Abstract
Foundation species provide habitat to other organisms and enhance ecosystem functions, such as nutrient cycling, carbon storage and sequestration, and erosion control. We focus on freshwater wetlands because these ecosystems are often characterized by foundation species; eutrophication and other environmental changes may cause the loss of some of these species, thus severely damaging wetland ecosystems. To better understand how wetland primary producer foundation species support other species and ecosystem functions across environmental gradients, we reviewed ~150 studies in subtropical, boreal, and temperate freshwater wetlands. We look at how the relative dominance of conspicuous and well-documented species (i.e., sawgrass, benthic diatoms and cyanobacteria, Sphagnum mosses, and bald cypress) and the foundational roles they play interact with hydrology, nutrient availability, and exposure to fire and salinity in representative wetlands. Based on the evidence analyzed, we argue that the foundation species concept should be more broadly applied to include organisms that regulate ecosystems at different spatial scales, notably the microscopic benthic algae that critically support associated communities and mediate freshwater wetlands’ ecosystem functioning. We give recommendations on how further research efforts can be prioritized to best inform the conservation of foundation species and of the freshwater wetlands they support.
Keywords:
foundation species; wetlands; algae; vascular plants; nonvascular plants; hydrology; phosphorus; salinity; fire 1. Introduction
1.1. The Importance of Foundation Species in Freshwater Wetlands
Foundation species are abundant organisms that facilitate whole communities of other species by creating complex habitat, ameliorating local biotic and abiotic stressors, and enhancing resource availability [1,2,3]. These species also regulate nutrient cycling by storing nutrients, trapping debris and supporting gas exchange, facilitate the growth and survival of other plant and animal species, provide refuge from predators and competitors, alter microclimate and water availability, and stabilize substrata [4,5,6,7]. As a result of these community impacts and their dominance, foundation species are key drivers of biodiversity and ecosystem functioning in terrestrial, marine, and freshwater ecosystems. Foundation species often have species-specific traits and physiological adaptations enabling them to best cope with specific environmental conditions and to facilitate the growth and survival of other species that are less tolerant to such conditions [8]. Although the concept of foundation species is well-recognized in marine and coastal ecosystems, such as coral reefs, kelp forests, seagrass meadows, salt marshes and mangroves [9,10,11], it has been less commonly recognized in freshwater wetlands where vascular plants and benthic algae often fulfill foundational roles [5,12].
In wetlands, dominant primary producers create distinct (microtopographic) environments, such as ridges/hummocks, slough/hollows, and benthic mats. In addition, in coastal areas where wetlands form at the interface of terrestrial and marine systems, hydrology, sediment delivery, nutrient availability, and top-down forcing influence the identity and status of the foundations species that structure salt marshes and mangrove forests. As global change is causing ecosystem state changes and productivity declines in freshwater wetlands around the world [13], it is now urgent to explore the degree to which such state changes may be caused by the loss of foundation species or changes in their composition. By better understanding the attributes that support foundation species in freshwater wetlands, ecosystem changes could be managed in an anticipatory way to preserve these species’ roles. Because freshwater wetlands (especially oligotrophic ones) are often characterized by their foundation species, they serve as an ideal model of how such species control and may stabilize ecosystem structure and functions. Many nonlinear and abrupt changes in ecosystems are connected to the loss of foundation species [3]. Here, we summarize available evidence to motivate the importance of preserving these species and their foundational roles. Foundation species will ultimately contribute to maintaining the resilience of wetland ecosystems facing numerous and severe threats around the world [14].
1.2. Drivers and Factors Affecting Foundation Species in Freshwater Wetlands
To understand the role of foundation species in ecosystems and forecast consequences of changes in their distribution, dominance and/or roles, we need to quantify these species’ environmental preferences and tolerances (niches) and the effects of interspecific interactions on their spatial distribution. In freshwater wetlands, the fundamental niche of foundation species may be defined by (i) hydrologic and water quality gradients; (ii) long-term presses (i.e., chronic, gradual disturbances), such as eutrophication, infrastructure that diverts water flows, and directional climate change (i.e., increased saltwater intrusion rates and fire and drought frequency and/or intensity [10,15]); and (iii) short-term pulses (i.e., acute events) that episodically disturb ecosystem structure, such as specific droughts, fires, cold spells, and storms [16,17,18]. The realized ecological niche of these species is therefore determined by how they react to these gradients, presses, and pulses.
There is debate over whether foundation species may better facilitate other species under high physical stress and/or consumer pressure [19] or under moderately stressful conditions [15,20,21,22]. In some cases, a few species may be dominant in nutrient-poor and disturbance-prone ecosystems, as in the case of mangroves and salt marsh graminoids [10] in tidal coastal wetlands and diatom taxa in U.S. streams [23]. In other cases, algal species may be dominant in wetlands with high nutrient levels or in warmer climates [24], and plant species, such as cattail, may be dominant in areas with high phosphorus (P) concentrations [25]. To better predict how freshwater wetlands will change on different magnitudes and timescales, we need to know how their foundation species facilitate other organisms and how they are responding to environmental drivers [26]. Here we focus on hydrology, nutrients, and other environmental conditions such as fire and salinity, and on how press and pulse drivers may impact foundation species dynamics in these aquatic ecosystems.
1.3. Foundation Species Changes and Loss in Freshwater Wetlands
Wetlands provide valuable ecosystem services, such as food and drinking water, erosion control, carbon storage and sequestration, water purification, and flood and storm protection [27], which are often controlled, at least in part, by their foundation species. These ecosystem services are threatened by overexploitation of natural resources, land reclamation for agriculture, reservoir and dike construction, invasive species, and climate change [14,28]. Foundation species in various freshwater wetlands are particularly stressed by climate change that is altering temperature, drought frequency, fire susceptibility, nutrient availability and, in coastal settings, increasing saltwater intrusion [16]. In particular, eutrophication often alters the productivity and composition of foundation species, leading to changes in fundamental ecosystem processes that result in regime shifts between alternative stable states, for example from a plant-dominated to an algae-dominated regime in shallow lakes [29]. To guide conservation actions around these shifts in an anticipatory way, we need to understand the environmental constraints on foundation species and how a loss of foundation species may cascade to influence other ecosystem attributes, or lead to replacement by other foundation species through a regime shift. A better understanding of wetland foundation species dynamics can help with conservation planning for the future under climate change, as these species’ traits will define whether ecosystems can adapt, persist, or decline [13], or whether new communities with or without adapted foundation species can form.
1.4. Focus, Goal, and Objectives
In this review, we focus on the species identity, dominance, and appearance and roles of primary producer foundation species in freshwater wetlands, especially oligotrophic ones where organisms cope with nutrient scarcity by means of specific traits and adaptations that enable them to outnumber other species. We review the relative dominance of foundation species, from microalgae to herbaceous vascular plants and trees, and their responses to hydrology (see Section 2.1 and Section 2.2), nutrients and other environmental factors (e.g., fire and salinity) in freshwater wetlands located at different latitudes [30]. Many of these wetlands are typically characterized by flat topography; however, the foundation species living there biogenically create landforms that may be patterned by water flow [31,32]. We evaluate ways in which changes in the environmental conditions that support foundation species cascade to influence the abundance of other species and how these changes may influence ecosystem functions. We conclude with a discussion of how future scenarios of changing climate and disturbance regimes may influence foundation species and their functions in freshwater wetlands, implications for conservation and future research directions (see Section 3.1, Section 3.2 and Section 3.3). Through this analysis, we developed an evidence-based case for the integration of foundation species into our understanding of ecosystem organization and functioning in imperiled wetlands and for a more explicit consideration of these species in wetland conservation and management. Our objectives are to synthesize available knowledge on (i) which foundation species provide habitat to other species and support other ecosystem functions across freshwater wetlands; (ii) how foundation species dominance varies in relation to hydrology, nutrients, and other environmental factors; and (iii) how foundation species interact with other species in response to ongoing environmental changes. This review will help us better understand how foundation species control freshwater wetlands’ ecosystem structure and function across environmental gradients, how these species can be conserved and how they can support wetland restoration.
2. Foundation Species Dynamics in Freshwater Wetlands
We reviewed 154 publications discussing (explicitly or implicitly) how dominant species of plants and algae play foundational roles in freshwater wetlands (of which 95 publications on subtropical wetlands and 55 on boreal and temperate wetlands; 34 on algae, 54 on trees, and 67 on other plants; Table 1 and Table S1). Foundation species are more or less dominant at different points of the hydrology and nutrient gradients in subtropical wetlands (Figures 2 and 3) and boreal/temperate wetlands (Figures 5 and 6) and cope with water and nutrient scarcity and other environmental challenges in different ways and at different scales depending on their organization level, i.e., algae, nonvascular plants, and vascular plants (e.g., sedges and trees).
2.1. Foundation Species in Tropical and Subtropical Wetlands
This review focuses on a pivotal subtropical wetland, the Florida Everglades (USA), but also discusses other marshes in Belize, Mexico, Jamaica and forested wetlands in Louisiana and Puerto Rico. The Everglades is a rainfall-dependent ‘river of grass’ in South Florida [33] that flows in a highly regulated way from Lake Okeechobee to the Gulf of Mexico and Florida Bay due to agricultural and urban development [34]; a vast restoration program is ongoing to increase this wetland’s water quantity, flow and quality. We found that at least five species play foundation roles in the freshwater Everglades marshes and in similar karstic wetlands in the Caribbean region and other wetlands in the southeastern U.S.: two cyanobacteria (Schizothrix calcicola and Scytonema hofmanii), one diatom (Mastogloia calcarea), one sedge (Cladium jamaicense, hereafter referred to as sawgrass), and one tree species (Taxodium distichum, hereafter referred to as bald cypress) (Figure 1). In disturbed habitats, Typha domingensis (hereafter referred to as cattail) may also perform a foundation species role.


Figure 1.
Examples of subtropical wetland ecosystems (left column) and the associated foundation species (right column). (A) Microbial (periphyton) mats in the ridge and slough landscape of the Florida Everglades (photo by Franco Tobias). (B) Scanning electron micrograph of Mastogloia calcarea, a diatom foundation species in microbial mats restricted to the South Florida and Caribbean regions (photo by Sylvia Lee from a sample collected in Shark River Slough, Everglades National Park; scale bar: 10 µm [35]). (C) Sawgrass-dominated ridge and slough landscape in the Everglades (photo by Luca Marazzi). (D) Sawgrass in the Everglades (photo by staff at Florida International University (FIU)’s Geographical Information System Lab). (E) Bald cypress dome (in the background) in the Florida Everglades with visible encroachment of scrub mangroves (in the foreground) due to propagule seeding by the 1960 Hurricane Donna and saltwater intrusion caused by sea level rise (photo by Luca Marazzi). (F) Bald cypress trees in Everglades National Park (photo by staff at FIU’s South Florida Terrestrial Ecology Lab).
2.1.1. Foundation Species Interactions with Hydrology
Algae
In freshwater Everglades and other karstic Caribbean marshes, the dominant microscopic primary producers are the cyanobacteria Scytonema hofmanii and Schizothrix calcicola and the diatoms Encyonema evergladianum, Encyonopsis microcephala, Mastogloia smithii, Nitzchia palea var. debilis, and Brachysira neoexilis [5,36,37,38,39,40,41]. Of these, the cyanobacteria S. hofmanii and S. calcicola and the diatom Mastogloia calcarea (described by Lee et al. [42] and previously called M. smithii [36]) play particular foundation roles by producing mucilage (extracellular polysaccharides—EPS) and calcium carbonate that bind the microbial mats together [43]. These microbial (periphyton) mats act as sponges that retain soil moisture, protecting algae and other plants (particularly their seeds), bacteria, fungi, and aquatic infauna (i.e., midge larvae, amphipods, crayfish, and snails) from drought and grazing/predation [5,44]. Thick and cohesive mats with high biomass prevail in seasonally flooded areas while loose mats with low biomass prevail in permanently flooded areas [5,39,45]. While M. calcarea tends to be more dominant in shorter hydroperiod sites, S. hofmanii and S. calcicola may be dominant in algal biovolume terms (an estimate of biomass through geometric calculations of cell volumes) in short to long hydroperiod sites [5,40] (Figure 2). These foundation species tolerate water scarcity (desiccation stress), UV radiation (e.g., through photoprotective pigments), and high temperatures [5,40,46] (Figure 2). In shallow oligotrophic marshes in northern Belize, other cyanobacteria, such as Leptolyngbya, appear to join the foundation species discussed above to form a microbial consortium that provides habitat to other microbial and protozoan colonizers [47]. In the El Edén wetland (Yucatan, Mexico), the coccoid cyanobacteria Cyanokybus, Aphanothece, Gloeocapsa, and the filamentous cyanobacteria Stigonema are most resistant to desiccation, as they protect themselves in benthic mats from dehydration by secreting mucilage that retains water [48].
