Food Shortage Amplifies Negative Sublethal Impacts of Low-Level Exposure to the Neonicotinoid Insecticide Imidacloprid on Stream Mayfly Nymphs


Abstract
1. Introduction
- Exposure to low concentrations of imidacloprid results in decreased survivorship, increased occurrence of impairment and immobility [6], and reduced moulting frequency in Deleatidium. Moulting is controlled by the endocrine and nervous systems; therefore, neurological disruption due to imidacloprid may interfere with moulting [50], which has been observed in a previous 96-h imidacloprid exposure involving Deleatidium nymphs [41].
- A higher density of mayfly individuals intensifies the effect of the starvation period, with the resulting increase in competition for resources also amplifying the predicted adverse effects of imidacloprid on all four mayfly responses, similarly to above. Density-dependent increases in competition intensity are common in ecological systems [51].
2. Materials and Methods
2.1. Mayfly Food Supply
2.2. Mayfly Collection
2.3. Experimental Design
2.4. Determination of Mayfly Responses
2.5. Validation of Imidacloprid Concentrations
2.6. Data Analysis
3. Results
3.1. Achieved Imidacloprid Concentrations
3.2. Sample Size
3.3. Mayfly Responses
3.3.1. Survivorship
3.3.2. Impairment
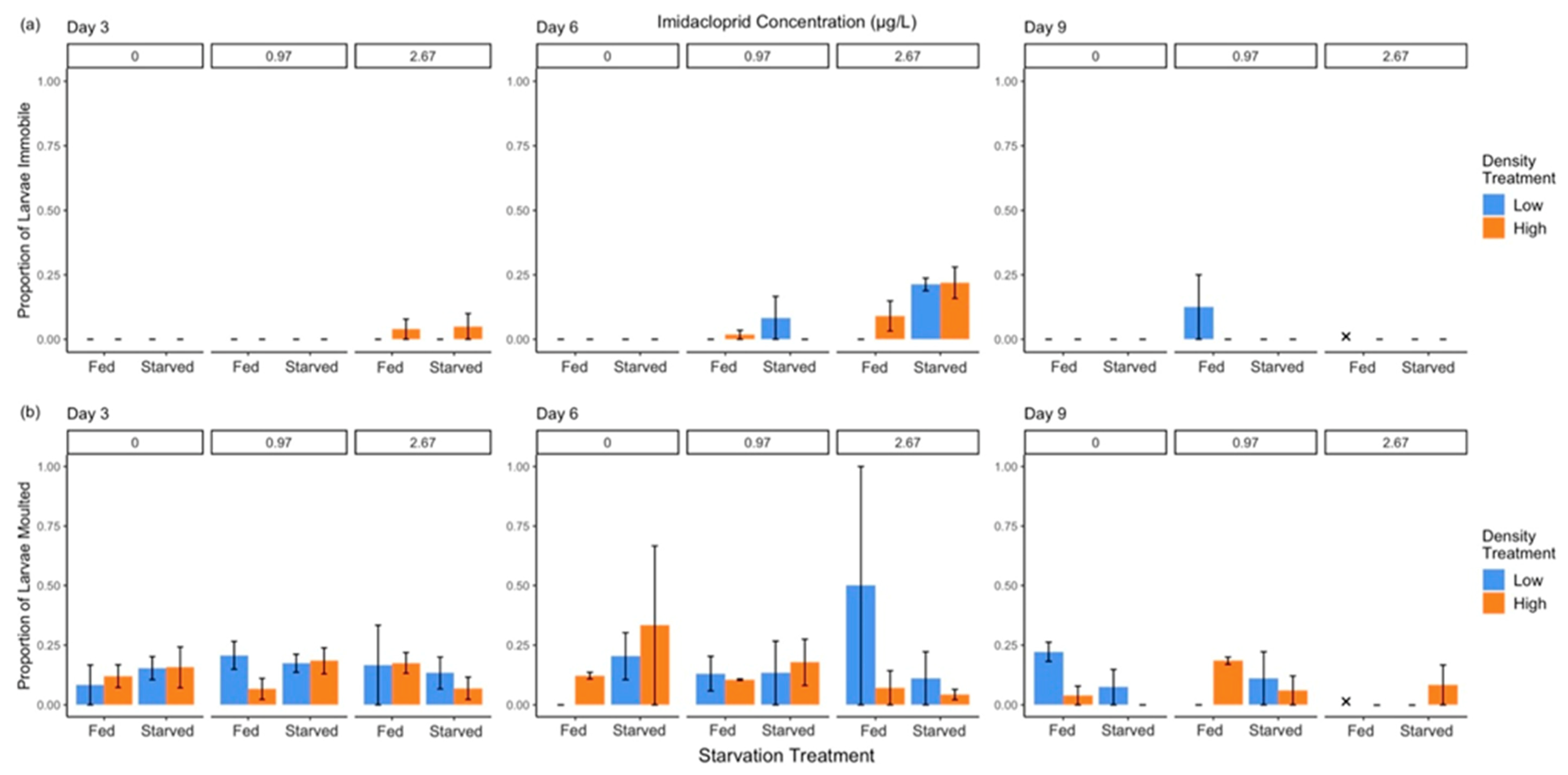
3.3.3. Immobility and Moulting
4. Discussion
4.1. Imidacloprid Effects on Mayflies
4.2. Starvation Effects and Interactions with Imidacloprid
4.3. Mayfly Density Effects and Interactions with Starvation or Imidacloprid
4.4. Which Are the Most Informative Sublethal Mayfly Responses?
4.5. Limitations of Our Study
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jeschke, P.; Nauen, R.; Schindler, M.; Elbert, A. Overview of the status and global strategy for neonicotinoids. J. Agric. Food Chem. 2010, 59, 2897–2908. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D. An overview of the environmental risks posed by neonicotinoid insecticides. J. Appl. Ecol. 2013, 50, 977–987. [Google Scholar] [CrossRef]
