Crucian Carp (Carassius carassius) Strongly Affect C/N/P Stoichiometry of Suspended Particulate Matter in Shallow Warm Water Eutrophic Lakes


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sampling and Analytical Methods
3. Results
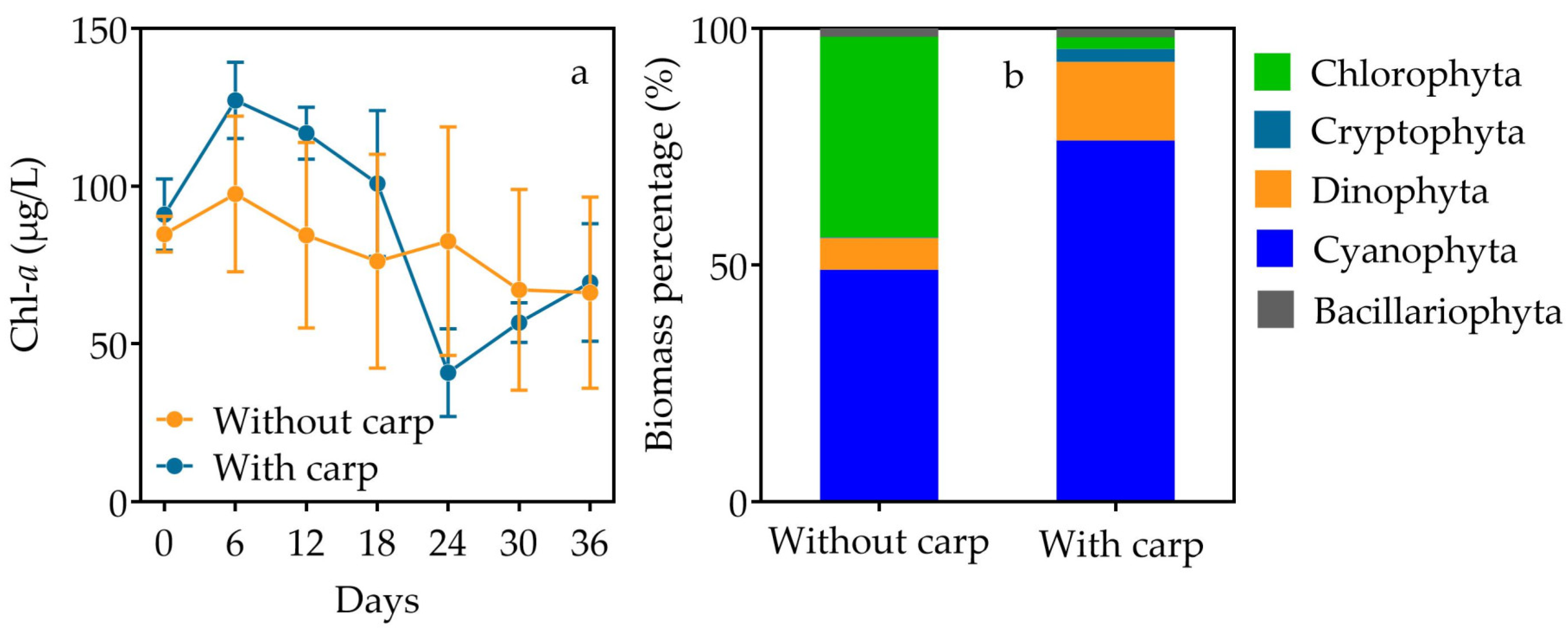
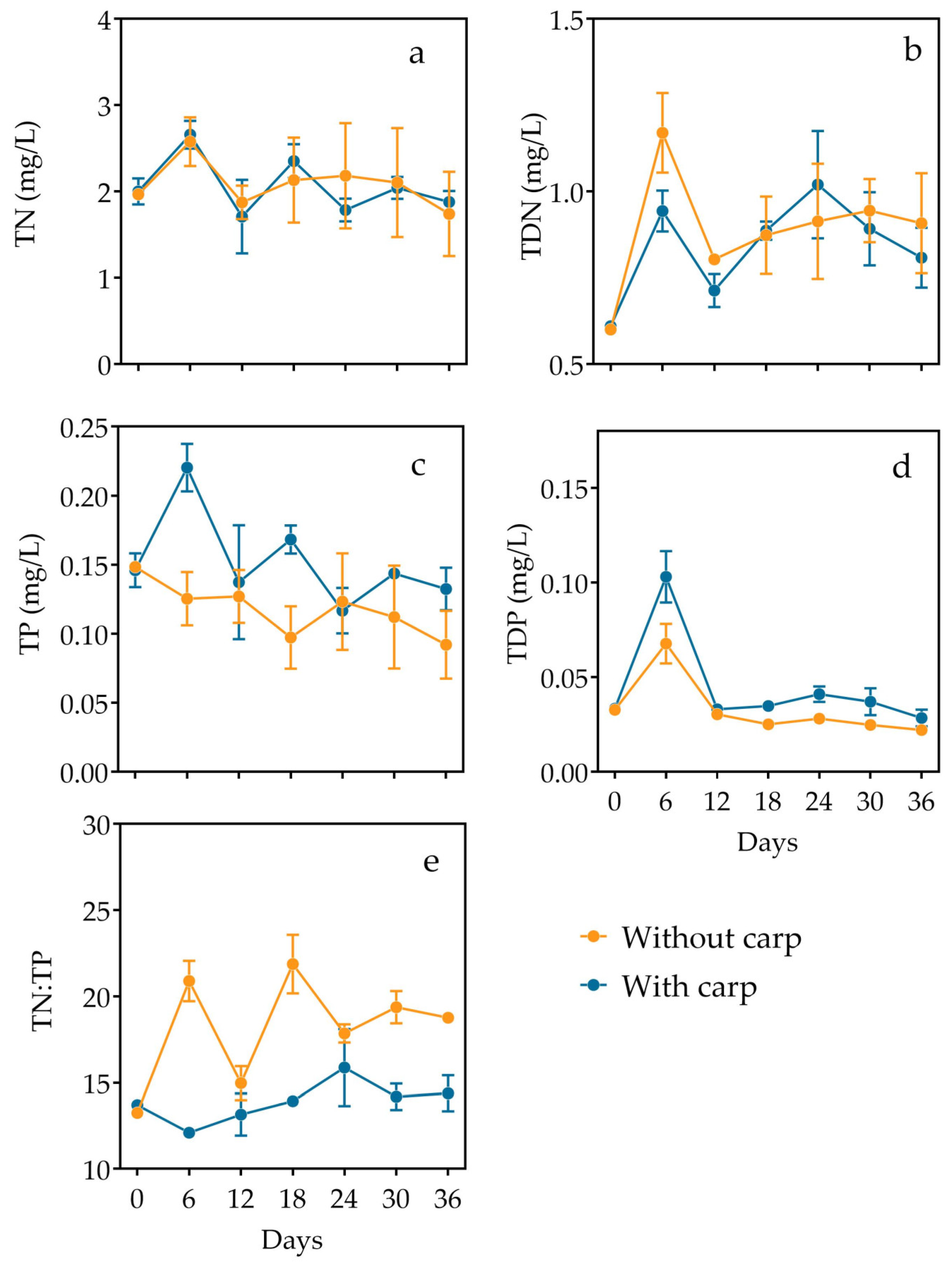
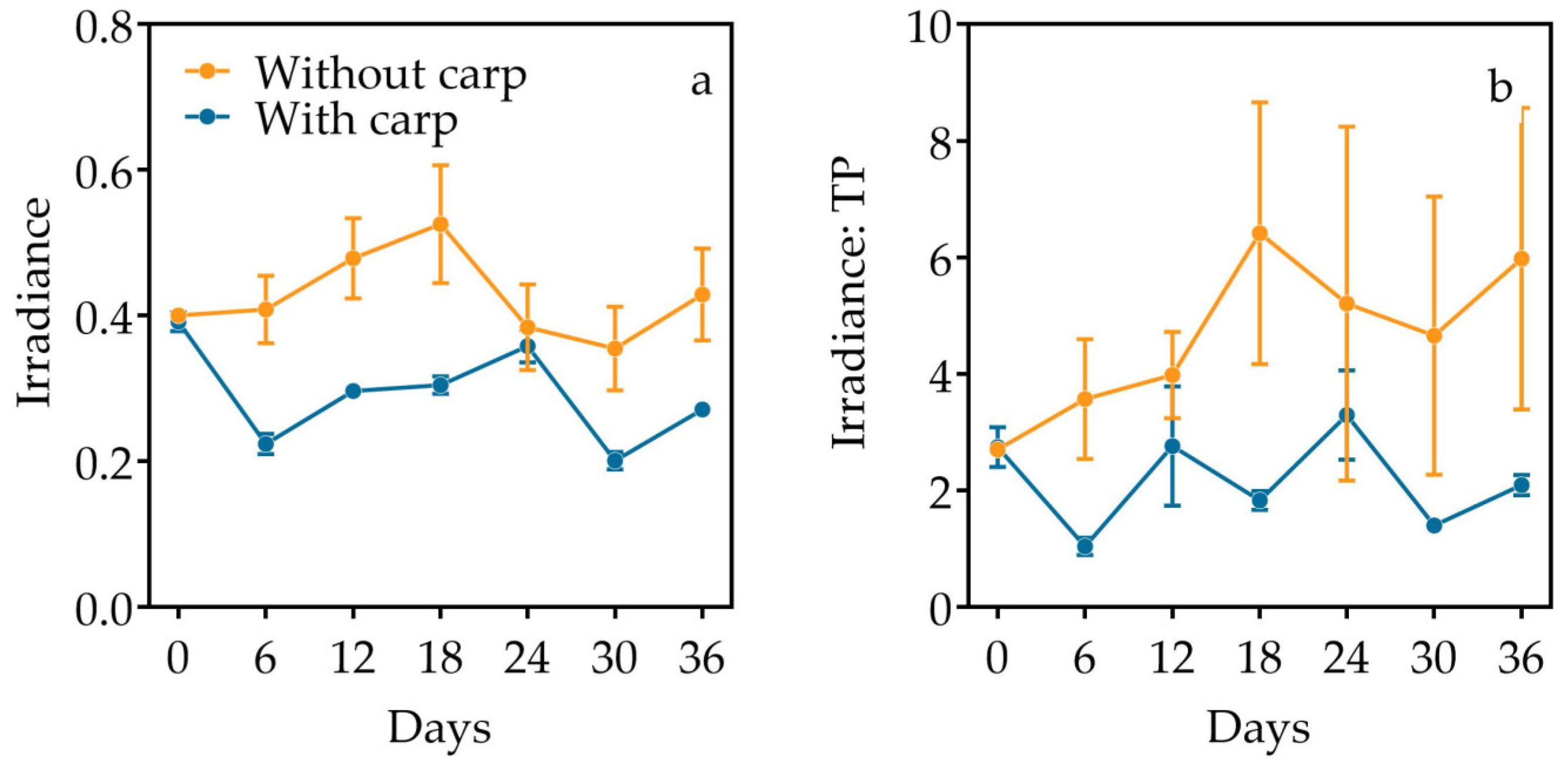
3.1. Nutrients and Light
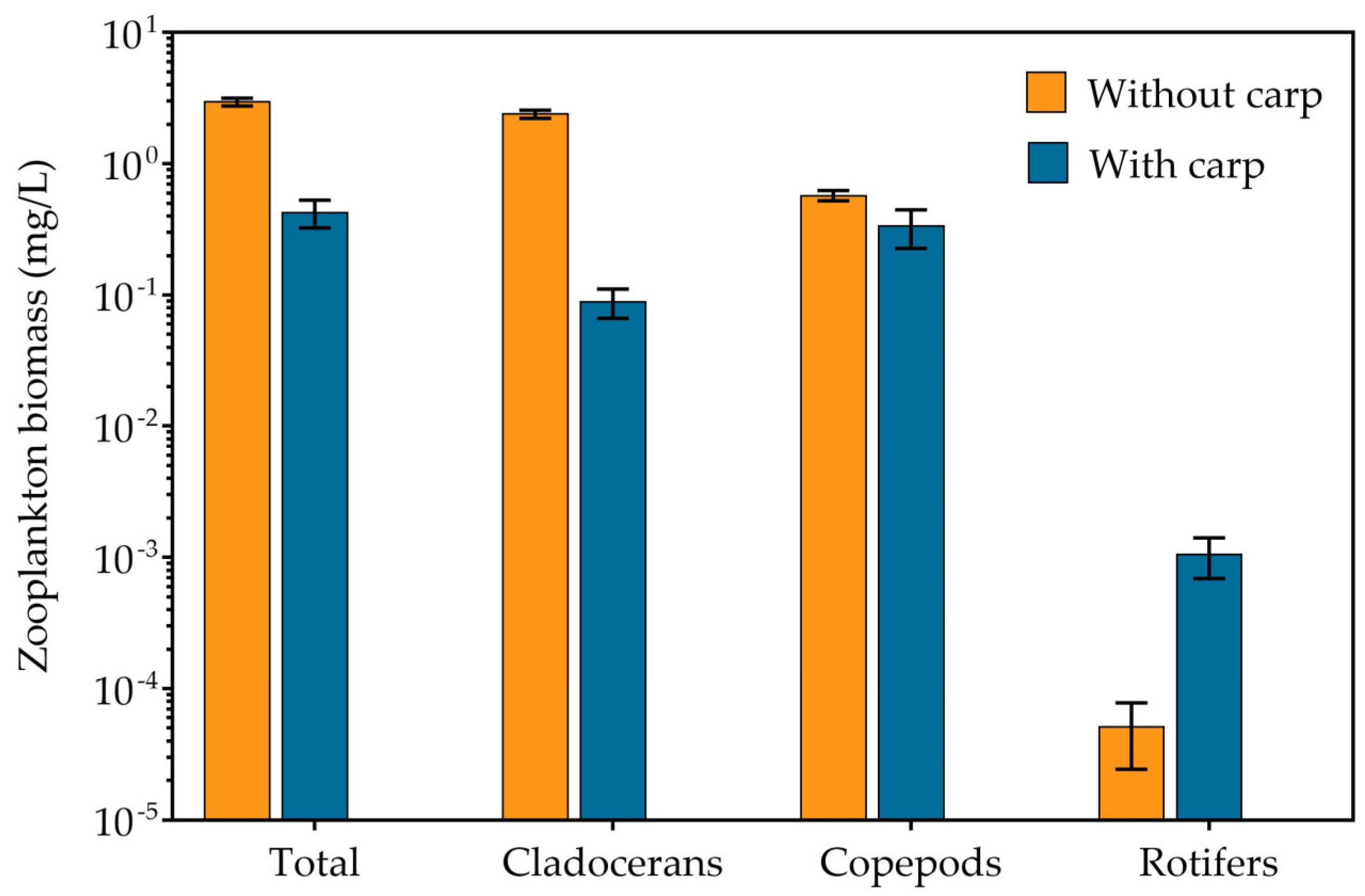
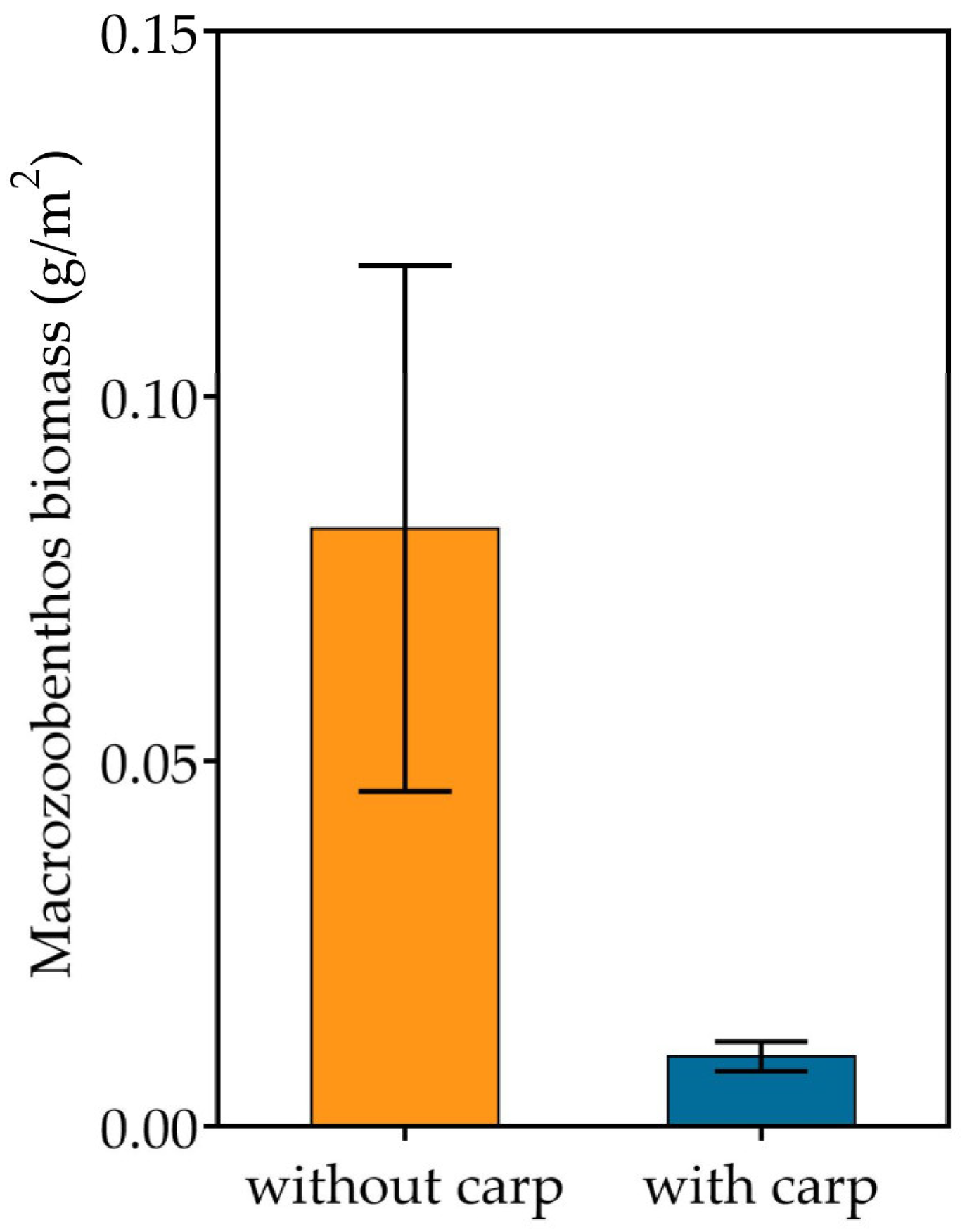
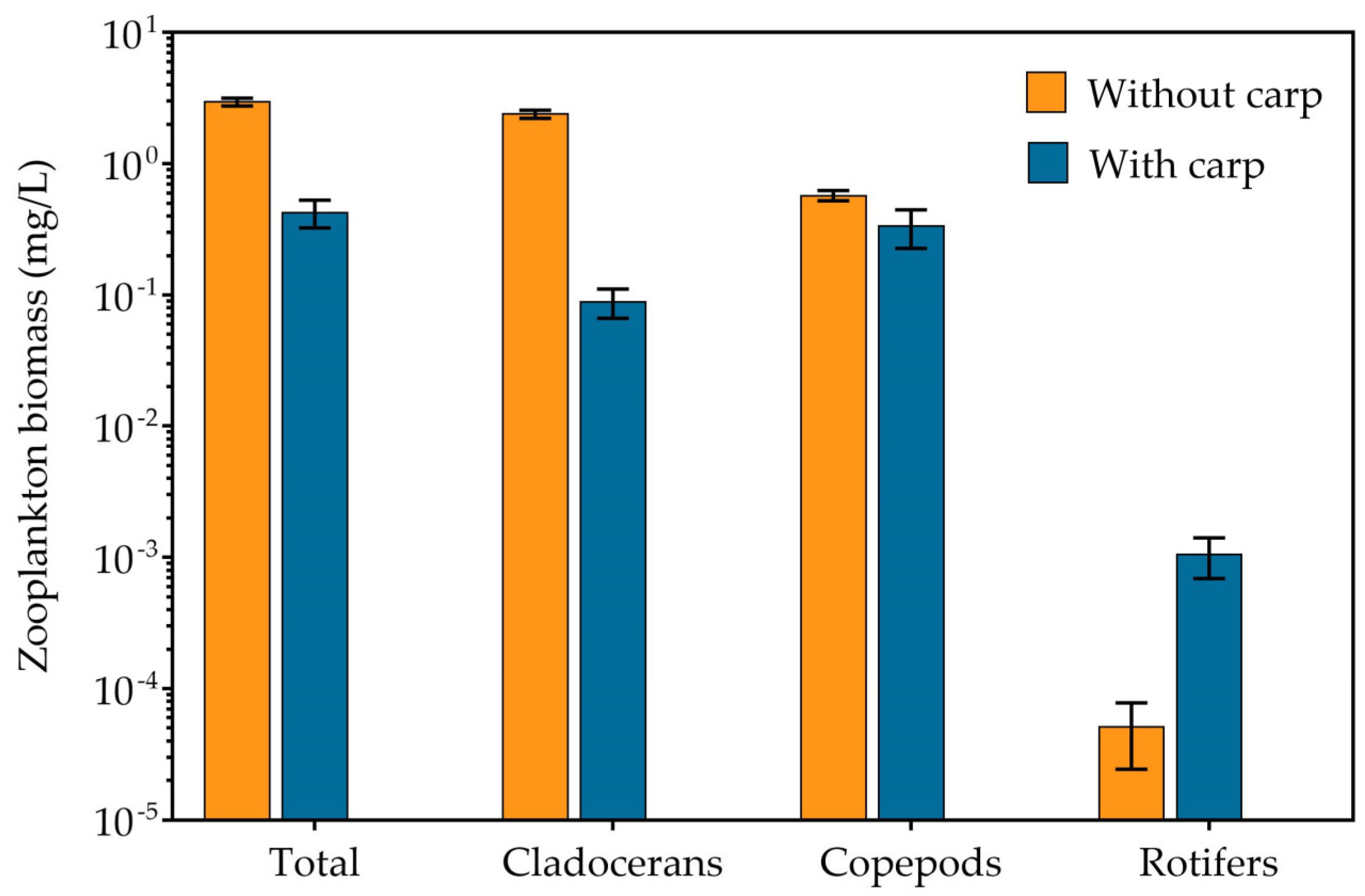
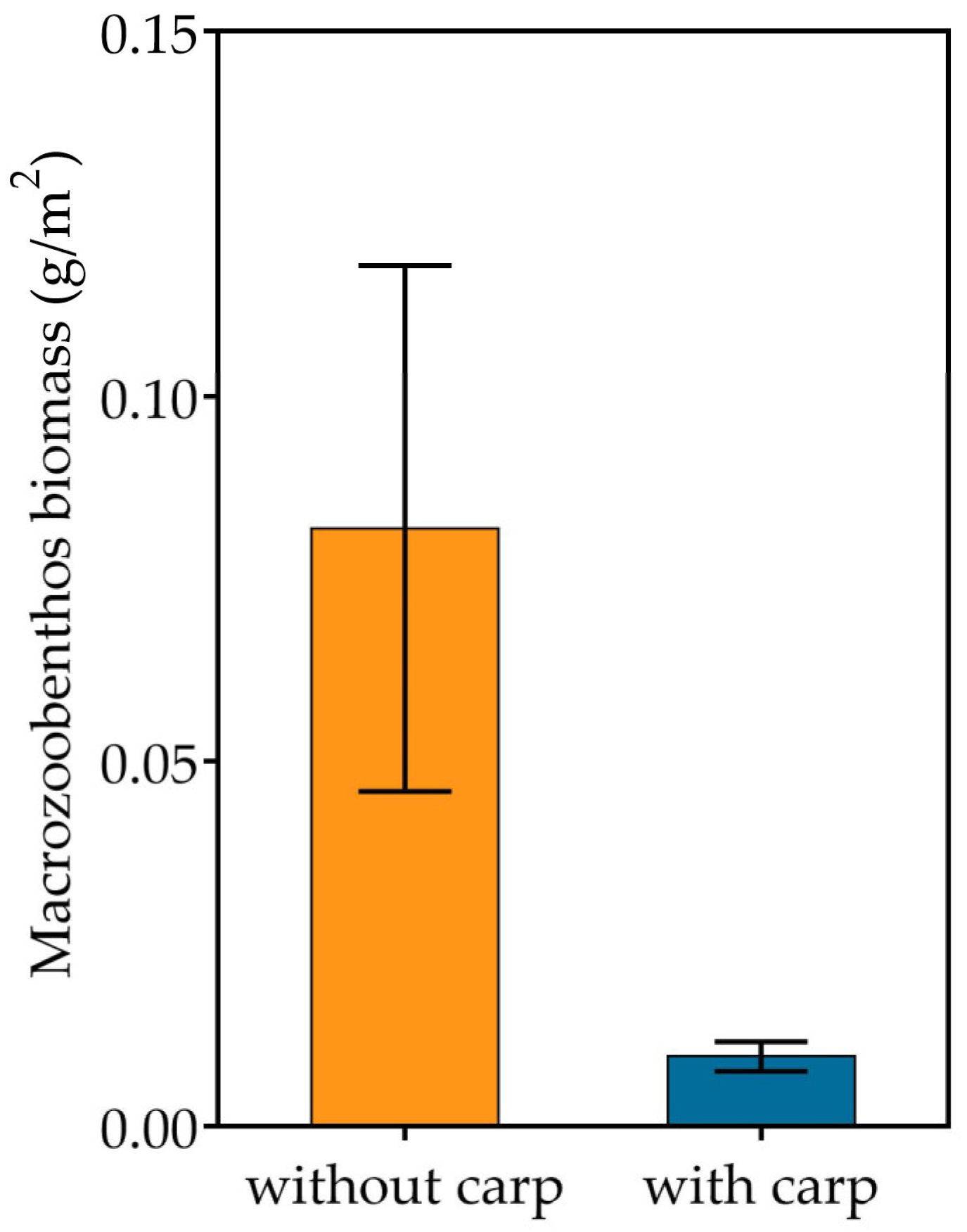
3.2. Phytoplankton, Zooplankton and Zoobenthos
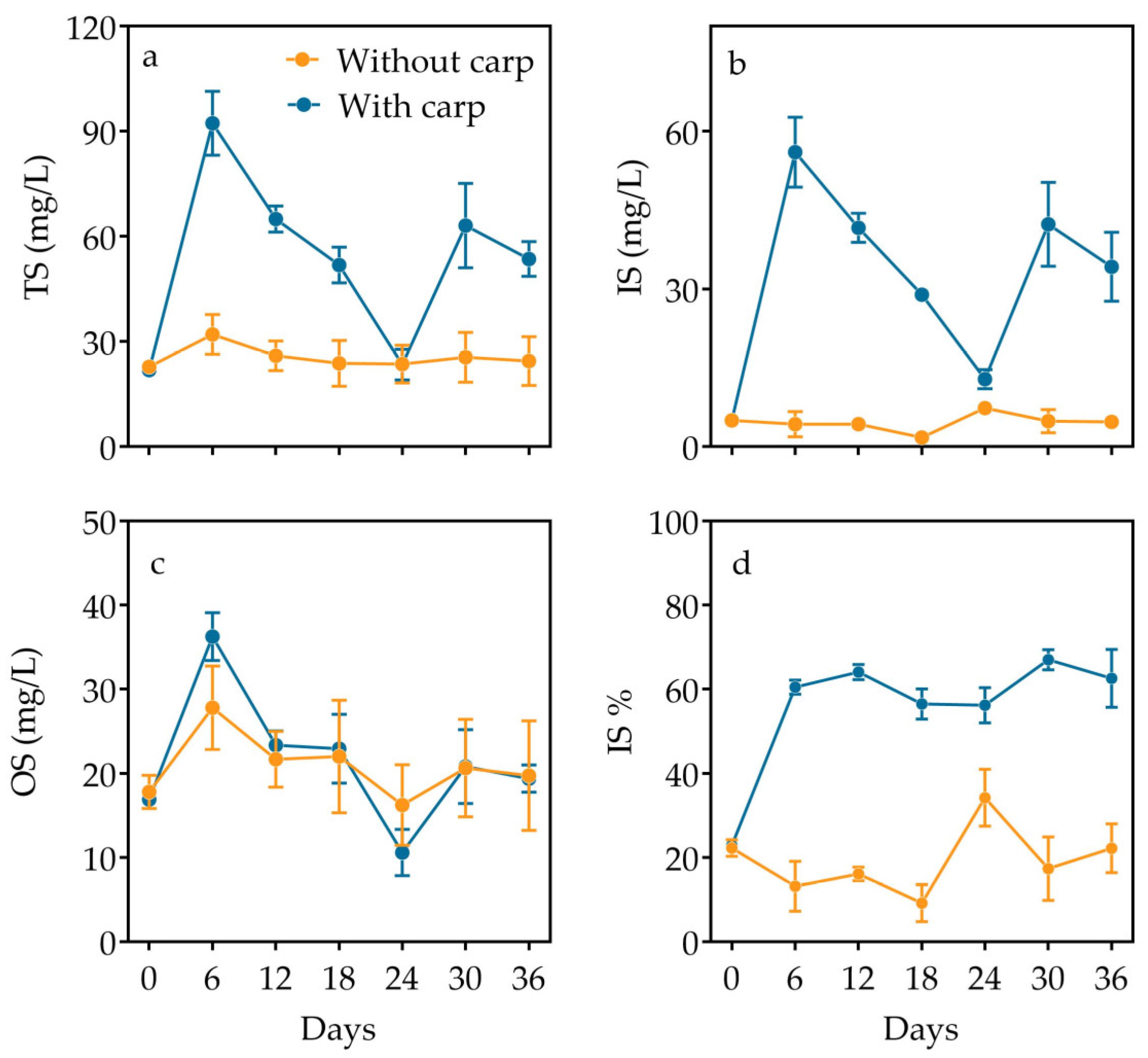
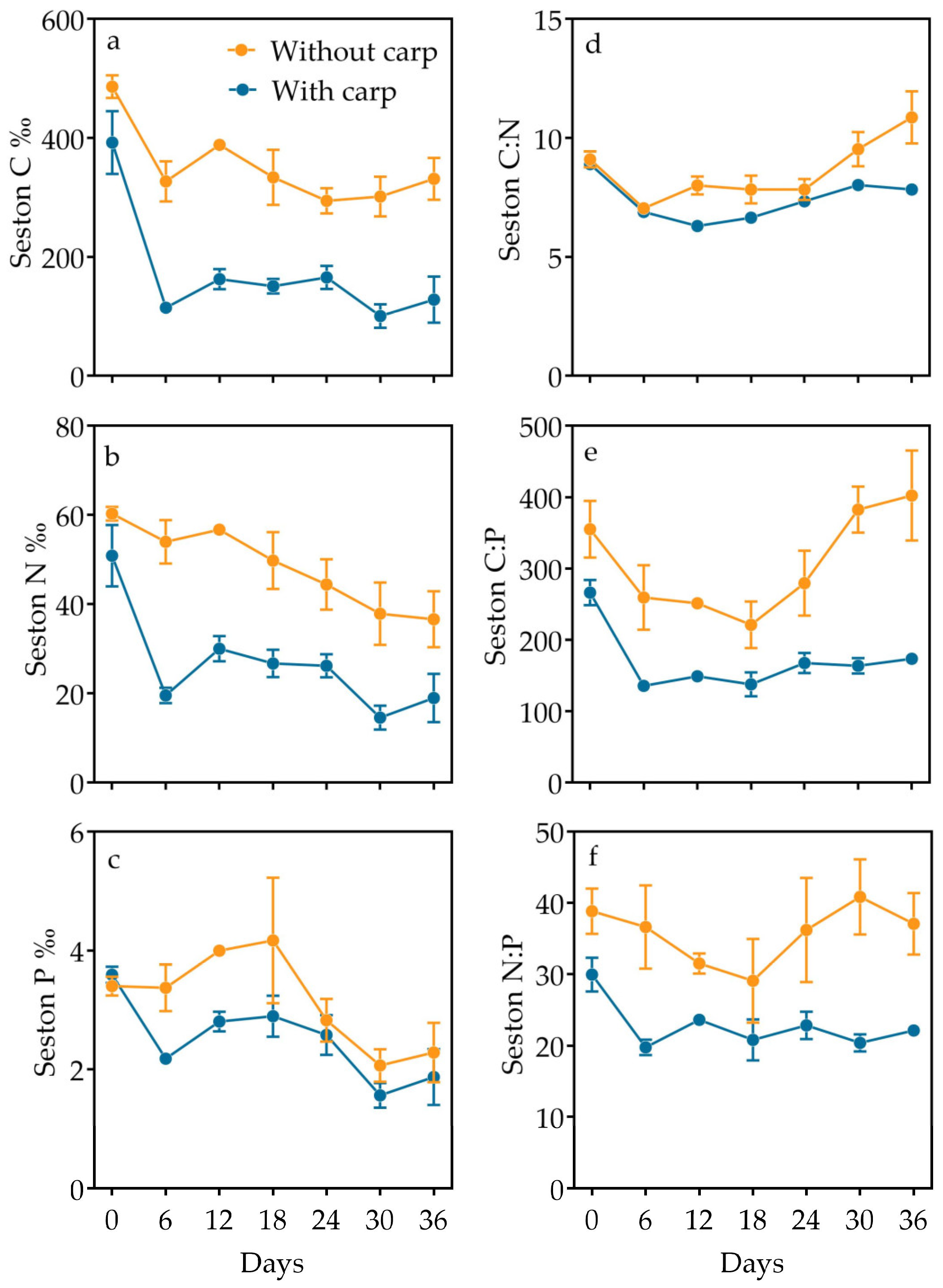
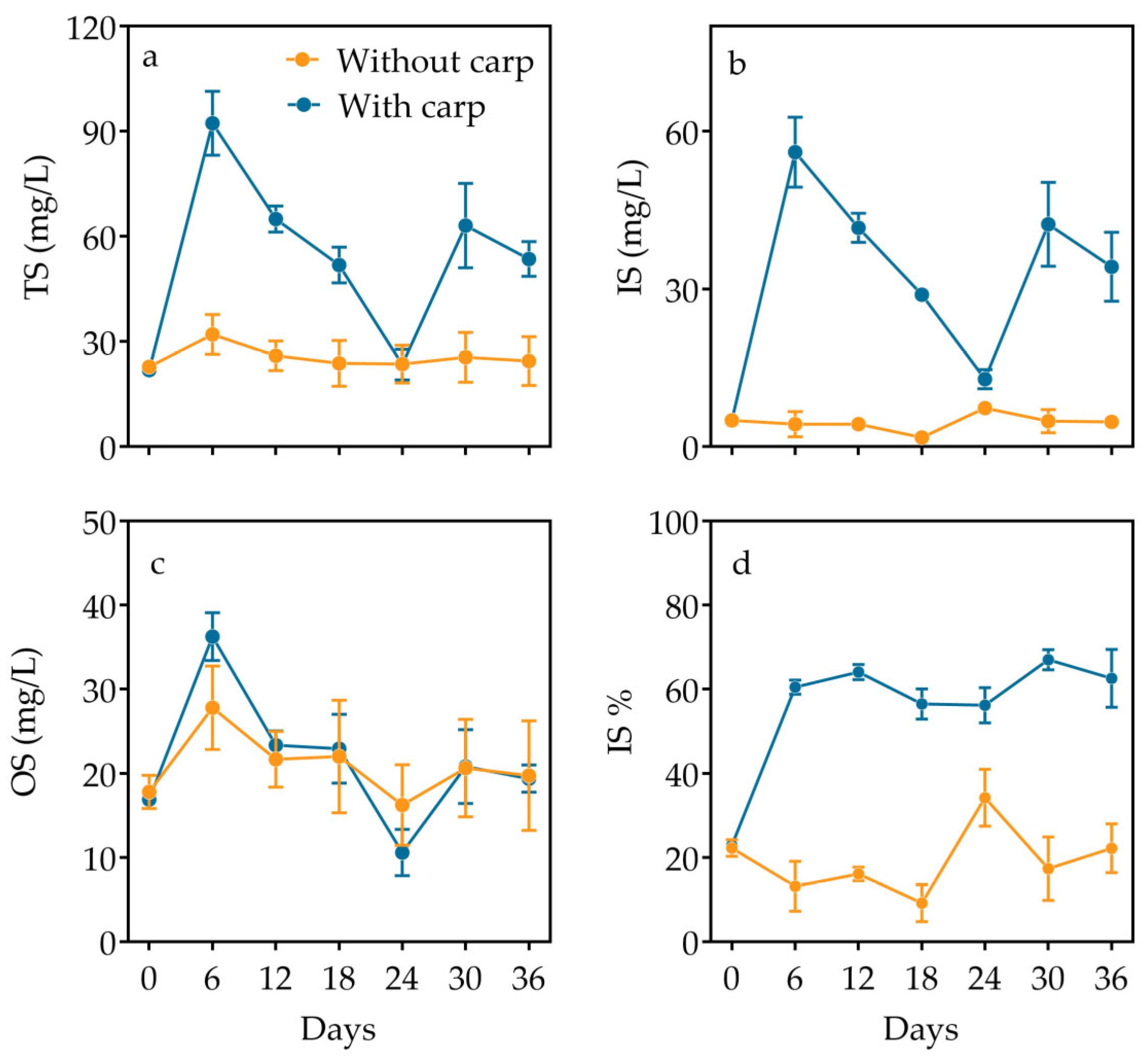
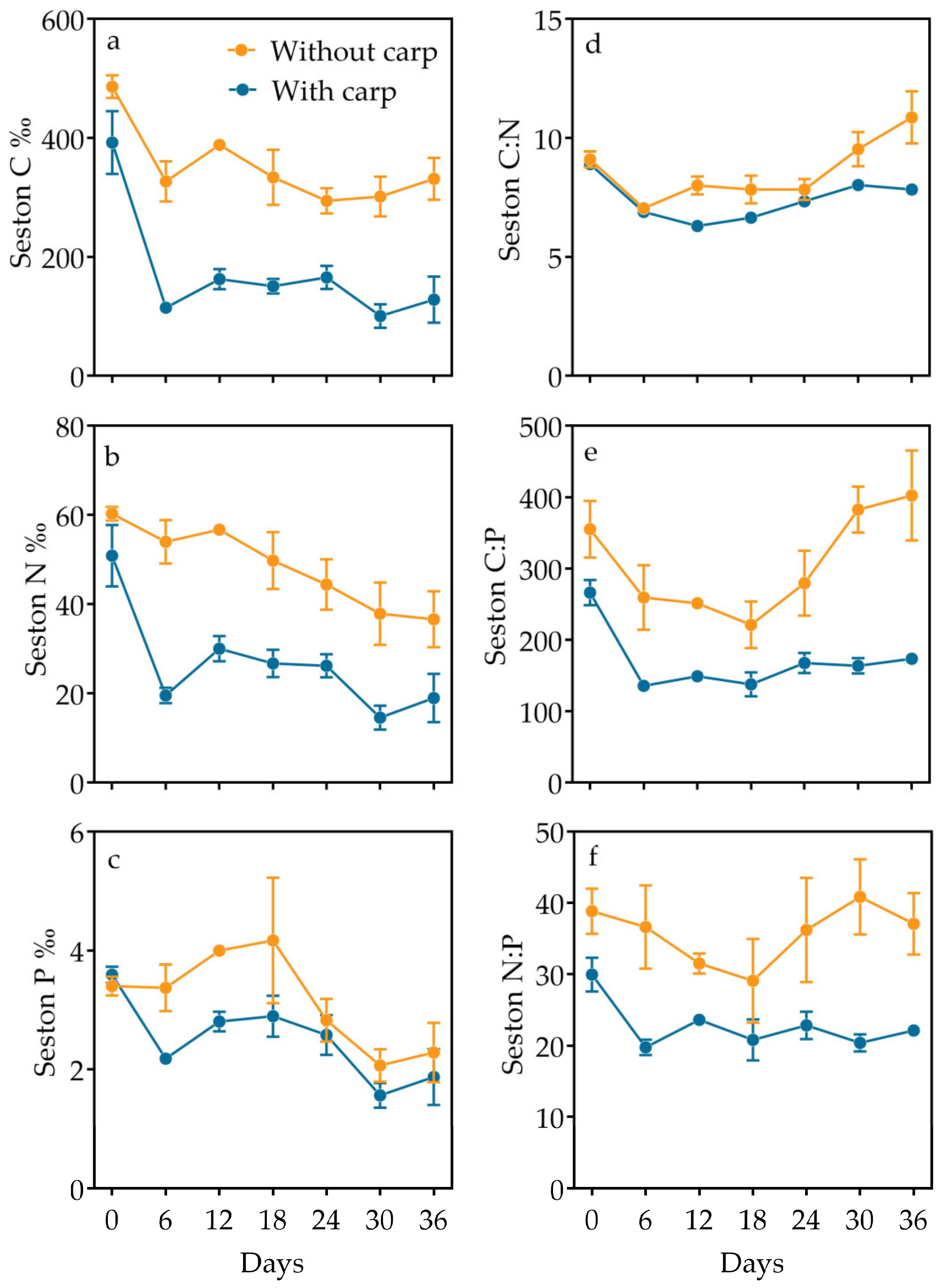
3.3. Inorganic and Organic Partition and Elemental Composition in Particulate Matter
4. Discussion
Potential Implications for Trophic Interactions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hecky, R.E.; Campbell, P.; Hendzel, L.L. The Stoichiometry of carbon, nitrogen, and phosphorus in particulate matter of lakes and oceans. Limnol. Oceanogr. 1993, 38, 709–724. [Google Scholar] [CrossRef]
- Sterner, R.W.; Andersen, T.; Elser, J.J.; Hessen, D.O.; Hood, J.M.; McCauley, E.; Urabe, J. Scale-dependent carbon:nitrogen: Phosphorus seston stoichimetry in marine and freshwaters. Limnol. Oceanogr. 2008, 53, 1169–1180. [Google Scholar] [CrossRef]
- Elser, J.J.; Kyle, M.; Steuer, L.; Nydick, K.R.; Baron, J.S. Nutrient availability and phytoplankton nutrient limitation across a gradient of atmospheric nitrogen deposition. Ecology 2009, 90, 3062–3073. [Google Scholar] [CrossRef] [PubMed]
- Dickman, E.M.; Newell, J.M.; González, M.J.; Vanni, M.J. Light, nutrients, and food-chain length constrain planktonic energy transfer efficiency across multiple trophic levels. Proc. Natl. Acad. Sci. USA 2008, 105, 18408–18412. [Google Scholar] [CrossRef] [PubMed]