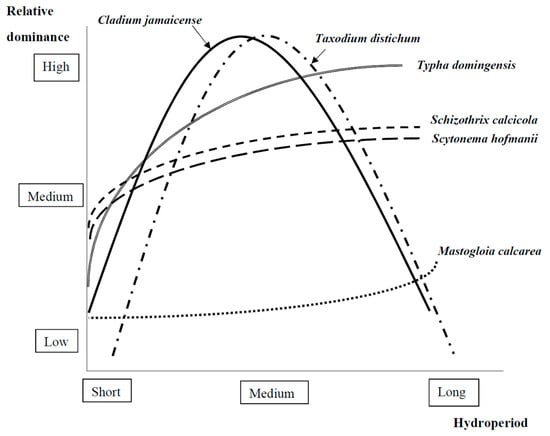
Figure 2.
Examples of observed relationships between relative dominance of different foundation species and hydroperiod in the freshwater Everglades ecosystem based on the literature reviewed (see Section 2.1.1 and refer to Marazzi et al. [40] for cyanobacteria and diatom species; Ross et al. [53], Todd et al. 2010 [55], Sah et al. [56], Toth [57] and Busch et al. [58] for sawgrass; Newman et al. [25] for cattail; and Allen et al. [68], Megonigal & Day [69], Shanklin & Kozlowski [70] and Elcan J.M.; Pezeshki [77] for bald cypress). Scytonema hofmanii and Schizothrix calcicola are difficult to identify through microscopy and hence some studies refer to them as Scytonema sp. and Schizothrix sp. (e.g., Marazzi et al. [40]).
Herbaceous Vascular Plants
In karstic freshwater marshes of the Caribbean, sawgrass is the most dominant plant species, and in the Everglades, historically comprised 50–70% of the total vegetation cover [49]. Sawgrass is a foundation species because it provides a critical source of organic matter that structures the characteristic ridge and slough landscape [25]. Moreover, this sedge supports abundant benthic algae that grow on its stems as epiphytic accumulations locally called sweaters as they appear thick, fluffy, and protective [5]. In seasonally flooded short hydroperiod prairies underlain by marl, sawgrass is a major component of vegetation that provides habitat to the Cape Sable seaside sparrow, to other endangered nesting birds and to their prey organisms, such as amphibians and invertebrates [34]. Sawgrass is dominant in subtropical karstic wetlands because it can reproduce by various modes [50] and it regrows rapidly after fire [51] and with high water levels [52]. This marsh species tolerates hydroperiod ranging from 2 to 11 months (i.e., from seasonally flooded prairies to semipermanently flooded marshes [53,54,55]) and reaches highest abundance with intermediate flooding [56] (Figure 2). The foundation species role of sawgrass is most evident in drier marshes (i.e., inundated for 6–10 months [53]) where it forms dense and tall sawgrass stands (>2 m) [57,58], while sparse and short sawgrass (<2 m height) is dominant in wetter marshes [59]. By contrast, in deeper water, sawgrass is part of a mixed plant community with floating (e.g., water lily, Nymphaea odorata) and submerged species (e.g., spikerush, Eleocharis cellulosa) that find space among sawgrass tussocks [60]. Sawgrass is dominant in other freshwater wetlands in the Caribbean, such as the Sian Ka’an Biosphere Reserve in Mexico [61], the Grande-Terre in Lesser Antilles [62], and the New River Lagoon in Belize [63], especially with shallow to intermediate water depth [64]. An undesirable foundation species, at least in the Everglades, is cattail, an herbaceous perennial plant that becomes dominant with water depth higher than 1.2 m [65] (Figure 2) where TP concentrations are higher due to water diversion and impoundment closer to agricultural discharges (see Section 2.1.2).
Trees
In freshwater forested wetlands (i.e., swamps) of southeastern North America, bald cypress is an important foundation species; its presence and density control sediment and nutrient flows and water table height, whilst influencing the community structure of associated plants and animals [12]. In particular, bald cypress provides habitat for native species, such as the American alligator, leopard, grass frogs, beavers, otters, the endangered Florida panther, and nesting birds [34]. The removal of bald cypress for logging purposes can drastically change hydrology, reduce primary productivity, and increase sedimentation in Florida and Alabama (see Sun et al. [66] and references therein), thus demonstrating this species’ foundational role. Optimal conditions for bald cypress are present in seasonally flooded areas in Everglades National Park, for example in the western and eastern part of Shark River Slough [67] (Figure 2). This foundation species is relatively flood tolerant as it is best adapted to periodic flooding (optimal water depth < 1 m), while its seed germination and growth can be reduced in deep, permanently flooding areas [68]. Experimental flooding for 14 consecutive weeks at 2 cm above the soil surface significantly decreased seedling height, leaf area, and total dry mass [69]. Permanent flooding can inhibit bald cypress stomatal conductance [70], root elongation [71], and RuBisCO activity [72] through anoxia stress. However, this foundation species and other conifers cope well with intermittent flooding and related anoxia by producing adventitious roots and through intercellular spaces that help carry oxygen from plant structures above the water table to the roots [73,74,75]. Because of these properties, bald cypress has been recommended for vegetation restoration projects in riparian zones [76]. As bald cypress prefers moderate flooding, it is also sensitive to drought [73]; its net photosynthetic rate was shown to decrease with moderate drought stress [77] while germinating seedling density was decreased by repeated drought conditions [78].
2.1.2. Foundation Species Interactions with Nutrients
Algae
Periphyton mats play a foundation role in regulating nutrients in karstic wetlands, including the Everglades, the Sian Ka’an Biosphere Reserve in Quintana Roo (Mexico), the New River Lagoon in Orange Walk (Belize), and the Broad River in the Black River Morass (Jamaica) [38]. In the Caribbean marshes above-mentioned, Mastogloia calcarea has an optimum TP between 140 and 240 µg/g [39,79], which indicates oligotrophic conditions [80]; in the Everglades this species is more dominant in areas with TP < 300 µg/g (Marazzi and Gaiser, unpublished data; see [40]) (Figure 3). Both Scytonema hofmanii and Schizothrix calcicola are more dominant with lower TP [40] (Figure 3). In the Belize wetlands, benthic mats with abundant foundation species of filamentous cyanobacteria (i.e., Scytonema sp. and Schizothrix sp. [47]) and diatoms (i.e., M. calcarea [38]) preserve nutrients and thus increase resource use efficiency [81]. However, where and when P concentrations increase, for example downstream of intensively cultivated areas, heterotrophic bacteria activity increases as they efficiently exchange dissolved, adsorbed, and organic P [82], causing a shift from cohesive mats to loose mats [43]. This cohesive structural breakdown comes with a loss of the foundation species that create the mat, as these are replaced by green algae and by more generalist diatom species; although these algae are more palatable to invertebrates [83], microbial mats become loose, net ecosystem heterotrophy increases, carbon is lost and, ultimately, a transition to a nutrient-enriched ecosystem state takes place [79,84].
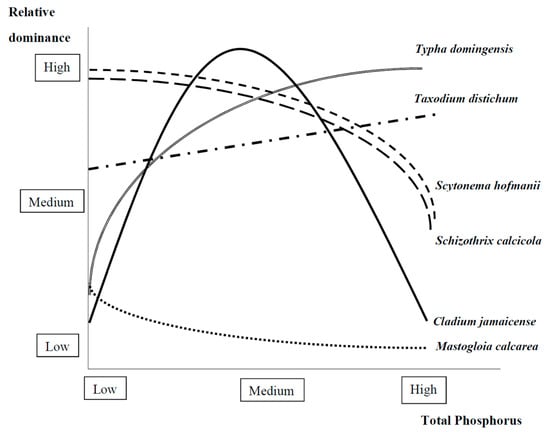
Figure 3.
Examples of observed relationships between relative dominance of different foundation species and total phosphorus (TP) concentrations in the freshwater Everglades based on the literature reviewed (see Section 2.1.2 and refer to Marazzi et al. [40] and Gaiser et al. [79] for cyanobacteria and diatom species; Steward & Ornes [87,88], Davis [89], Noe et al. [90] and Richardson et al. [91] for sawgrass; Grace [65], Richardson et al. [91], Newman et al. [97], Davis & Ogden [98], and Hagerthey et al. [99] for cattail; and Effler et al. [100], Krauss et al. [101], Nyman & Lindau [102], Dickson & Broyer [103], Dickson et al. [104], Effler & Goyer [105] and Day et al. [106] for bald cypress).
Herbaceous Vascular Plants
In karstic wetlands in the Caribbean, such as in the Northern Belize Rio Hondo and New River marshes, sawgrass and cattail are more dominant with lower and higher extractable soil P concentrations, respectively [85]. The Everglades is an icon for such karstic wetlands that share the same foundation species and other ecological hydrological, biogeochemical, and characteristics [86]. In this wetland, sawgrass often dominates space and resources and becomes very dense with very low (limiting) P concentrations [50,87,88,89,90,91] (Figure 3). In deep water habitats with high P cattail grows fast, and allocates little mass to its roots thus outcompeting sawgrass and supporting high peat accumulation rates [92]. Cattail dominance leads to a further increase in P-availability so that this species feeds back positively on its own success and creates monocultures [50,93]. In these high-P habitats, benthic algal mats dominated by diatom and cyanobacteria foundation species cannot grow because of shading [94] or allelopathy [95]. Similar to other species growing in infertile soils [96], sawgrass allocates the little P available to its leaves, roots, and rhizomes whilst storing more nutrients in shoot bases [50,97]. In the 1970s and 1980s, an increase in soil and water P concentrations [98] triggered the replacement of sawgrass by cattail—an autochthonous sedge [99]—which is more competitive in high-nutrient environments [95]. By then, decades of agricultural development and disturbance events, such as fire, drought, or freeze events, had taken a severe environmental toll on the Everglades ridge and slough landscape [25]. Moreover, while small experimental P additions have been shown to increase shoot length and dry matter production, large P additions inhibited sawgrass growth [88]. Where cattail takes over in the Everglades landscape, it replaces more diverse plant communities dominated by sawgrass and forms impenetrable monocultures [95] that take over primary production and nutrient cycling, and thus play an undesirable foundation species role.
Trees
Bald cypress showed positive responses to nutrient increases in both field studies (e.g., [100,101,102]) and greenhouse studies (e.g., [103,104]) (Figure 3). Nutrient enrichment caused by municipal wastewaters increased long-term growth of bald cypress [105] and fertilization increased its diameter growth under flooding stress [104,106]. In oligotrophic environments, bald cypress can become a nutrient sink when nutrients are added from adjacent landscapes through increased growth and leaf nutrient content [100]. However, the type of fertilizer (reduced or oxidized form) may have different effects on growth: for instance, fertilization with urea (a reduced source of nitrogen) results in more growth than the application of nitrate (an oxidized source of N) [107,108]. In Southern Louisiana, bald cypress exhibited a positive short-term growth effect of nutrient addition that was however offset by a long-term negative effect of prolonged inundation and subsidence [109].
2.1.3. Foundation Species Interactions with Other Environmental Conditions
Algae
The abundance and distribution of algal foundation species in karstic microbial mats are strongly regulated by pH, conductivity, and salinity, but also by UV exposure and temperature [5]. In coastal regions where saltwater intrusion is occurring, changes in the dominance of foundation species is altering the integrity of periphyton mats and the roles they play [110,111]. Specifically, the diatom foundation species Mastogloia calcarea tolerates increased salinity of 3–10 ppt (as compared to 2.7–6.5 ppt ambient) and then rapidly declines in abundance, with ecosystem consequences [5,40,79,111]. The ecosystem metabolism and functioning of coastal wetlands are increasingly dependent on the balance of freshwater flow and sea level rise [112], and the way that this balance influences foundation species.
Herbaceous Vascular Plants
Sawgrass has a complex relationship with fire and salinity. In these marshes, frequent fires help other plants grow in dense sawgrass areas, such as Muhlenbergia capillaris (hereafter muhly grass) in short hydroperiod marl prairies [113], while fires suppression promotes the growth of woody plants [62]. Although sawgrass is not killed by fire in the Everglades, sawgrass can show significant mortality when fire is immediately followed by flooding [51,114]. For instance, the cover of sawgrass and other codominant species, including muhly grass, had not returned to pre-burn levels in short hydroperiod marl prairies even after five years due to a 2005 fire followed by >30 cm flooding that lasted several days [113]. Although sawgrass can grow in areas where salinity exceeds 10 ppm [62], saltwater intrusion is causing landward encroachment of mangroves and the displacement of freshwater plants, including sawgrass [115,116]. This ongoing displacement of sawgrass by mangroves is predicted to have significant effects on fish and bird habitats, coastal protection from storms, nutrient cycling, and water quality [10]. Therefore, fire and salinity are tolerated by sawgrass, but only to the extent that woody plant and mangrove encroachment are limited, respectively.