- Lewis, K.A.; Tzilivakis, J.; Warner, D.; Green, A. An international database for pesticide risk assessments and management. Hum. Ecol. Risk Assess. 2016, 22, 1050–1064. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Hyne, R.V. Detection and analysis of neonicotinoids in river waters—Development of a passive sampler for three commonly used insecticides. Chemosphere 2014, 143–151. [Google Scholar] [CrossRef]
- Morrissey, C.A.; Mineau, P.; Devries, J.H.; Sánchez-Bayo, F.; Liess, M.; Cavallaro, M.C.; Liber, K. Neonicotinoid contamination of global surface waters and associated risk to aquatic invertebrates: A review. Environ. Int. 2015, 74, 291–303. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Goka, K.; Hayasaka, D. Contamination of the aquatic environment with neonicotinoids and its implication for ecosystems. Front. Environ. Sci. 2016, 4, 1–14. [Google Scholar] [CrossRef]
- Van Dijk, T.C.; Van Staalduinen, M.A.; Van der Sluijs, J.P. Macro-invertebrate decline in surface water polluted with imidacloprid. PLoS ONE 2013, 8, e62374. [Google Scholar] [CrossRef]
- Kreuger, J.; Graaf, S.; Patring, J.; Adielsson, S. Pesticides in Surface Water in Areas with Open Ground and Greenhouse Horticultural Crops In Sweden 2008. Technical Report. Swedish University of Agricultural Sciences. Available online: https://pub.epsilon.slu.se/5413/ (accessed on 1 May 2019).
- Starner, K.; Goh, K.S. Detections of the neonicotinoid insecticide imidacloprid in surface waters of three agricultural regions of California, USA, 2010–2011. Bull. Environ. Contam. Toxicol. 2012, 88, 316–321. [Google Scholar] [CrossRef]
- Hladik, M.L.; Kolpin, D.W. First national-scale reconnaissance of neonicotinoid insecticides in streams across the USA. Environ. Chem. 2015, 13, 12–20. [Google Scholar] [CrossRef]
- Stehle, S.; Bub, S.; Schulz, R. Compilation and analysis of global surface water concentrations for individual insecticide compounds. Sci. Total Environ. 2018, 639, 516–525. [Google Scholar] [CrossRef]
- Aquatic Life Benchmarks and Ecological Risk Assessments for Registered Pesticides. United States Environmental Protection Agency. Available online: https://www.epa.gov/pesticide-science-and-assessing-pesticide-risks/aquatic-life-benchmarks-and-ecological-risk (accessed on 1 September 2019).