- Sterner, R.W.; Elser, J.J.; Fee, E.J.; Guildford, S.J.; Chrzanowski, T.H. The light: Nutrient ratio in lakes: The balance of energy and materials affects ecosystem structure and process. Am. Nat. 1997, 150, 663–684. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Sterner, R.W.; Galford, A.E.; Chrzanowski, T.H.; Findlay, D.L.; Mills, K.H.; Paterson, M.J.; Stainton, M.J.; Schindler, D.W. Pelagic C:N:P stoichiometry in a eutrophied lake: Responses to a whole-lake food-web manipulation. Ecosystems 2000, 3, 293–307. [Google Scholar] [CrossRef]
- Mette, E.M.; Vanni, M.J.; Newell, J.M.; Gonzàlez, M.J. Phytoplankton communities and stoichiometry are interactively affected by light, nutrients, and fish. Limnol. Oceanogr. 2011, 56, 1959–1975. [Google Scholar] [CrossRef]
- Weber, M.J.; Brown, M.L. Effects of common carp on aquatic ecosystems 80 years after “Carp as a Dominant”: Ecological insights for fisheries management. Rev. Fish. Sci. 2009, 17, 524–537. [Google Scholar] [CrossRef]
- Richardson, M.J.; Whoriskey, F.G.; Roy, L.H. Turbidity generation and biological impacts of an exotic fish Carassius auratus, introduced into shallow seasonally anoxic ponds. J. Fish Biol. 1995, 47, 576–585. [Google Scholar] [CrossRef]
- Ruppert, J.L.; Docherty, C.; Neufeld, K.; Hamilton, K.; MacPherson, L.; Poesch, M.S. Native freshwater species get out of the way: Prussian carp (Carassius gibelio) impacts both fish and benthic invertebrate communities in North America. R. Soc. Open Sci. 2017, 4, 170400. [Google Scholar] [CrossRef]
- Hart, R.C. Zooplankton feeding rates in relation to suspended sediment content: Potential influences on community structure in a turbid reservoir. Freshw. Biol. 1988, 19, 123–139. [Google Scholar] [CrossRef]