Trees
Various plant species dominate forested wetlands in subtropical regions, such as the flowering plants Eugenia spp. in Malaysia (a member of the Myrtaceae family [117]) and Pterocarpus officinalis in Puerto Rico (a member of the Fabaceae family [118]), species of palms in Puerto Rico floodplains [119], and hardwood species assemblages [120]. These swamps are increasingly exposed to flooding and saltwater intrusion caused by sea level rise that result in decreased growth and increased mortality due to anoxia and salinity stress [121,122,123]. Bald cypress can tolerate salinity of 3 to 6 ppt through osmotic adjustments [124], but its basal area was halved by a chronic salinity increase from 0.7 to 1.3 ppt over a year (from 87 m2 ha−1 to 40 m2 ha−1 [101]). This species is adapted to long-term cycles of sedimentation, weathering, deposition, and erosion caused by sea level fluctuations [125]. High salinity causes osmotic stress and decreased stomatal conductance [126], as indicated by decreased water use efficiency from 100 kg/day on a freshwater site to 23.9 kg/day on a saline water site [127,128]. In tidal forested wetlands, increased soil water salinity negatively reduces nutrient uptake in bald cypress [128,129,130,131,132], which in turn may decrease nitrogen and P burial [133].
2.2. Foundation Species in Boreal and Temperate Wetlands
In boreal and temperate wetlands, Sphagnum moss, pines and beeches, and algae, such as benthic diatoms, perform a foundation species role (Figure 4). We draw information on foundation species dynamics from field and experimental studies in freshwater wetlands, especially peatlands (i.e., wetlands with almost permanent water saturation and consequent anoxic conditions accumulating dead organic material formed in situ [134]) in Alaska, Canada, Sweden, Finland, Ireland, Japan, and New Zealand. These ecosystems include rain-fed (ombrotrophic) wetlands and groundwater-fed (minerotrophic) wetlands.


Figure 4.
Examples of boreal and temperate wetland ecosystems (left column) and the associated foundation species (right column). (A) Ponded wetlands in Wood Buffalo National Park (Canada) (photo by Stephen Davis III). (B) Scanning electron micrograph of Cymbella pusilla (now called Navicymbula pusilla, photo by Marina Potapova from a sample from Wolf Creek, Missouri, USA; scale bar: 5 µm [135]). (C) Boreal wetland in the Northwest Territories, Canada (Boreal Songbird Initiative; photo by Chad Delany). (D) Sphagnum mosses are widely distributed in boreal wetlands and elsewhere (photo taken by Todd Crowl in Upper Michigan). (E) Aerial photograph of a patterned peatland (the Great Vasyugan Bog, Siberia, Russia; photo by Maarten Eppinga), where tree species such as Betula pubescens and Pinus sylvestris dominate elevated patches. (F) Close-up view of an elevated ridge with B. pubescens with P. sylvestris in the background (photo by Maarten Eppinga).
2.2.1. Foundation Species Interactions with Hydrology
Algae
In coastal wetlands of Lakes Michigan and Huron, N2-fixing benthic diatoms and cyanobacteria, such as Epithemia, Rhopalodia, Anabaena, and Calothrix, can grow in high abundances on artificial substrata despite the high nutrient availability promoted by flowing water [136]. In ponds in Wood Buffalo National Park (Canada), the dominant diatoms were the benthic Cymbella pusilla, Mastogloia smithii, and Nitzschia amphibia species adapted to high conductivity and alkalinity that feed invertebrates that are in turn eaten by the protected whooping cranes [137]. Diatom assemblages form and develop in response to limiting nutrient availability and to the physical architecture of the benthic biofilm/mat they live in [138]. In shallow rivers in New Zealand, the diatom Didymosphenia geminata forms thick epilithic blooms and facilitated high densities of invertebrates, such as Oligochaeta, Chironomidae, Cladocera, and Nematoda [139]. Therefore, benthic algae may play a foundation species role as habitat creators by building biofilms on substrata where other producers and consumers live and/or as nutrient cyclers through food web dynamics and biochemical processing of nutrients.
Vascular and Nonvascular Plants
In boreal and temperate wetlands, the distance between the peatland surface and the water table is an important determinant of productivity and decomposition [140]. Hollow moss species perform well when the water table is near or above the surface, for example Sphagnum cuspidatum, S. cuspidatum var. majus, S. tenellum, and S. apiculatum [140,141,142,143], as well as bog pool species, such as Warnstorfia fluitans [144] (Figure 5). Compared to vascular plants, these peat mosses have low productivity [145]. Hence, only small amounts of precipitation are needed for Sphagnum-dominated bogs to maintain or decrease the distance to the water table [31]. Moreover, in systems where Sphagnum mosses dominate, peat tends to have low porosity so that little water can create waterlogged conditions, which in turn increases water table height [31,146]. These observations suggest that Sphagnum mosses promote higher water tables, providing aquatic habitats for water mites such as Arrenurus stecki, Piersigia intermedia, and Zschokkea oblonga [31,147]. Hummock species of Sphagnum, such as S. magellanicum and S. fuscum, occupy drier niches along the hydrological gradient [140,148] and can also play a foundational role once they overtake other moss species and subsequently provide a scaffold in the hummock that vascular plants colonize [149]. Furthermore, analogously to what trees do, the grass Sasa palmata and the shrub Betula nana can lower the water table by increasing the rate of evapotranspiration [148,150,151,152]. These various foundation species effects on wetland hydrology are evidence of their functional role at low to mid latitudes.
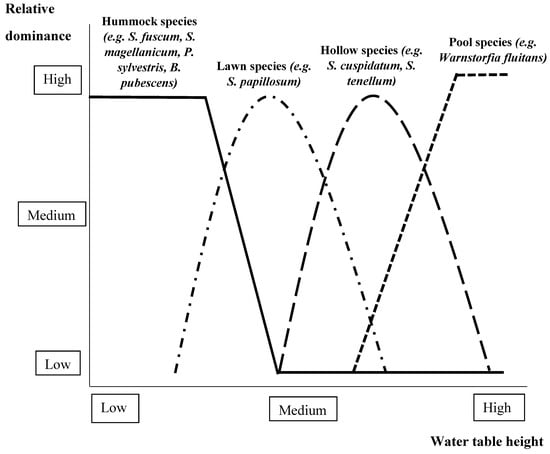
Figure 5.
Examples of relationships observed in the literature reviewed between relative dominance of different foundation species and water table height (lower/higher water table height means drier/wetter conditions) in boreal and temperate peatlands (see Section 2.2.1 and refer to Belyea & Clymo [140], Barkman [141], Robroek et al. [142], Hájek & Beckett [143], Vitt [144], Eppinga et al. [148], Frankl & Schmeidl [150], Takagi et al. [152], Lamers et al. [153], Rutter [158], and Limpens et al. [159]). S. = Sphagnum, P. = Pinus, B. = Betula.
Trees
Scots pine (Pinus sylvestris), several species of birch (Betula spp., e.g., B. papyrifera, B. pubescens, and B. pendula), and of spruce (e.g., Picea abies and P. mariana) are among the trees that can be dominant and function as foundation species in boreal and temperate wetlands [153,154,155]. These tree species require a relatively low water table: as higher water tables lead to substantial dieback of seedlings [156,157] (Figure 5). Invasion of trees into wetlands may therefore depend on the presence of hummocks that provide sufficiently dry conditions for tree seedlings [155,156]. Importantly, once established, tree species can actively modify their habitat in ways that stimulate water table drawdown [31]. For example, the canopy of tree species may increase the amount of precipitation that is intercepted, which creates dryer soil conditions [158]. Furthermore, P. sylvestris and B. pubescens may induce a lowering of the water table by increasing the rate of evapotranspiration [148,150,152].
2.2.2. Foundation Species Interactions with Nutrients
Algae
In the Stordalen Mire in northern Sweden, several species of the diatom genera Eunotia and Pinnularia and associated sedges create a shallow peat due to biogenic silica deposition and nutrient release following surface vegetation decay; therefore, diatoms and sedges facilitate one another [160]. In riparian wetlands in Alaska, benthic diatoms (e.g., Nitzschia) and euglenoids (e.g., Euglena and Trachelomonas) thrive in habitats with high levels of nutrients released by permafrost thawing and little light because of their ability to migrate towards the surface [161]. In these wetlands’ biofilms, the algae that dominate in abundance or biomass terms and/or are the most edible likely play a foundation role by creating habitat for and feeding invertebrates, such as snails [161]. Within the temperate and boreal latitudes of the northern hemisphere, certain areas are projected to experience greater increases in evapotranspiration than precipitation in the next century [162]. As such changes may yield drier conditions for part of the growing season, the foundation role of algae could become more important. For example, in various wetland ecosystems drought has already increased benthic algal production and, consequently, organic matter due to high temperature and light penetration, slow water flow, and minimal shade [163,164].
Vascular and Nonvascular Plants
In boreal and temperate peatlands, the competition between Sphagnum mosses and vascular plants is typically asymmetric [165]. Specifically, each Sphagnum moss species fixes nutrient inputs from atmospheric deposition [76,149,166,167], thus supporting their own growth [146] and that of other Sphagnum species [142]. However, whether Sphagnum functions as an effective filter for atmospheric nutrients depends on the level of atmospheric deposition [153]. Specifically, under low levels of atmospheric nitrogen deposition (<12 kg N.ha−1.yr−1), peatlands may be dominated by species, such as S. magellanicum, effectively absorbing nitrogen [153,159] (Figure 6). Under intermediate levels of atmospheric nitrogen deposition (12–18 kg N.ha−1.yr−1), strong competitors such as S. recurvum may dominate over other species, as nitrogen may no longer be limiting [153,168] (Figure 6). Under high nitrogen deposition (>18 kg N.ha−1.yr−1), the filter function of Sphagnum largely fails and higher growing vascular plant species like the grass Molinia caerulea may become dominant, suppressing Sphagnum species [153,169,170] (Figure 6).
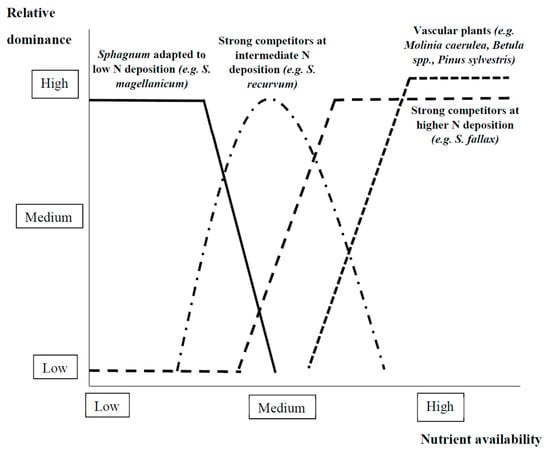
Figure 6.
Examples of relationships observed in the literature reviewed between relative dominance of different foundation species and nutrient availability in boreal and temperate wetlands (see Section 2.2.2 and refer to Lamers et al. [153], Limpens et al. [167], Twenhöven [168], Bobbink et al. [169], and Tomassen et al. [172]).
Trees
Tree growth in boreal and temperate wetlands mainly depends on nutrients that are released within peatland soils via mineralization [149,171] that is actively stimulated by the effects of trees on water table height and nutrient cycling. Specifically, trees stimulating water table drawdown increase the amount of nutrients released through aerobic decomposition [140]. When atmospheric deposition increases the input of nutrients, these may partly infiltrate into the root zone of trees [153], facilitating the frequently observed invasion of wetlands by trees [172]. In addition, trees may attract and accumulate nutrients from the surrounding environment through advective transport by groundwater, which is driven by the active transpiration of vascular plants [31,148,173,174,175,176]. This latter mechanism, however, is expected to be restricted to climates where annual evapotranspiration approaches precipitation [162,173]. Finally, dead tree stems provide a nutrient-rich environment that is suitable for successful colonization by tree seedlings [177,178].
2.2.3. Foundation Species Interactions with Other Environmental Conditions
Algae
Apart from the changes in evapotranspiration and precipitation previously mentioned, peatlands within the temperate and boreal latitudes of the northern hemisphere are also being exposed to the strongest increases in temperature due to climate change [179]. Potential effects of these climatic changes include algae and other primary producers becoming more frequently exposed to water scarcity stress during droughts, which will likely affect their community structure. For example, in an Alaskan fen, an experimental drought event triggered nutrient concentration increases and consequent community shifts from coccoid (edible) to filamentous (inedible) growth forms, as grazers (i.e., heterotrophic bacteria and macroinvertebrates) started feeding selectively on the most edible algae in dry soil [180]. In boreal wetlands, epiphytic nitrogen-fixing cyanobacteria, such as Nostoc, Calothrix, and Stigonema are often associated the red-stemmed feathermoss Pleurozium schreberi and other mosses [181], thus contributing to nutrient cycling and driving nutrient availability [182]. Therefore, these cyanobacteria can be seen as secondary foundation species [183,184] that grow on moss (primary foundation species) and contribute to nutrient cycling and other ecosystem functions in wetlands.
Vascular and Nonvascular Plants
Sphagnum mosses are an important source of acidity in boreal wetlands [185,186], most likely because they produce organic acids through the humification of dead tissue [186,187]. As a result, Sphagnum mosses can substantially reduce local pH [188], which may inhibit vascular plant growth [31,146]. The thermal properties of Sphagnum tissue can modify growing season length in a way that changes with depth [31]. Specifically, the living tissue of Sphagnum occurring at peatlands’ top layers (<5 cm) has insulating properties that lengthen the growing season of Sphagnum itself [146,189]. By contrast, Sphagnum peat conducts heat poorly, shortening the growing season of vascular plants that are rooted in Sphagnum-dominated peat [156,190,191]. Forested boreal wetlands receiving lower precipitation are more vulnerable to disturbance by fire than wetlands receiving more rainfall, where the relatively high water content of Sphagnum mosses reduces the likelihood of fire propagation or the combustion depth of fires [154,192]. A positive feedback mechanism may be involved, as the Sphagnum species that create the most fire-resistant patches struggle to regenerate after a fire [193].
