oxygen Depletion Affects Kinematics and Shoaling Cohesion of Cyprinid Fish" Water 11, no. 4: 642. https://doi.org/10.3390/w11040642
APA StyleHayes, D. S., Branco, P., Santos, J. M., & Ferreira, T. (2019). Oxygen Depletion Affects Kinematics and Shoaling Cohesion of Cyprinid Fish. Water, 11(4), 642. https://doi.org/10.3390/w11040642