- Gu, J.; Jin, H.; He, H.; Ning, X.Y.; Yu, J.L.; Tan, B.C.; Jeppesen, E.; Li, K.Y. Effects of small-sized crucian carp (Carassius carassius) on the growth of submerged macrophytes: Implications for shallow lake restoration. Ecol. Eng. 2016, 95, 567–573. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Z.; Jeppesen, E.; Taylor, W.D.; Rudstam, L.G. Effects of benthic-feeding common carp and filter-feeding silver carp on benthic-pelagic coupling: Implications for shallow lake management. Ecol. Eng. 2016, 88, 256–264. [Google Scholar] [CrossRef]
- Dantas, D.D.F.; Rubim, P.L.; de Oliveira, F.A.; da Costa, M.R.; de Moura, C.G.; Teixeira, L.H.; Attayde, J.L. Effects of benthivorous and planktivorous fish on phosphorus cycling, phytoplankton biomass and water transparency of a tropical shallow lake. Hydrobiologia 2018, 829, 31–41. [Google Scholar] [CrossRef]
- He, H.; Hu, E.; Yu, J.L.; Luo, X.G.; Li, K.Y.; Jeppesen, E.; Liu, Z. Does turbidity induced by Carassius carassius limit phytoplankton growth? A mesocosm study. Environ. Sci. Pollut. Res. 2017, 24, 5012–5018. [Google Scholar] [CrossRef]
- Hall, S.R.; Leibold, M.A.; Lytle, D.A.; Smith, V.H. Grazers, producer stoichiometry, and the light: Nutrient hypothesis revisited. Ecology 2007, 88, 1142–1152. [Google Scholar] [CrossRef]
- Holmroos, H.; Hietanen, S.; Niemistö, J.; Horppila, J. Sediment resuspension and denitrification affect the nitrogen to phosphorus ratio of shallow lake waters. Fundam. Appl. Limnol. 2012, 180, 193–205. [Google Scholar] [CrossRef]
- Niemistö, J.; Holmroos, H.; Pekcan-Hekim, Z.; Horppila, J. Interactions between sediment resuspension and sediment quality decrease the TN: TP ratio in a shallow lake. Limnol. Oceanogr. 2008, 53, 2407–2415. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, Q. Spatial distribution of sediment carbon, nitrogen and phosphorus and pollution evaluation of sediment in Taihu Lake Basin. Acta Sci. Circumst. 2014, 34, 3057–3064. (In Chinese) [Google Scholar]