Trees
As noted in Section 2.2.2, fires may propagate into boreal and temperate wetlands, combusting aboveground biomass and, depending on water table height, burn upper peat layers [194]. Peat that is not water logged will more easily burn. Therefore, in upland forests with lower water tables, fires are more frequent and fire return intervals are much shorter than in wetlands with few trees and a higher water table. Whereas typical fire return intervals in upland forests are in the order of a century, fire return intervals in wetlands with trees may be longer than 400 years [195,196]. Yet, tree species such as Picea mariana may create conditions leading to fire propagation by providing flammable material and lowering the water table by increasing interception and evapotranspiration, especially during extended periods of drought [193,197].
3. Discussion
3.1. The Impacts of Foundation Species on Wetland Ecosystem Structure and Functions
In this review, we provided an overview on which foundation species define the structural features and ecosystem functioning of representative freshwater wetlands and how they influence and respond to hydrology, nutrient and habitat dynamics (Table 1). In addition, we explored how the relative dominance of foundation species changes across key wetland environmental gradients (Figure 2, Figure 3, Figure 5 and Figure 6).

Table 1.
Foundation species analyzed and their respective roles in controlling ecosystem functions in freshwater wetlands. Direct roles are indicated by ‘x’, while indirect ones are indicated by ‘(x)’—for example, some foundation species lower the water table, which increases the rate of aerobic decomposition, in turn accelerating nutrient cycling and thus increasing nutrient availability.
Foundation species typically have cascading impacts at different spatial scales, from small-scale microscopic algae forming cohesive benthic mats that retain water and nutrients to trees that create extensive above- and belowground habitats to numerous other organisms, including secondary foundation species that further support biodiversity, such as vascular epiphytes [183,184]. These foundation species may build and maintain self-sustaining systems as they facilitate themselves to maintain their dominance on local scales, for example in moss-dominated bogs [198], tree islands [199], and periphyton mats [200]. However, these species can also compete over larger scales for key resources, causing both spatial heterogeneity and patchiness in patterned wetlands [199,201]. Therefore, foundation species need to be better documented and conserved in freshwater wetlands as well as in other ecosystems.
3.2. Foundation Species as Targets of Conservation
The ecosystem-scale role that foundation species play and their relevance to conservation have been only recently recognized and studied in-depth [1,202,203]. By contrast, nature conservation efforts often target species that strongly regulate community structure and ecosystem functions (keystone species), inspire public support for conservation (flagship species), or serve as indicators for ecosystem conditions (umbrella species) [204,205]. In light of this historic focus, we argue that foundation species deserve more scientific and conservation attention because they support ecosystem functioning as well as flagship, keystone, and umbrella species [3]. Thus, the conservation of foundation species may enable the conservation of entire food webs and/or the restoration of wetland ecosystems. For example, conserving sawgrass and dominant benthic algae can help maintain oligotrophic marshes in the Everglades [40,176] and elsewhere: planting salt-tolerant bald cypress trees can help restore degraded coastal wetlands [124]; introducing submerged aquatic macrophytes can stabilize sediment, reduce the light available to bloom-forming algae, and provide refuge to fish species that eat algae [206]; preserving microphytic crusts (algae, cyanobacteria, and fungi) can reduce water scarcity as these organisms retain water [207]; and conserving Sphagnum species can ensure the carbon-sequestering function of boreal peatlands [208].
Conservation actions need to take into account that biological communities depending on foundation species may be particularly sensitive to ecosystem shifts between alternative stable states [209]. For example, because of long-term nutrient enrichment (a press driver) caused by decades of agricultural fertilization [94,97,210,211,212,213], cattail has expanded replacing thousands of hectares of sawgrass marshes and aquatic slough areas in the central Everglades [50], in marshes in northern Belize [47], and wetlands in Guadeloupe (Lesser Antilles [62]). Similarly, decades of elevated atmospheric nitrogen deposition in northern temperate wetlands may have induced a transition towards ecosystem states that will eventually be dominated by currently invading vascular plants such as Molinia and Betula spp. [31,172,175]. To best protect native pristine species communities and ecosystem functioning in wetlands subject to switches between alternative stable states, the environmental conditions supporting native species need to be rehabilitated. For instance, in the Everglades, the restoration of low surface water P concentrations, greater water flow, and natural marsh vegetation (i.e., a more desirable stable state than eutrophic waters with high cattail dominance) is the major goal of ongoing management efforts. The recognition of the roles of all foundation species, including undesirable ones, such as cattail, can ultimately facilitate ecosystem conservation. Given the critical role of foundation species for wetland ecosystem structure and functioning, their recovery from the impacts of eutrophication, drought or saltwater intrusion can lead to positive cascading effects on wetlands undergoing restoration or subject to mitigated environmental impacts [214,215].
3.3. Future Research Directions
We provide an overview of foundation species distribution, roles, and relative dominance in freshwater wetlands that can inform their conservation and that of the species that rely on them (e.g., wading birds), and of the key ecosystem functions that foundation species contribute to. For example, we have shown how the flow of clean freshwater is essential to maintaining high dominance of foundation species from benthic algae to marsh plant species in various subtropical wetlands. Without such clean water flow, cattail and other plant and algal species adapted to high nutrient levels can take over primary production and transform ecosystem structure and functioning towards monoculture and loose benthic mats with generalist algal species. To support effective wetland conservation and restoration, it is imperative to identify which environmental drivers affect foundation species and to understand the mechanisms by which they do so through empirical, experimental and/or modeling and forecasting studies.
Complementary modeling approaches can help identify the key environmental drivers of the current distribution and abundance of foundation species and to understand how drivers and distributions are changing as result of global trends, such as climatic changes and increasing atmospheric nutrient deposition levels [31,162,216,217]. To understand and predict the changes in species distribution and community structure and function in response to changes in environmental drivers, researchers have used a suite of modeling approaches. For instance, individual-based gap models can track individual tree growth based on biotic (e.g., competition and species-specific life history traits) and abiotic (e.g., resource availability) factors, thus helping better understand how vegetation shifts from freshwater marsh species to mangroves [218], or from Sphagnum-dominated to vascular plant-dominated boreal wetlands [31]. The Everglades Landscape Vegetation Succession (ELVeS) model [219] informs Everglades adaptive management (i.e., management that takes new knowledge into account and addresses unforeseen issues) through predictions of how hydrologic conditions and limiting nutrient levels affect the distribution of foundation species (e.g., sawgrass) and how they are going to adapt to environmental changes. Simulation models can be used to select performance measures in ecosystem restoration efforts by linking ecosystem attributes to specific mechanisms of system behavior [220]. Importantly, modeling and synthesis efforts need to address the interconnected effects of multiple press and pulse drivers on hydrology, nutrients and other environmental conditions and thus on foundation species in freshwater wetlands.
The key outstanding research need is to forecast the distribution and dominance patterns of foundation species along climate change, wetland restoration, and ecosystem development trajectories (e.g., community structure, hydrology, demography, and nutrient dynamics) [221]. We recommend that new research focuses on (i) predicting how desired foundation species may decline within their current habitat or how undesired foundation species may expand into new geographical areas, (ii) identifying conditions where a foundation species could be introduced into a degraded habitat to build ecosystem resilience, and (iii) projecting the impact of future environmental conditions on the dynamics of foundation species and assessing how these changes may cascade through the ecosystem and the food web toward regime shifts. Importantly, experiments to determine foundation species’ environmental thresholds in terms of, for example, water and nutrient availability, can inform models on the impacts of press drivers, such as climate change and eutrophication, on these species. We hope that this paper will inspire new research and encourage a broader adoption of the concept of foundation species into the science, management and conservation of freshwater wetlands around the world.
Supplementary Materials
The following is available online at https://www.mdpi.com/2073-4441/11/2/265/s1, Table S1: Publications used for the literature review (references only cited in the introduction and discussion and/or used only for theoretical context are excluded; studies with useful findings, but not specific to subtropical or boreal/temperate wetlands, are left unticked in the 3rd and 4th columns) (Section 2).
Author Contributions
L.M., E.E.G., E.C.-M., and J.P.S. conceived the manuscript idea and developed the review structure in collaboration with L.Z., C.A., and M.B.E.; L.M., M.B.E., J.P.S., and L.Z. conducted the literature review and wrote the manuscript with regular input, feedback, and advice from E.E.G., C.A. and E.C.-M.
Funding
This research received no external funding. The APC was funded by Florida International University and University of Florida.
Acknowledgments
This research was conducted with the support of the National Science Foundation through the Florida Coastal Everglades Long-Term Ecological Research program under Cooperative Agreements #DEB-1237517, #DBI-0620409, and #DEB-9910514. This is contribution number 896 from the Southeast Environmental Research Center in the Institute of Water & Environment at Florida International University.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Angelini, C.; Altieri, A.H.; Silliman, B.R.; Bertness, M.D. Interactions among foundation species and their consequences for community organization, biodiversity, and conservation. BioScience 2011, 61, 782–789. [Google Scholar] [CrossRef]
- Dayton, P.K. Toward an understanding of community resilience and the potential effects of enrichments to the benthos at McMurdo Sound, Antarctica. In Proceedings of the Colloquium on Conservation Problems in Antarctica; Parker, B.C., Ed.; Allen Press: Dublin, Ireland, 1972; pp. 81–95. [Google Scholar]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- Faunce, C.H.; Serafy, J.E. Selective use of mangrove shorelines by snappers, grunts and great barracuda. Mar. Ecol. Prog. Ser. 2008, 356, 153–162. [Google Scholar] [CrossRef]
- Gaiser, E.E.; McCormick, P.V.; Hagerthey, S.E.; Gottlieb, A.D. Landscape patterns of periphyton in the Florida Everglades. Crit. Rev. Environ. Sci. Technol. 2011, 41, 92–120. [Google Scholar] [CrossRef]
- Parker, G.G.; Harmon, M.A.; Lefsky, J.Q. Three-dimensional structure of an old-growth Pseudotsuga tsuga canopy and its implications for radiation balance, microclimate, and gas exchange. Ecosystems 2004, 7, 440–453. [Google Scholar] [CrossRef]
- Stachowicz, J.J. Mutualism, facilitation, and the structure of ecological communities. BioScience 2001, 51, 235–246. [Google Scholar] [CrossRef]
- Bruno, J.F.; Bertness, M.D. Habitat modification and facilitation in benthic marine communities. In Marine Community Ecology; Bertness, M.D., Gaines, S.D., Hay, M.E., Eds.; Sinauer: Sunderland, MA, USA, 2001; pp. 201–218. [Google Scholar]
- Gedan, K.B.; Bertness, M.D. How will warming affect the salt marsh foundation species Spartina patens and its ecological role? Oecologia 2010, 164, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Osland, M.J.; Enwright, N.; Day, R.H.; Doyle, T.W. Winter climate change and coastal wetland foundation species: Salt marshes vs. mangrove forests in the southeastern United States. Glob. Chang. Biol. 2013, 19, 1482–1494. [Google Scholar] [CrossRef]
- Thompson, D.K.; Baisley, A.S.; Waddington, J.M. Seasonal variation in albedo and radiation exchange between a burned and unburned forested peatland: Implications for peatland evaporation. Hydrol. Process. 2015, 29, 3227–3235. [Google Scholar] [CrossRef]
- Sharitz, R.; Mitsch, W.J. Southern floodplain forests. In Biodiversity of the Southeastern United States: Lowland Terrestrial Communities; Echternacht, A.C., Ed.; John Wiley and Sons: New York, NY, USA, 1993. [Google Scholar]
- Kominoski, J.S.; Gaiser, E.E.; Baer, S.G. Advancing Theories of Ecosystem Development through Long-Term Ecological Research. BioScience 2018, 68, 554–562. [Google Scholar] [CrossRef]
- Junk, W.J.; An, S.; Finlayson, C.M.; Gopal, B.; Květ, J.; Mitchell, S.A.; Mitsch, W.J.; Robarts, R.D. Current state of knowledge regarding the world’s wetlands and their future under global climate change: A synthesis. Aquat. Sci. 2013, 75, 151–167. [Google Scholar] [CrossRef]
- Scrosati, R.A. Community-level facilitation by macroalgal foundation species peaks at an intermediate level of environmental stress. Algae 2017, 32, 41–46. [Google Scholar] [CrossRef]
- Smith, T.J.; Foster, A.M.; Tiling-Range, G.; Jones, J.B., Jr. Dynamics of mangrove-marsh ecotones in subtropical coastal wetlands: Fire, sea-level rise, and water levels. Fire Ecol. 2013, 9, 66–77. [Google Scholar] [CrossRef]
- Barr, J.G.; Engel, V.; Smith, T.J.; Fuentes, J.D. Hurricane disturbance and recovery of energy balance, CO2 fluxes and canopy structure in a mangrove forest of the Florida Everglades. Agric. For. Meteorol. 2012, 153, 54–66. [Google Scholar] [CrossRef]
- Davis, S.E.; Boucek, R.; Castañeda-Moya, E.; Dessu, S.; Gaiser, E.; Kominoski, J.; Sah, J.P.; Surratt, D.; Troxler, T. Episodic disturbances drive nutrient dynamics along freshwater-to-estuary gradients in a subtropical wetland. Ecosphere 2018, 9, e02296. [Google Scholar] [CrossRef]
- Bertness, M.D.; Callaway, R. Positive interactions in communities. Trends Ecol. Evol. 1994, 9, 191–193. [Google Scholar] [CrossRef]
- Michalet, R.; Broker, R.W.; Cavieres, L.A.; Kikvidze, Z.; Lortie, C.J.; Pugnaire, F.I.; Valiente-Banuet, A.; Callaway, R.M. Do biotic interactions shape both sides of the humped-back model of species richness in plant communities? Ecol. Lett. 2006, 9, 767–773. [Google Scholar] [CrossRef]
- Holmgren, M.; Scheffer, M. Strong facilitation in mild environments: The stress gradient hypothesis revisited. J. Ecol. 2010, 98, 1269–1275. [Google Scholar] [CrossRef]
- He, Q.; Bertness, M.D.; Altien, A.H. Global shifts towards positive species interactions with increasing environmental stress. Ecol. Lett. 2013, 16, 695–706. [Google Scholar] [CrossRef]
- Passy, S.I. Abundance inequality in freshwater communities has an ecological origin. Am. Nat. 2016, 187, 502–516. [Google Scholar] [CrossRef]
- Kosten, S.; Huszar, V.L.M.; Bécares, E.; Costa, L.S.; van Donk, E.; Hansson, L.A.; Jeppesen, E.; Kruk, C.; Lacerot, G.; Mazzeo, N.; et al. Warmer climates boost cyanobacterial dominance in shallow lakes. Glob. Chang. Biol. 2012, 18, 118–126. [Google Scholar] [CrossRef]
- Newman, S.; Schuette, J.; Grace, J.B.; Rutchey, K.; Fontaine, T.; Reddy, K.R.; Pietrucha, M. Factors influencing cattail abundance in the northern Everglades. Aquat. Bot. 1998, 60, 265–280. [Google Scholar] [CrossRef]
- Crain, C.M.; Bertness, M.D. Community impacts of a tussock sedge: Is ecosystem engineering important in benign habitats? Ecology 2005, 86, 2695–2704. [Google Scholar] [CrossRef]
- Costanza, R.; de Groot, R.; Sutton, P.; Van der Ploeg, S.; Anderson, S.J.; Kubiszewski, I.; Farber, S.; Turner, R.K. Changes in the global value of ecosystem services. Glob. Environ. Chang. 2014, 26, 152–158. [Google Scholar] [CrossRef]
- Davidson, N.C. How much wetland has the world lost? Long-term and recent trends in global wetland area. Mar. Freshw. Res. 2014, 65, 934–941. [Google Scholar] [CrossRef]
- Scheffer, M.; Hosper, S.H.; Meijer, M.L.; Moss, B.; Jeppesen, E. Alternative equilibria in shallow lakes. Trends Ecol. Evol. 1993, 8, 275–279. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gosselink, J.G. Wetlands, 3rd ed.; Wiley: New York, NY, USA, 2000; 920p. [Google Scholar]
- Eppinga, M.B.; Rietkerk, M.; Wassen, M.J.; De Ruiter, P.C. Linking habitat modification to catastrophic shifts and vegetation patterns in bogs. Plant Ecol. 2009, 200, 53–68. [Google Scholar] [CrossRef]
- Larsen, L.G.; Harvey, J.W.; Crimaldi, J.P. A delicate balance: Ecohydrological feedbacks governing landscape morphology in a lotic peatland. Ecol. Monogr. 2007, 77, 591–614. [Google Scholar] [CrossRef]
- Douglas, M.S. The Everglades: River of Grass; Pineapple Press Inc.: Sarasota, FL, USA, 2007. [Google Scholar]
- Lodge, T.E. The Everglades Handbook: Understanding the Ecosystem; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Lee, S.S. Mastogloia calcarea. 2014. Available online: https://diatoms.org/species/mastogloia_calcarea (accessed on 30 October 2018).
- Gaiser, E.E.; La Hée, J.M.L.; Tobias, F.A.; Wachnicka, A.H. Mastogloia smithii var. lacustris Grun.: A structural engineer of calcareous mats in karstic subtropical wetlands. Proc. Acad. Natl. Sci. USA 2010, 160, 99–112. [Google Scholar]
- Gottlieb, A.D.; Richards, J.H.; Gaiser, E.E. Comparative study of periphyton community structure in long and short-hydroperiod Everglades marshes. Hydrobiologia 2006, 569, 195–207. [Google Scholar] [CrossRef]
- La Hée, J.M.; Gaiser, E.E. Benthic diatom assemblages as indicators of water quality in the Everglades and three tropical karstic wetlands. Freshw. Sci. 2012, 31, 205–221. [Google Scholar] [CrossRef]
- Lee, S.S.; Gaiser, E.E.; Trexler, J.C. Diatom-based models for inferring hydrology and periphyton abundance in a subtropical karstic wetland: Implications for ecosystem-scale bioassessment. Wetlands 2013, 33, 157–173. [Google Scholar] [CrossRef]
- Marazzi, L.; Gaiser, E.E.; Tobias, F.A. Phosphorus scarcity and desiccation stress increase the occurrence of dominant taxa in wetland benthic primary producer communities. Aquat. Ecol. 2017, 51, 571–589. [Google Scholar] [CrossRef]
- Van Meter, N.N. Some Quantitative and Qualitative Aspects of Periphyton in the Everglades. Master’s Thesis, University of Miami, Coral Gables, FL, USA, 1965; 98p. [Google Scholar]
- Lee, S.S.; Gaiser, E.E.; Van De Vijver, B.; Edlund, M.B.; Spaulding, S.A. Morphology and typification of Mastogloia smithii and M. lacustris, with descriptions of two new species from the Florida Everglades and the Caribbean region. Diatom Res. 2014, 29, 325–350. [Google Scholar] [CrossRef]
- Hagerthey, S.E.; Bellinger, B.J.; Wheeler, K.; Gantar, M.; Gaiser, E.E. Everglades periphyton: A biogeochemical perspective. Crit. Rev. Environ. Sci. Technol. 2011, 41 (Suppl. 1), 309–343. [Google Scholar] [CrossRef]
- Trexler, J.C.; Loftus, W.F. Invertebrates of the Florida Everglades. In Invertebrates in Freshwater Wetlands: An International Perspective on their Ecology; Batzer, D., Boix, D., Eds.; Springer: New York, NY, USA, 2016; pp. 321–356. [Google Scholar]
- Gaiser, E.E.; Gottlieb, A.D.; Lee, S.S.; Trexler, J.C. The Importance of Species-Based Microbial Assessment of Water Quality in Freshwater Everglades Wetlands. In Microbiology of the Everglades Ecosystem; Entry, J., Jayachandran, K., Gottlieb, A.D., Ogram, A., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 115–130. [Google Scholar]
- Gottlieb, A.; Richards, J.; Gaiser, E. Effects of desiccation duration on the community structure and nutrient retention of short and long-hydroperiod Everglades periphyton mats. Aquat. Bot. 2005, 82, 99–112. [Google Scholar] [CrossRef]
- Rejmánková, E.; Komárková, J. A function of cyanobacterial mats in phosphorus-limited tropical wetlands. Hydrobiologia 2000, 431, 135–153. [Google Scholar] [CrossRef]
- Novelo, E.; Tavera, R. The role of periphyton in the regulation and supply of nutrients in a wetland at El Edén, Quintana Roo. In The Lowland Maya Area: Three Millennia at the Human-Wildland Interface; Food Products Press: New York, NY, USA, 2003; pp. 217–236. [Google Scholar]
- Loveless, C.M. A study of the vegetation in the Florida Everglades. Ecology 1959, 40, 1–9. [Google Scholar] [CrossRef]
- Miao, S.L.; Sklar, F.H. Biomass and nutrient allocation of sawgrass and cattail along a nutrient gradient in the Florida Everglades. Wetl. Ecol. Manag. 1998, 5, 245–263. [Google Scholar] [CrossRef]
- Herndon, A.; Gunderson, L.; Stenberg, J. Sawgrass (Cladium jamaicense) survival in a regime of fire and flooding. Wetlands 1991, 11, 17–28. [Google Scholar] [CrossRef]
- Givnish, T.J.; Volin, J.C.; Owen, V.D.; Volin, V.C.; Muss, J.D.; Glaser, P.H. Vegetation differentiation in the patterned landscape of the central Everglades: Importance of local and landscape drivers. Glob. Ecol. Biogeogr. 2008, 17, 384–402. [Google Scholar] [CrossRef]
- Ross, M.S.; Reed, D.L.; Sah, J.P.; Ruiz, P.L.; Lewin, M.T. Vegetation:environment relationships and water management in Shark Slough, Everglades National Park. Wetl. Ecol. Manag. 2003, 11, 291–303. [Google Scholar] [CrossRef]
- Ross, M.S.; Sah, J.P.; Ruiz, P.L.; Jones, D.T.; Cooley, H.C.; Travieso, R.; Snyder, J.R.; Hagyari, D. Effect of Hydrology Restoration on the Habitat of the Cape Sable Seaside Sparrow; Report to Everglades National Park, SERC Research Reports. 85; South Florida Natural Resource Center: Homestead, FL, USA, 2006; pp. 1–46.
- Todd, M.J.; Muneepeerakul, R.; Pumo, D.; Azaele, S.; Miralles-Wilhelm, F.; Rinaldo, A.; Rodriguez-Iturbe, I. Hydrological drivers of wetland vegetation community distribution within Everglades National Park, Florida. Adv. Water Resour. 2010, 33, 1279–1289. [Google Scholar] [CrossRef]
- Sah, J.P.; Ross, M.S.; Saha, S.; Minchin, P.; Sadle, J. Trajectories of vegetation response to water management in Taylor Slough, Everglades National Park, Florida. Wetlands 2014, 34, 65–79. [Google Scholar] [CrossRef]
- Toth, L.A. Effects of Hydrologic Regimes on Lifetime Production and Nutrient Dynamics of Sawgrass (Technical Publication# 87-6); Environmental Sciences Division: West Palm Beach, FL, USA, 1987. [Google Scholar]
- Busch, D.E.; Loftus, W.F.; Bass, O.L. Long-term hydrologic effects on marsh plant community structure in the southern Everglades. Wetlands 1998, 18, 230–241. [Google Scholar] [CrossRef]
- Armentano, T.V.; Sah, J.P.; Ross, M.S.; Jones, D.T.; Cooley, H.C.; Smith, C.S. Rapid responses of vegetation to hydrological changes in Taylor Slough, Everglades National Park, Florida, USA. Hydrobiologia 2006, 569, 293–309. [Google Scholar] [CrossRef]
- Gunderson, L.H. South florida: The reality of change and the prospects for sustainability: Managing surprising ecosystems in southern Florida. Ecol. Econ. 2001, 37, 371–378. [Google Scholar] [CrossRef]
- Mazzotti, F.J.; Fling, H.E.; Merediz, G.; Lazcano, M.; Lasch, C.; Barnes, T. Conceptual ecological model of the Sian Ka’an Biosphere Reserve, Quintana Roo, Mexico. Wetlands 2005, 25, 980–997. [Google Scholar] [CrossRef]
- Imbert, D.; Delbé, L. Ecology of fire-influenced Cladium jamaicense marshes in Guadeloupe, Lesser Antilles. Wetlands 2006, 26, 289–297. [Google Scholar] [CrossRef]
- Meerman, J.C. Ecological Characterization of the New River Lagoon, Orange Walk District, Belize; Report prepared for the Freshwater Program; Programme for Belize: Belize City, BZ, USA, 2006; pp. 2–26. [Google Scholar]
- Rejmánková, E.; Pope, K.O.; Post, R.; Maltby, E. Herbaceous wetlands of the Yucatan peninsula: Communities at extreme ends of environmental gradients. Int. Rev. Gesamten Hydrobiol. 1996, 81, 223–252. [Google Scholar] [CrossRef]
- Grace, J.B. The effects of nutrient additions on mixtures of Typha latifolia L. and Typha domingensis Pers. along a water depth gradient. Aquat. Bot. 1988, 31, 83–92. [Google Scholar] [CrossRef]
- Sun, G.; McNulty, S.G.; Shepard, J.P.; Amatya, D.M.; Riekerk, H.; Comerford, N.B.; Skaggs, W.; Swift, L., Jr. Effects of timber management on the hydrology of wetland forests in the southern United States. For. Ecol. Manag. 2001, 143, 227–236. [Google Scholar] [CrossRef]
- Feliciano, E.A. Multi-Scale Remote Sensing Assessments of Forested Wetlands: Applications to the Everglades National Park. Open Access Diss. 2015, p. 1417. Available online: https://scholarlyrepository.miami.edu/oa_dissertations/1417 (accessed on 1 September 2018).