- FAO. Cultured Aquatic Species Information Programme: Carassius carassius. Available online: http://www.fao.org/fishery/culturedspecies/Carassius_carassius/en (accessed on 13 March 2019).
- Yu, J.; Liu, Z.; He, H.; Guan, B.H.; Chen, F.Z.; Li, K.Y.; Zhong, P.; Teixeira-de Mello, F.; Jeppesen, E. Submerged macrophytes facilitate dominance of omnivorous fish in a subtropical shallow lake: Implications for lake restoration. Hydrobiologia 2016, 775, 97–107. [Google Scholar] [CrossRef]
- Gu, J.; He, H.; Jin, H.; Yu, J.L.; Jeppesen, E.; Nairn, R.W.; Li, K.Y. Synergistic negative effects of small-sized benthivorous fish and nitrogen loading on the growth of submerged macrophytes—Relevance for shallow lake restoration. Sci. Total Environ. 2018, 610–611, 1572–1580. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Liu, Z.; Jeppesen, E. Fish community assemblages changed but biomass remained similar after lake restoration by biomanipulation in a Chinese tropical eutrophic lake. Hydrobiologia 2014, 724, 127–140. [Google Scholar] [CrossRef]
- Liu, Z.; Hu, J.R.; Zhong, P.; Zhang, X.F.; Ning, J.J.; Larsen, S.E.; Chen, D.Y.; Gao, Y.M.; He, H.; Jeppesen, E. Successful restoration of a tropical shallow eutrophic lake: Strong bottom-up but weak top-down effects recorded. Water Res. 2018, 146, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Conley, D.J.; Paerl, H.W.; Howarth, R.W.; Boesch, D.F.; Seitzinger, S.P.; Havens, K.E.; Lancelot, C.; Likens, G.E. Controlling Eutrophication: Nitrogen and phosphorus. Science 2009, 323, 1014–1015. [Google Scholar] [CrossRef]
- Liu, E.S.; Liu, Z.; Bao, C.H. The changes of catches of Carassius auratus and relationship with environment in Lake Taihu, China. J. Lake Sci. 2007, 19, 345–350. (In Chinese) [Google Scholar]