- Allen, J.A.; Pezeshki, S.R.; Chambers, J.L. Interaction of flooding and salinity stress on baldcypress (Taxodium distichum). Tree Physiol. 1996, 16, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Megonigal, J.P.; Day, F.P. Effects of flooding on root and shoot production of bald cypress in large experimental enclosures. Ecology 1992, 73, 1182–1193. [Google Scholar] [CrossRef]
- Shanklin, J.; Kozlowski, T.T. Effect of flooding of soil on growth and subsequent responses of Taxodium distichum seedlings to SO2. Environ. Pollut. Ser. A Ecol. Biol. 1985, 38, 199–212. [Google Scholar] [CrossRef]
- Pezeshki, S.R.; Delaune, R.D.; Patrick, W.H., Jr. Gas exchange characteristics of bald cypress (Taxodium distichum L.): Evaluation of responses to leaf aging, flooding, and salinity. Can. J. For. Res. 1986, 16, 1394–1397. [Google Scholar] [CrossRef]
- Pezeshki, S.R. Root responses of flood-tolerant and flood-sensitive tree species to soil redox conditions. Trees 1991, 5, 180–186. [Google Scholar] [CrossRef]
- Pezeshki, S. Response of baldcypress (Taxodium distichum) seedlings to hypoxia: Leaf protein content, ribulose-1,5-bisphosphate carboxy-lase/oxygenase activity and photosynthesis. Photosynthetica 1994, 30, 59–68. [Google Scholar]
- Crawford, R.M.M. Root survival in flooded soils. In Ecosystems of the World; Gore, A.J.P., Ed.; Elsevier Scientific: Amsterdam, The Netherlands, 1983; pp. 257–283. [Google Scholar]
- Hook, D.D. Adaptations to flooding with fresh water. In Flooding and Plant Growth; Kozlowski, T.T., Ed.; Academic Press: Orlando, FL, USA, 1984; pp. 265–294. [Google Scholar]
- Wang, M.; Talbot, J.; Moore, T.R. Drainage and fertilization effects on nutrient availability in an ombrotrophic peatland. Sci. Total Environ. 2017, 121, 1255–1263. [Google Scholar] [CrossRef]
- Elcan, J.M.; Pezeshki, S.R. Effects of Flooding on Susceptibility of Taxodium distichum L. Seedlings to Drought. Photosynthetica 2002, 40, 177–182. [Google Scholar] [CrossRef]
- Lei, T.; Middleton, B.A. Repeated Drought Alters Resistance of Seed Bank Regeneration in Baldcypress Swamps of North America. Ecosystems 2018, 21, 190–201. [Google Scholar] [CrossRef]
- Gaiser, E.E.; Richards, J.H.; Trexler, J.C.; Jones, R.D.; Childers, D.L. Periphyton responses to eutrophication in the Florida Everglades: Cross-system patterns of structural and compositional change. Limnol. Oceanogr. 2006, 51, 617–630. [Google Scholar] [CrossRef]
- Gaiser, E.E. Periphyton as an indicator of restoration in the Florida Everglades. Ecol. Indic. 2009, 9, S37–S45. [Google Scholar] [CrossRef]
- Pinowska, A.; Francoeur, S.N.; Manoylov, K.M. Substratum-associated microbiota. Water Environ. Res. Lit. Rev. 2008, 80, 1858–1891. [Google Scholar] [CrossRef]
- Dodds, W.K. The role of periphyton in phosphorus retention in shallow freshwater aquatic systems. J. Phycol. 2003, 39, 840–849. [Google Scholar] [CrossRef]
- Lamberti, G.A. The role of periphyton in benthic food webs. In Algal Ecology, 1st ed.; Stevenson, R.J., Bothwell, M.L., Lowe, R.L., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 533–564. [Google Scholar]
- Hagerthey, S.E.; Cole, J.J.; Kilbane, D. Aquatic metabolism in the Everglades: Dominance of water column heterotrophy. Limnol. Oceanogr. 2010, 55, 653–666. [Google Scholar] [CrossRef]
- Rejmankova, E.; Pope, K.O.; Pohl, M.D.; Rey-Benayas, J.M. Freshwater wetland plant communities of northern Belize: Implications for paleoecological studies of Maya wetland agriculture. Biotropica 1995, 27, 28–36. [Google Scholar] [CrossRef]
- Gaiser, E.E.; Anderson, E.P.; Castañeda-Moya, E.; Collado-Vides, L.; Fourqurean, J.W.; Heithaus, M.R.; Jaffé, R.; Lagomasino, D.; Oehm, N.J.; Price, R.M.; et al. New perspectives on an iconic landscape from comparative international long-term ecological research. Ecosphere 2015, 6, 1–18. [Google Scholar] [CrossRef]
- Steward, K.K.; Ornes, W.H. The autecology of sawgrass in the Florida Everglades. Ecology 1975, 56, 162–171. [Google Scholar] [CrossRef]
- Steward, K.K.; Ornes, W.H. Mineral nutrition of sawgrass (Cladium jamaicense Crantz) in relation to nutrient supply. Aquat. Bot. 1983, 16, 349–359. [Google Scholar] [CrossRef]
- Davis, S.M. Sawgrass and cattail nutrient flux: Leaf turnover, decomposition, and nutrient flux of sawgrass and cattail in the Everglades. Aquat. Bot. 1991, 40, 203–224. [Google Scholar] [CrossRef]
- Noe, G.B.; Childers, D.L.; Jones, R.D. Phosphorus Biogeochemistry and the Impact of Phosphorus Enrichment: Why Is the Everglades so Unique? Ecosystems 2001, 4, 603–624. [Google Scholar] [CrossRef]
- Richardson, C.J.; Ferrell, G.M.; Vaithiyanathan, P. Nutrient effects on stand structure, resorption efficiency, and secondary compounds in Everglades sawgrass. Ecology 1999, 80, 2182–2192. [Google Scholar] [CrossRef]
- Brix, H.; Lorenzen, B.; Mendelssohn, I.A.; McKee, K.L.; Miao, S.L. Can differences in phosphorus uptake kinetics explain the distribution of cattail and sawgrass in the Florida Everglades? Plant Biol. 2010, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Beare, P.A.; Zedler, J.B. Cattail invasion and persistence in a coastal salt marsh: The role of salinity. Estuaries 1987, 10, 165–170. [Google Scholar] [CrossRef]
- Thomas, S.E.; Gaiser, E.E.; Tobias, F.A.C. Effects of shading on calcareous benthic periphyton in a short-hydroperiod oligotrophic alkaline wetland (Everglades, FL, USA). Hydrobiologia 2006, 569, 209–221. [Google Scholar] [CrossRef]
- Grimshaw, H.J.; Rosen, M.; Swift, D.R.; Rodberg, K.; Noel, J.M. Marsh phosphorus concentrations, phosphorus content and species composition of Everglades periphyton communities. Arch. Hydrobiol. 1993, 139, 17–27. [Google Scholar]
- Chapin, F.S., III. The mineral nutrition of wild plants. Ann. Rev. Ecol. System. 1980, 11, 233–260. [Google Scholar] [CrossRef]
- Newman, S.; Grace, J.B.; Koebel, J.W. Effects of nutrients and hydroperiod on Typha, Cladium, and Eleocharis: Implications for Everglades restoration. Ecol. Appl. 1996, 6, 774–783. [Google Scholar] [CrossRef]
- Davis, S.M.; Ogden, J.C. Towards ecosystem restoration. In Everglades, The Ecosystem and Its Restoration; Davis, S.M., Ogden, J.C., Eds.; St. Lucie Press: Delray Beach, FL, USA, 1994; pp. 769–796. [Google Scholar]
- Hagerthey, S.E.; Newman, S.; Rutchey, K.; Smith, E.P.; Godin, J. Multiple regime shifts in a subtropical peatland: Community-specific thresholds to eutrophication. Ecol. Monogr. 2008, 78, 547–565. [Google Scholar] [CrossRef]
- Effler, R.S.; Goyer, R.A.; Lenhard, G.J. Baldcypress and water tupelo responses to insect defoliation and nutrient augmentation in Maurepas Swamp, Louisiana, USA. For. Ecol. Manag. 2006, 236, 295–304. [Google Scholar] [CrossRef]
- Krauss, K.W.; Duberstein, J.A.; Doyle, T.W.; Conner, W.H.; Day, R.H.; Inabinette, L.W.; Whitbeck, J.L. Site condition, structure, and growth of baldcypress along tidal/non-tidal salinity gradients. Wetlands 2009, 29, 505–519. [Google Scholar] [CrossRef]
- Nyman, J.A.; Lindau, C.W. Nutrient availability and flooding stress interact to affect growth and mercury concentration in Taxodium distichum (L.) Rich. seedlings. Environ. Exp. Bot. 2016, 125, 77–86. [Google Scholar] [CrossRef]
- Dickson, R.E.; Broyer, T.C. Effects of aeration, water supply, and nitrogen source on growth and development of tupelo gum and bald cypress. Ecology 1972, 53, 626–634. [Google Scholar] [CrossRef]
- Dickson, R.E.; Broyer, T.C.; Johnson, C.M. Nutrient uptake by tupelo gum and bald cypress from saturated or unsaturated soil. Plant Soil 1972, 37, 297–308. [Google Scholar] [CrossRef]
- Effler, R.S.; Goyer, R.A. Baldcypress and water tupelo sapling response to multiple stress agents and reforestation implications for Louisiana swamps. For. Ecol. Manag. 2006, 226, 330–340. [Google Scholar] [CrossRef]
- Day, R.H.; Doyle, T.W.; Draugelis-Dale, R.O. Interactive effects of substrate, hydroperiod, and nutrients on seedling growth of Salix nigra and Taxodium distichum. Environ. Exp. Bot. 2006, 55, 163–174. [Google Scholar] [CrossRef]
- Hesse, I.D.; Day, J.W.; Doyle, T.W. Long-Term Growth Enhancement of Baldcypress (Taxodium distichum) from Municipal Wastewater Application. Environ. Manag. 1998, 22, 119–127. [Google Scholar] [CrossRef]
- Day, J.W., Jr.; Westphal, A.; Pratt, R.; Hyfield, E.; Rybczyk, J.; Kemp, G.P.; Day, J.N.; Marx, B. Effects of long-term municipal effluent discharge on the nutrient dynamics, productivity, and benthic community structure of a tidal freshwater forested wetland in Louisiana. Ecol. Eng. 2006, 27, 242–257. [Google Scholar] [CrossRef]
- Keim, R.F.; Izdepski, C.W.; Day, J.W. Growth Responses of Baldcypress to Wastewater Nutrient Additions and Changing Hydrologic Regime. Wetlands 2012, 32, 95–103. [Google Scholar] [CrossRef]
- Mazzei, V.; Gaiser, E. Scale and spatial consistency of specialization in an endemic and abundant freshwater diatom from the Caribbean Basin. Freshw. Sci. 2017, 36, 542–554. [Google Scholar] [CrossRef]
- Mazzei, V.; Gaiser, E.E.; Kominoski, J.S.; Troxler, T.; Wilson, B.; Servais, S.; Bauman, L.; Davis, S.; Kelly, S.; Sklar, F.; et al. Functional and compositional responses of periphyton mats to simulated saltwater intrusion in the southern Everglades. Estuaries Coasts 2018, 41, 2105–2119. [Google Scholar] [CrossRef]
- Obeysekera, J.; Barnes, J.; Nungesser, M. Climate sensitivity runs and regional hydrologic modeling for predicting the response of the greater Florida Everglades ecosystem to climate change. Environ. Manag. 2015, 55, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Sah, J.P.; Ross, M.S.; Snyder, J.R.; Stoffella, S.; Blanco, J.; Freixa, J. Re-Sampling of Vegetation Survey Sites within Cape Sable Seaside Sparrow Habitat; South Florida Natural Resources Center, Everglades & Dry Tortugas National Parks: Homestead, FL, USA, 2015; p. 53.
- Alexander, T.R. Sawgrass biology related to the future of the Everglades ecosystem. Proc. Soil Crop Sci. Soc. Fla. 1971, 31, 72–74. [Google Scholar]
- Doyle, T.W.; Krauss, K.W.; Conner, W.H. Predicting the retreat and migration of tidal forests along the northern Gulf of Mexico under sea-level rise. For. Ecol. Manag. 2010, 259, 770–777. [Google Scholar] [CrossRef]
- Ross, M.S.; Meeder, J.F.; Sah, J.P.; Ruiz, P.L.; Telesnicki, G.J. The southeast saline Everglades revisited: 50 years of coastal vegetation change. J. Veg. Sci. 2000, 11, 101–112. [Google Scholar] [CrossRef]
- Furtado, J.I.; Verghese, S.; Liew, K.S.; Lee, T.H. Litter production in a freshwater swamp forest Tasek Bera, Malaysia. In Tropical Ecology and Development, Proceedings of the 5th International Symposium of Tropical Ecology, Kuala Lumpur, Malaysia, 16–21 April 1979; Furtado, J.I., Ed.; International Society of Tropical Ecology: Kuala Lumpur, Malaysia; pp. 815–822.
- Alvarez-López, M. Ecology of Pterocarpus officinalis forested wetlands in Puerto Rico. In Forested Wetlands; Elsevier: Amsterdam, The Netherlands, 1990; pp. 251–265. [Google Scholar]
- Frangi, J.L.; Lugo, A.E. Ecosystem dynamics of a subtropical floodplain forest. Ecol. Monogr. 1985, 55, 351–369. [Google Scholar] [CrossRef]
- Lugo, A.E.; Brown, S.; Brinson, M.M. Forested wetlands in freshwater and salt-water environments. Limnol. Oceanogr. 1988, 33, 894–909. [Google Scholar]
- Liu, X.; Conner, W.H.; Song, B.; Jayakaran, A.D. Forest composition and growth in a freshwater forested wetland community across a salinity gradient in South Carolina, USA. For. Ecol. Manag. 2017, 389, 211–219. [Google Scholar] [CrossRef]
- Pierfelice, K.N.; Lockaby, B.G.; Krauss, K.W.; Conner, W.H.; Noe, G.B.; Ricker, M.C. Salinity Influences on Aboveground and Belowground Net Primary Productivity in Tidal Wetlands. J. Hydrol. Eng. 2017, 22, D5015002. [Google Scholar] [CrossRef]
- Thomas, B.L.; Doyle, T.; Krauss, K. Annual Growth Patterns of Baldcypress (Taxodium distichum) Along Salinity Gradients. Wetlands 2015, 35, 831–839. [Google Scholar] [CrossRef]
- Krauss, K.W.; Chambers, J.L.; Creech, D. Selection for salt tolerance in tidal freshwater swamp species: Advances using baldcypress as a model for restoration. In Ecology of Tidal Freshwater Forested Wetlands of the Southeastern United States; Conner, W.H., Doyle, T.W., Krauss, K.W., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 385–410. [Google Scholar]
- Allen, J.A.; Chambers, J.L.; Pezeshki, S.R. Effects of salinity on baldcypress seedlings: Physiological responses and their relation to salinity tolerance. Wetlands 1997, 17, 310–320. [Google Scholar] [CrossRef]
- Duever, M.J.; Meeder, J.F.; Duever, L.C. Ecosystems of the Big Cypress Swamp. In Cypress Swamps; Ewel, K.C., Odum, H.T., Eds.; University Presses of Florida: Gainesville, FL, USA, 1984; pp. 294–303. [Google Scholar]
- Krauss, K.W.; Duberstein, J.A. Sapflow and water use of freshwater wetland trees exposed to saltwater incursion in a tidally influenced South Carolina watershed. Can. J. For. Res. 2010, 40, 525–535. [Google Scholar] [CrossRef]
- Fry, B.; Bern, A.L.; Ross, M.S.; Meeder, J.F. δ15N Studies of Nitrogen Use by the Red Mangrove, Rhizophora mangle L. in South Florida. Estuar. Coast. Shelf Sci. 2000, 50, 291–296. [Google Scholar] [CrossRef]
- McKee, K.L.; Feller, I.C.; Popp, M.; Wanek, W. Mangrove isotopic (δ15N and δ13C) fractionation across a nitrogen vs. phosphorus limitation gradient. Ecology 2002, 83, 1065–1075. [Google Scholar] [CrossRef]
- McWilliams, W.H.; Rosson, J.F., Jr. Composition and vulnerability of bottomland hardwood forests of the coastal plain province in the south central United States. For. Ecol. Manag. 1990, 33/34, 485–501. [Google Scholar] [CrossRef]
- Zhai, L.; Krauss, K.W.; Liu, X.; Duberstein, J.A.; Conner, W.H.; DeAngelis, D.L.; Sternberg, L.d.S.L. Growth stress response to sea level rise in species with contrasting functional traits: A case study in tidal freshwater forested wetlands. Environ. Exp. Bot. 2018, 155, 378–386. [Google Scholar] [CrossRef]
- Krauss, K.W.; Duberstein, J.A.; Conner, W.H. Assessing stand water use in four coastal wetland forests using sapflow techniques: Annual estimates, errors and associated uncertainties. Hydrol. Process. 2015, 29, 112–127. [Google Scholar] [CrossRef]
- Kemp, P.G.; Conner, W.H.; Day, J.W., Jr. Effects of flooding on decomposition and nutrient cycling in a Louisiana swamp forest. Wetlands 1985, 5, 35–50. [Google Scholar] [CrossRef]
- Schumann, M.; Joosten, H. Global Peatland Restoration Manual. Institute of Botany and Landscape Ecology; Greifswald University: Greifswald, Germany, 2008; p. 103. [Google Scholar]
- Potapova, M. Navicymbula pusilla. 2011. Available online: https://diatoms.org/species/navicymbula_pusilla (accessed on 30 October 2018).
- Cooper, M.J.; Costello, G.M.; Francoeur, S.N.; Lamberti, G.A. Nitrogen limitation of algal biofilms in coastal wetlands of Lakes Michigan and Huron. Freshw. Sci. 2016, 35, 25–40. [Google Scholar] [CrossRef]
- Timoney, K.; Zoltai, S.C.; Goldsborough, L.G. Boreal diatom ponds: A rare wetland associated with nesting whooping cranes. Wetlands 1997, 17, 539–551. [Google Scholar] [CrossRef]
- Johnson, R.E.; Tuchman, N.C.; Peterson, C.G. Changes in the vertical microdistribution of diatoms within a developing periphyton mat. J. N. Am. Benthol. Soc. 1997, 16, 503–519. [Google Scholar] [CrossRef]
- Kilroy, C.; Larned, S.T.; Biggs, B.J.F. The non-indigenous diatom Didymosphenia geminata alters benthic communities in New Zealand rivers. Freshw. Biol. 2009, 54, 1990–2002. [Google Scholar] [CrossRef]
- Belyea, L.R.; Clymo, R.S. Feedback control of the rate of peat formation. Proc. R. Soc. B 2001, 268, 1315–1321. [Google Scholar] [CrossRef] [PubMed]
- Barkman, J.J. Plant communities and synecology of bogs and heath pools in the Netherlands. In Fens and Bogs in the Netherlands: Vegetation, History, Nutrient Dynamics and Conservation; Verhoeven, J.T.A., Ed.; Springer: Dordrecht, The Netherlands, 1992; pp. 173–236. [Google Scholar]
- Robroek, B.J.M.; Limpens, J.; Breeuwer, A.; van Ruijven, J.; Schouten, M.G.C. Precipitation determines the persistence of hollow Sphagnum species on hummocks. Wetlands 2007, 27, 976–986. [Google Scholar] [CrossRef]
- Hájek, T.; Beckett, R.P. Effect of water content components on desiccation and recovery in Sphagnum mosses. Ann. Bot. 2007, 101, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Vitt, D.H. Functional Characteristics and Indicators of Boreal Peatlands. In Boreal Peatland Ecosystems; Wieder, R.K., Vitt, D.H., Eds.; Ecological Studies (Analysis and Synthesis); Springer: Berlin/Heidelberg, Germany, 2006; Volume 188. [Google Scholar]
- Bengtsson, F.; Granath, G.; Rydin, H. Photosynthesis, growth, and decay traits in Sphagnum—A multispecies comparison. Ecol. Evol. 2016, 6, 3325–3341. [Google Scholar] [CrossRef]
- Van Breemen, N. How Sphagnum bogs down other plants. Trends Ecol. Evol. 1995, 10, 270–275. [Google Scholar] [CrossRef]
- Więcek, M.; Martin, P.; Gąbka, M. Distribution patterns and environmental correlates of water mites (Hydrachnidia, Acari) in peatland microhabitats. Exp. Appl. Acarol. 2013, 61, 147–160. [Google Scholar] [CrossRef]
- Eppinga, M.B.; Rietkerk, M.; Borren, W.; Lapshina, E.D.; Bleuten, W.; Wassen, M.J. Regular surface patterning of peatlands: Confronting theory with field data. Ecosystems 2008, 11, 520–536. [Google Scholar] [CrossRef]
- Malmer, N.; Albinsson, C.; Svensson, B.M.; Wallén, B. Interferences between Sphagnum and vascular plants: Effects on plant community structure and peat formation. Oikos 2003, 100, 469–482. [Google Scholar] [CrossRef]
- Frankl, R.; Schmeidl, H. Vegetation change in a South German raised bog: Ecosystem engineering by plant species, vegetation switch or ecosystem level feedback mechanisms? Flora 2000, 195, 267–276. [Google Scholar] [CrossRef]
- Hirano, T.; Suzuki, K.; Hirata, R. Energy balance and evapotranspiration changes in a larch forest caused by severe disturbance during an early secondary succession. Agric. For. Meteorol. 2017, 232, 457–468. [Google Scholar] [CrossRef]
- Takagi, K.; Tsuboya, T.; Takahashi, H.; Inoue, T. Effect of the invasion of vascular plants on heat and water balance in the Sarobetsu mire, northern Japan. Wetlands 1999, 19, 246–254. [Google Scholar] [CrossRef]
- Lamers, L.P.M.; Bobbink, R.; Roelofs, J.G.M. Natural nitrogen filter fails in polluted raised bogs. Glob. Chang. Biol. 2007, 6, 583–586. [Google Scholar] [CrossRef]
- Fay, E.; Lavoie, C. The impact of birch seedlings on evapotranspiration from a mined peatland: An experimental study in southern Quebec, Canada. Mires Peat 2009, 5, 3. [Google Scholar]
- Ohlson, M.; Økland, R.H.; Nordbakken, J.-F.; Dahlberg, B. Fatal interactions between Scots pine and Sphagnum mosses in bog ecosystems. Oikos 2001, 94, 425–432. [Google Scholar] [CrossRef]
- Gunnarsson, U.; Rydin, H. Demography and recruitment of Scots pine on raised bogs in eastern Sweden and relationships to microhabitat differentiation. Wetlands 1998, 18, 133–141. [Google Scholar] [CrossRef]
- Iremonger, S.F.; Kelly, D.L. The responses of four Irish wetland tree species to raised soil water levels. New Phytol. 1988, 109, 491–497. [Google Scholar] [CrossRef]
- Rutter, A.J. Studies in the water relations of Pinus sylvestris in plantation conditions. I. Measurements of rainfall and interception. J. Ecol. 1963, 51, 191–203. [Google Scholar] [CrossRef]
- Limpens, J.; Berendse, F.; Klees, H. How phosphorus availability affects the impact of nitrogen deposition on Sphagnum and vascular plants in bogs. Ecosystems 2004, 7, 793–804. [Google Scholar] [CrossRef]
- Kokfelt, U.; Struyf, E.; Randsalu, L. Diatoms in peat–Dominant producers in a changing environment? Soil Biol. Biochem. 2009, 41, 1764–1766. [Google Scholar] [CrossRef]
- Rober, A.R.; Stevenson, R.J.; Wyatt, K.H. The role of light availability and herbivory on algal responses to nutrient enrichment in a riparian wetland, Alaska. J. Phycol. 2015, 51, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Eppinga, M.B.; Rietkerk, M.; Belyea, L.R.; Nilsson, M.B.; Ruiter, P.C.D.; Wassen, M.J. Resource contrast in patterned peatlands increases along a climatic gradient. Ecology 2010, 91, 2344–2355. [Google Scholar] [CrossRef] [PubMed]
- Caruso, B.S. Temporal and spatial patterns of extreme low flows and effects on stream ecosystems in Otago, New Zealand. J. Hydrol. 2002, 257, 115–133. [Google Scholar] [CrossRef]
- Mosley, L.M. Drought impacts on the water quality of freshwater systems; review and integration. Earth-Sci. Rev. 2015, 140, 203–214. [Google Scholar] [CrossRef]
- Pastor, J.; Peckham, B.; Bridgham, S.; Weltzin, J.; Chen, J. Plant community dynamics, nutrient cycling, and alternative stable equilibria in peatlands. Am. Nat. 2002, 160, 553–568. [Google Scholar] [CrossRef] [PubMed]
- Heijmans, M.M.; Klees, H.; Berendse, F. Competition between Sphagnum magellanicum and Eriophorum angustifolium as affected by raised CO2 and increased N deposition. Oikos 2002, 97, 415–425. [Google Scholar] [CrossRef]
- Limpens, J.; Bohlin, E.; Nilsson, M.B. Phylogenetic or environmental control on the elemental and organo-chemical composition of Sphagnum mosses? Plant Soil 2017, 417, 69–85. [Google Scholar] [CrossRef]
- Twenhöven, F.L. Competition between two Sphagnum species under different deposition levels. J. Bryol. 1992, 17, 71–80. [Google Scholar] [CrossRef]
- Bobbink, R.; Heil, G.W.; Raessen, M.B.A.G. Atmospheric deposition and canopy exchange processes in heathland ecosystems. Environ. Pollut. 1992, 75, 29–37. [Google Scholar] [CrossRef]
- Heil, G.W.; Werger, M.J.A.; De Mol, W.; Van Dam, D.; Heijne, B. Capture of atmospheric ammonium by grassland canopies. Science 1988, 239, 764–765. [Google Scholar] [CrossRef] [PubMed]