- Jin, X.; Tu, Q. The Standard Methods for Obervation and Analysis in Lake Eutrophication; Environmental Science Press: Beijing, China, 1990. (In Chinese) [Google Scholar]
- SEPA. Analytical Methods for Water and Wasterwater Monitor, 4th ed.; Chinese Environmental Science Press: Beijing, China, 2002. (In Chinese) [Google Scholar]
- Hu, H.J. The Freshwater Algae of China: Systematics, Taxonomy and Ecology; Science Press: Beijing, China, 2006. (In Chinese) [Google Scholar]
- Zhang, Z.; Huang, X. Methods in Freshwater Plankton Study; Science Press: Beijing, China, 1991. (In Chinese) [Google Scholar]
- Wang, J.J. Fauna Sinica, Rotifer; Science Press: Beijing, China, 1961. (In Chinese) [Google Scholar]
- Chiang, S.C.; Du, N.S. Fauna Sinica, Crustacea; Science Press: Beijing, China, 1979. (In Chinese) [Google Scholar]
- Shen, J.R.; Du, N.S. Fauna Sinica, Crustacea, Freshwater Copepoda; Science Press: Beijing, China, 1979. (In Chinese) [Google Scholar]
- Huang, X.F. Survey Observation and Analysis of Lake Ecology; Standards Press of China: Beijing, China, 1999. (In Chinese) [Google Scholar]
- Wang, H.Z. Studies on Taxonomy, Distribution and Ecology of Microdrile Oligochaetes of China, with Descriptions of Two New Species from the Vicinity of the Great Wall Station of China, Antarctica; Higher Education Press: Beijing, China, 2002. (In Chinese) [Google Scholar]
- Tang, H. Biosystematic Study on the Chironomid Larvae in China (Diptera, Chironomidae); Nankai University: Tianjin, China, 2006. (In Chinese) [Google Scholar]
- Huynh, H.; Feldt, L.S. Estimation of the box correction for degrees of freedom from sample data in randomized block and split-plot designs. J. Educ. Stat. 1976, 1, 69–82. [Google Scholar] [CrossRef]