- Malmer, N.; Svensson, B.M.; Wallén, B. Interactions between Sphagnum mosses and field layer vascular plants in the development of peat-forming systems. Folia Geobot. Phytotaxon. 1994, 29, 483–496. [Google Scholar] [CrossRef]
- Tomassen, H.B.M.; Smolders, A.J.P.; Lamers, L.P.M.; Roelofs, J.G.M. Stimulated growth of Betula pubescens and Molinia caerulea on ombrotrophic bogs: Role of high levels of atmospheric nitrogen deposition. J. Ecol. 2003, 91, 357–370. [Google Scholar] [CrossRef]
- Eppinga, M.B.; De Ruiter, P.C.; Wassen, M.J.; Rietkerk, M. Nutrients and hydrology indicate the driving mechanisms of peatland surface patterning. Am. Nat. 2009, 173, 803–818. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Mineral Nutrition of Higher Plants; Academic Press: London, UK, 1995. [Google Scholar]
- Rietkerk, M.; Dekker, S.C.; De Ruiter, P.C.; van de Koppel, J. Self-organized patchiness and catastrophic shifts in ecosystems. Science 2004, 305, 1926–1929. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, P.R.; Van Der Valk, A.G.; Newman, S.; Gawlik, D.E.; Troxler Gann, T.; Coronado-Molina, C.A.; Childers, D.L.; Sklar, F.H. Maintaining tree islands in the Florida Everglades: Nutrient redistribution is the key. Front. Ecol. Environ. 2005, 3, 370–376. [Google Scholar] [CrossRef]
- Agnew, A.D.; Collins, S.L.; van der Maarel, E. Mechanisms and processes in vegetation dynamics: Introduction. J. Veg. Sci. 1993, 4, 146–148. [Google Scholar] [CrossRef]
- Hörnberg, G.; Ohlson, M.; Zackrisson, O. Influence of bryophytes and microrelief conditions on Picea abies seed regeneration patterns in boreal old-growth swamp forests. Can. J. For. Res. 1997, 27, 1015–1023. [Google Scholar] [CrossRef]
- Schuur, E.A.G.; Abbott, B.W.; Bowden, W.B.; Brovkin, V.; Camill, P.; Canadell, J.G.; Chanton, J.P.; Chapin, F.S.; Christensen, T.R.; Ciais, P.; et al. Expert assessment of vulnerability of permafrost carbon to climate change. Clim. Chang. 2013, 119, 359–374. [Google Scholar] [CrossRef]
- DeColibus, D.T.; Rober, A.R.; Sampson, A.M.; Shurzinske, A.C.; Walls, J.T.; Turetsky, M.R.; Wyatt, K.H. Legacy effects of drought alters the aquatic food web of a northern boreal peatland. Freshw. Biol. 2017, 62, 1377–1388. [Google Scholar] [CrossRef]
- Gentili, F.; Nilsson, M.C.; Zackrisson, O.; DeLuca, T.H.; Sellstedt, A. Physiological and molecular diversity of feather moss associative N2-fixing cyanobacteria. J. Exp. Bot. 2005, 56, 3121–3127. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.C.; Wardle, D.A. Understory vegetation as a forest ecosystem driver: Evidence from the northern Swedish boreal forest. Eur. J. Phycol. 2005, 3, 421–428. [Google Scholar] [CrossRef]
- Angelini, C.; Silliman, B.R. Secondary foundation species as drivers of trophic and functional diversity: Evidence from a tree–epiphyte system. Ecology 2014, 95, 85–196. [Google Scholar] [CrossRef]
- Angelini, C.; van der Heide, T.; Griffin, J.N.; Morton, J.P.; Derksen-Hooijberg, M.; Lamers, L.P.; Smolders, A.J.; Silliman, B.R. Foundation species’ overlap enhances biodiversity and multifunctionality from the patch to landscape scale in southeastern United States salt marshes. Proc. R. Soc. B 2015, 282. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Siegel, D.I.; Zhu, C.; Chanton, J.P.; Glaser, P.H. Geochemical mixing in peatland waters: The role of organic acids. Wetlands 2015, 35, 567–575. [Google Scholar] [CrossRef]
- Siegel, D.I.; Glaser, P.H.; So, J.; Janecky, D.R. The dynamic balance between organic acids and circumneutral groundwater in a large boreal peat basin. J. Hydrol. 2006, 320, 421–431. [Google Scholar] [CrossRef]
- Glaser, P.H.; Hansen, B.; Siegel, D.I.; Reeve, A.S.; Morin, P.J. Rates, pathways and drivers for peatland development in the Hudson Bay Lowlands, northern Ontario, Canada. J. Ecol. 2004, 92, 1036–1053. [Google Scholar] [CrossRef]
- Bragazza, L.; Alber, R.; Gerdol, R. Seasonal chemistry of pore water in hummocks and hollows in a poor mire in the southern Alps (Italy). Wetlands 1998, 18, 320–328. [Google Scholar] [CrossRef]
- Hayward, P.M.; Clymo, R.S. The growth of Sphagnum: Experiments on, and simulation of some effects of light flux and water-table depth. J. Ecol. 1982, 71, 845–863. [Google Scholar] [CrossRef]
- Backéus, I. Production and depth distribution of fine roots in a boreal open bog. Ann. Bot. Fenn. 1986, 27, 261–265. [Google Scholar]
- Laiho, R.; Finer, L. Change in root biomass after water level drawdown on pine mires in southern Finland. Scand. J. For. Res. 1996, 11, 251–260. [Google Scholar] [CrossRef]
- Shetler, G.; Turetsky, M.R.; Kane, E.; Kasischke, E. Sphagnum mosses limit total carbon consumption during fire in Alaskan black spruce forests. Can. J. For. Res. 2008, 38, 2328–2336. [Google Scholar] [CrossRef]
- Thompson, D.K.; Waddington, J.M. Wildfire effects on vadose zone hydrology in forested boreal peatland microforms. J. Hydrol. 2013, 486, 48–56. [Google Scholar] [CrossRef]
- Zoltai, S.C.; Morissey, L.A.; Livingston, G.P.; Groot, W.J. Effects of fire on carbon cycling in North American boreal peatlands. Environ. Rev. 1998, 6, 13–24. [Google Scholar] [CrossRef]
- Hörnberg, G.; Zackrisson, O.; Segerström, U.; Svensson, B.W.; Ohlson, M.; Bradshaw, R.H.W. Boreal swamp forests: Biodiversity “hotspots” in an impoverished landscape. Bioscience 1998, 48, 795–802. [Google Scholar]
- Zoltai, S.C.; Taylor, S.J.; Jeglum, J.K.; Mills, G.F.; Johnson, J.D. Wetlands of Canada; National Wetlands Working Group, Polyscience Publictions Inc.: Montreal, QC, Canada, 1988; pp. 97–154. [Google Scholar]
- Lynch, J.A.; Clark, J.S.; Bigelow, N.H.; Edwards, M.E.; Finney, B.P. Geographic and temporal variations in fire history in boreal ecosystems of Alaska. J. Geophys. Res.-Atmos. 2003, 107, 8152. [Google Scholar] [CrossRef]
- Couwenberg, J.; Joosten, H. Self-organization in raised bog patterning: The origin of microtope zonation and mesotope diversity. J. Ecol. 2005, 93, 1238–1248. [Google Scholar] [CrossRef]
- Rietkerk, M.; Van de Koppel, J. Regular pattern formation in real ecosystems. Trends Ecol. Evol. 2008, 23, 169–175. [Google Scholar] [CrossRef]
- Iwaniec, D.M. Regulation and Organization of Periphyton from the Florida Everglades, U.S.A. Master’s Thesis, Florida International University, Miami, FL, USA, 2008. [Google Scholar]
- Wilson, J.B.; Agnew, A.D. Positive-feedback switches in plant communities. Adv. Ecol. Res. 1992, 23, 263–336. [Google Scholar]
- Thomsen, M.S.; Altieri, A.H.; Angelini, C.; Bishop, M.J.; Gribben, P.E.; Lear, G.; He, Q.; Schiel, D.R.; Silliman, B.R.; South, P.M.; et al. Secondary foundation species enhance biodiversity. Nat. Ecol. Evol. 2018, 2, 634–639. [Google Scholar] [CrossRef] [PubMed]
- van der Zee, E.M.; Angelini, C.; Govers, L.L.; Christianen, M.J.; Altieri, A.H.; van der Reijden, K.J.; Silliman, B.R.; van de Koppel, J.; van der Geest, M.; van Gils, J.A.; et al. How habitat-modifying organisms structure the food web of two coastal ecosystems. Proc. R. Soc. B 2016, 283, 20152326. [Google Scholar] [CrossRef] [PubMed]
- Heywood, V.H. (Ed.) Global biodiversity assessment. In United Nations Environment Program; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Simberloff, D. Flagships, umbrellas, and keystones: Is single-species management passé in the landscape era? Biol. Conserv. 1998, 83, 247–257. [Google Scholar] [CrossRef]
- Byers, J.E.; Cuddington, K.; Jones, C.G.; Talley, T.S.; Hastings, A.; Lambrinos, J.G.; Crooks, J.A.; Wilson, W.G. Using ecosystem engineers to restore ecological systems. Trends Ecol. Evol. 2006, 21, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Eldridge, D.J.; Zaady, E.; Shachak, M. Infiltration through three contrasting biological soil crusts in patterned landscapes in the Negev, Israel. CATENA 2000, 40, 323–336. [Google Scholar] [CrossRef]
- Rochefort, L. Sphagnum—A keystone genus in habitat restoration. Bryologist 2000, 103, 503–508. [Google Scholar] [CrossRef]
- Dudgeon, S.; Petraitis, P.S. First year demography of the foundation species, Ascophyllum nodosum, and its community implications. Oikos 2005, 109, 405–415. [Google Scholar] [CrossRef]
- Boers, A.M.; Zedler, J.B. Stabilized Water Levels and Typha Invasiveness. Wetlands 2008, 28, 676–685. [Google Scholar] [CrossRef]
- Lagerwall, G.; Kiker, G.; Muñoz-Carpena, R.; Wang, N. Accounting for the Impact of Management Scenarios on Typha domingensis (Cattail) in an Everglades Wetland. Environ. Manag. 2017, 59, 129–140. [Google Scholar] [CrossRef]
- McCormick, P.V.; Newman, S.; Vilchek, L.W. Landscape responses to wetland eutrophication: Loss of slough habitat in the Florida Everglades, USA. Hydrobiologia 2009, 621, 105–114. [Google Scholar] [CrossRef]
- Richardson, C.J.; King, R.S.; Qian, S.S.; Vaithiyanathan, P.; Qualls, R.G.; Stow, C.A. Estimating ecological thresholds for phosphorus in the Everglades. Environ. Sci. Technol. 2007, 41, 8084–8091. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, P.R.; Davis, S.E., III; van Lent, T.; Davis, S.M.; Henriquez, H. Science synthesis for management as a way to advance ecosystem restoration: Evaluation of restoration scenarios for the Florida Everglades. Restor. Ecol. 2017, 25, S4–S17. [Google Scholar] [CrossRef]
- Naja, M.; Childers, D.L.; Gaiser, E.E. Water quality implications of hydrologic restoration alternatives in the Florida Everglades, United States. Restor. Ecol. 2017, 25 (Suppl. 1), S48–S58. [Google Scholar] [CrossRef]
- Larsen, L.G.; Eppinga, M.B.; Passalacqua, P.; Getz, W.M.; Rose, K.A.; Liang, M. Appropriate complexity landscape modeling. Earth Sci. Rev. 2016, 160, 111–130. [Google Scholar] [CrossRef]
- Larsen, L.G.; Eppinga, M.; Coulthard, T.; Thomas, C. Exploratory modelling: Extracting causality from complexity. EOS 2014, 95, 285–292. [Google Scholar] [CrossRef]
- Zhai, L.; Jiang, J.; DeAngelis, D.; Sternberg, L.D.S.L. Prediction of plant vulnerability to salinity increase in a coastal ecosystem by stable isotope composition (δ 18 O) of plant stem water: A model study. Ecosystems 2016, 19, 32–49. [Google Scholar] [CrossRef]
- Pearlstine, L.; Friedman, S.; Supernaw, M.; Team, E.M. Everglades Landscape Vegetation Succession Model (ELVeS) Ecological and Design Document: Freshwater Marsh & Prairie Component Version 1.1; South Florida Natural Resources Center, Everglades National Park, National Park Service: Homestead, FL, USA, 2011.
- Twilley, R.R.; Rivera-Monroy, V.H. Developing Performance Measures of Mangrove Wetlands Using Simulation Models of Hydrology, Nutrient Biogeochemistry, and Community Dynamics. J. Coast. Res. 2005, 40, 79–93. [Google Scholar]
- Twilley, R.R.; Rivera-Monroy, V.H.; Chen, R.; Botero, L. Adapting an ecological mangrove model to simulate trajectories in restoration ecology. Mar. Pollut. Bull. 1999, 37, 404–419. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).