- Wahl, D.H.; Wolfe, M.D.; Santucci, V.J.; Freedman, J.A. Invasive carp and prey community composition disrupt trophic cascades in eutrophic ponds. Hydrobiologia 2011, 678, 49–63. [Google Scholar] [CrossRef]
- Badiou, P.H.J.; Goldsborough, L.G. Ecological impacts of an exotic benthivorous fish, the common carp (Cyprinus carpio L.), on water quality, sedimentation, and submerged macrophyte biomass in wetland mesocosms. Hydrobiologia 2015, 755, 107–121. [Google Scholar] [CrossRef]
- Andersen, T.; Hessen, D.O. Carbon, nitrogen, and phosphorus content of freshwater zooplankton. Limonol. Oceanogr. 1991, 36, 807–814. [Google Scholar] [CrossRef]
- Penczak, T. Phosphorus, nitrogen, and carbon cycling by fish populations in two small lowland rivers in Poland. Hydrobiologia 1985, 120, 159–165. [Google Scholar] [CrossRef]
- Schaus, M.H.; Vanni, M.J.; Wissing, T.E.; Bremigan, M.T.; Garvey, J.E.; Stein, R.A. Nitrogen and phosphorus excretion by detritivorous gizzard shad in a reservoir ecosystem. Limnol. Oceanogr. 1997, 42, 1386–1397. [Google Scholar] [CrossRef]
- Sterner, R.W.; George, N.B. Carbon, nitrogen, and phosphorus stoichiometry of cyprinid fishes. Ecology 2000, 81, 127–140. [Google Scholar] [CrossRef]
- Schaus, M.H.; Vanni, M.J. Effects of gizzard shad on phytoplankton and nutrient dynamics: Role of sediment feeding and fish size. Ecology 2000, 81, 1701–1719. [Google Scholar] [CrossRef]
- Chrzanowski, T.H.; Grover, J.P. The light: Nutrient ratio in lakes: A test of hypothesized trends in bacterial nutrient limitation. Ecol. Lett. 2001, 4, 453–457. [Google Scholar] [CrossRef]
- Golz, A.; Burian, A.; Winder, M. Stoichiometric regulation in micro- and mesozooplankton. J. Plankton Res. 2015, 37, 293–305. [Google Scholar] [CrossRef]
- Laspoumaderes, C.; Modenutti, B.; Elser, J.J.; Balseiro, E. Does the stoichiometric carbon: Phosphorus knife edge apply for predaceous copepods? Oecologia 2015, 178, 557–569. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Effects | Statistical Parameters | |||
|---|---|---|---|---|---|
| F | DF1 | DF2 | p | ||
| TN | Time | 2.442 | 4.392 | 17.569 | 0.081 |
| Carp | 0.004 | 1 | 4 | 0.956 | |
| Interaction | 0.324 | 4.392 | 17.569 | 0.873 | |
| TP | Time | 2.351 | 6 | 24 | 0.063 |
| Carp | 5.803 | 1 | 4 | 0.184 | |
| Interaction | 2.132 | 6 | 24 | 0.087 | |
| TDN | Time | 8.597 | 3.848 | 15.391 | 0.001 |
| Carp | 0.225 | 1 | 4 | 0.660 | |
| Interaction | 1.122 | 3.848 | 15.391 | 0.381 | |
| TDP | Time | 42.212 | 2.147 | 8.586 | <0.001 |
| Carp | 5.226 | 1 | 4 | 0.084 | |
| Interaction | 3.235 | 2.147 | 8.586 | 0.088 | |
| TN/TP | Time | 5.268 | 6 | 24 | 0.001 |
| Carp | 41.644 | 1 | 4 | 0.003 | |
| Interaction | 5.754 | 6 | 24 | 0.001 | |
| Irradiance | Time | 15.352 | 4.836 | 14.507 | <0.001 |
| Carp | 15.564 | 1 | 3 | 0.029 | |
| Interaction | 17.851 | 4.836 | 14.507 | <0.001 | |
| Irradiance/TP | Time | 2.282 | 6 | 18 | 0.082 |
| Carp | 14.202 | 1 | 3 | 0.033 | |
| Interaction | 3.584 | 6 | 18 | 0.016 | |
| Chl-a | Time | 3.408 | 6 | 24 | 0.014 |
| Carp | 0.060 | 1 | 4 | 0.819 | |
| Interaction | 1.502 | 6 | 24 | 0.220 | |
| Variables | Effects | Statistical Parameters | |||
|---|---|---|---|---|---|
| F | DF1 | DF2 | p | ||
| TS | Time | 12.556 | 6 | 24 | <0.001 |
| Carp | 15.345 | 1 | 4 | 0.017 | |
| Interaction | 10.217 | 6 | 24 | <0.001 | |
| IS | Time | 19.327 | 2.68 | 10.721 | <0.001 |
| Carp | 59.715 | 1 | 4 | 0.002 | |
| Interaction | 22.045 | 2.68 | 10.721 | <0.001 | |
| OS | Time | 6.201 | 6 | 24 | <0.001 |
| Carp | 0.024 | 1 | 4 | 0.885 | |
| Interaction | 0.825 | 6 | 24 | 0.562 | |
| IS% | Time | 5.625 | 6 | 24 | 0.001 |
| Carp | 283.205 | 1 | 4 | <0.001 | |
| Interaction | 8.053 | 6 | 24 | <0.001 | |
| C | Time | 15.448 | 6 | 24 | <0.001 |
| Carp | 116.536 | 1 | 4 | <0.001 | |
| Interaction | 1.384 | 6 | 24 | 0.261 | |
| N | Time | 10.544 | 6 | 24 | <0.001 |
| Carp | 42.629 | 1 | 4 | 0.003 | |
| Interaction | 1.664 | 6 | 24 | 0.173 | |
| P | Time | 5.177 | 2.412 | 9.646 | 0.026 |
| Carp | 38.684 | 1 | 4 | 0.003 | |
| Interaction | 0.861 | 2.412 | 9.646 | 0.472 | |
| C/P | Time | 5.154 | 4.622 | 18.49 | 0.005 |
| Carp | 195.240 | 1 | 4 | <0.001 | |
| Interaction | 1.833 | 4.622 | 18.49 | 0.159 | |
| C/N | Time | 15.732 | 5.113 | 20.454 | <0.001 |
| Carp | 7.811 | 1 | 4 | 0.049 | |
| Interaction | 4.162 | 5.113 | 20.454 | 0.009 | |
| N/P | Time | 1.100 | 5.303 | 21.212 | 0.391 |
| Carp | 71.531 | 1 | 4 | 0.001 | |
| Interaction | 0.753 | 5.303 | 21.212 | 0.600 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, H.; Han, Y.; Li, Q.; Jeppesen, E.; Li, K.; Yu, J.; Liu, Z. Crucian Carp (Carassius carassius) Strongly Affect C/N/P Stoichiometry of Suspended Particulate Matter in Shallow Warm Water Eutrophic Lakes. Water 2019, 11, 524. https://doi.org/10.3390/w11030524
He H, Han Y, Li Q, Jeppesen E, Li K, Yu J, Liu Z. Crucian Carp (Carassius carassius) Strongly Affect C/N/P Stoichiometry of Suspended Particulate Matter in Shallow Warm Water Eutrophic Lakes. Water. 2019; 11(3):524. https://doi.org/10.3390/w11030524
Chicago/Turabian StyleHe, Hu, Yanqing Han, Qisheng Li, Erik Jeppesen, Kuanyi Li, Jinlei Yu, and Zhengwen Liu. 2019. "Crucian Carp (Carassius carassius) Strongly Affect C/N/P Stoichiometry of Suspended Particulate Matter in Shallow Warm Water Eutrophic Lakes" Water 11, no. 3: 524. https://doi.org/10.3390/w11030524
APA StyleHe, H., Han, Y., Li, Q., Jeppesen, E., Li, K., Yu, J., & Liu, Z. (2019). Crucian Carp (Carassius carassius) Strongly Affect C/N/P Stoichiometry of Suspended Particulate Matter in Shallow Warm Water Eutrophic Lakes. Water, 11(3), 524. https://doi.org/10.3390/w11030524