- Simon-Delso, N.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Chagnon, M.; Downs, C.; Furlan, L.; Gibbons, D.W.; Giorio, C.; Girolami, V.; et al. Systemic insecticides (neonicotinoids and fipronil): Trends, uses, mode of action and metabolites. Environ. Sci. Pollut. Res. 2015, 22, 5–34. [Google Scholar] [CrossRef] [PubMed]
- Tomizawa, M.; Casida, J.E. Neonicotinoid insecticide toxicology: Mechanisms of selective action. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 247–265. [Google Scholar] [CrossRef] [PubMed]
- Bonmatin, J.M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; Long, M.; Marzaro., M.; Mitchell, E.A.D.; et al. Environmental fate and exposure; neonicotinoids and fipronil. Environ. Sci. Pollut. Res. 2015, 22, 35–67. [Google Scholar] [CrossRef] [PubMed]
- Pisa, L.W.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Downs, C.A.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; McField, M.; et al. Effects of neonicotinoids and fipronil on non-target invertebrates. Environ. Sci. Pollut. Res. 2015, 22, 68–102. [Google Scholar] [CrossRef]
- Stoughton, S.J.; Liber, K.; Culp, J.; Cessna, A. Acute and chronic toxicity of imidacloprid to the aquatic invertebrates Chironomus tentans and Hyalella azteca under constant- and pulse-exposure conditions. Arch. Environ. Contam. Toxicol. 2008, 54, 662–673. [Google Scholar] [CrossRef]
- Pestana, J.L.; Loureiro, S.; Baird, D.J.; Soares, A.M. Fear and loathing in the benthos: Responses of aquatic insect larvae to the pesticide imidacloprid in the presence of chemical signals of predation risk. Aquat. Toxicol. 2009, 93, 138–149. [Google Scholar] [CrossRef]
- LeBlanc, H.M.K.; Culp, J.M.; Baird, D.J.; Alexander, A.C.; Cessna, A.J. Single versus combined lethal effects of three agricultural insecticides on larvae of the freshwater insect Chironomus dilutus. Arch. Environ. Contam. Toxicol. 2012, 63, 378–390. [Google Scholar] [CrossRef]
- Malev, O.; Klobucar, R.S.; Fabbretti, E.; Trebse, P. Comparative toxicity of imidacloprid and its transformation product 6-chloronicotinic acid to non-target aquatic organisms: Microalgae Desmodesmus subspicatus and amphipod Gammarus fossarum. Pestic. Biochem. Physiol. 2012, 104, 178–186. [Google Scholar] [CrossRef]
- Nyman, A.; Hintermeister, A.; Schirmer, K.; Ashauer, R. The insecticide imidacloprid causes mortality of the freshwater amphipod Gammarus pulex by interfering with feeding behavior. PLoS ONE 2013, 8, e62472. [Google Scholar] [CrossRef]
- Alexander, A.C.; Culp, J.M.; Liber, K.; Cessna, A.J. Effects of insecticide exposure on feeding inhibition in mayflies and oligochaetes. Environ. Toxicol. Chem. 2007, 26, 1726–1732. [Google Scholar] [CrossRef]
- Roessink, I.; Merga, L.B.; Zweers, H.J.; Van den Brink, P.J. The neonicotinoid imidacloprid shows high chronic toxicity to mayfly nymphs. Environ. Toxicol. Chem. 2013, 32, 1096–1100. [Google Scholar] [CrossRef] [PubMed]
- Maxted, J.R.; Barbour, M.T.; Gerritsen, J.; Proetti, V.; Primrose, N.; Silvia, A.; Penrose, D.; Renfrow, R. Assessment framework for mid-Atlantic coastal plain streams using benthic macroinvertebrates. J. Am. Benthol. Soc. 2000, 19, 128–144. [Google Scholar] [CrossRef]
- Klemm, D.J.; Blocksom, K.A.; Fulk, F.A.; Herlihy, A.T.; Hughes, R.M.; Kaufmann, P.R.; Peck, D.V.; Stoddard, J.L.; Thoeny, W.T.; Griffith, M.B.; et al. Development and evaluation of a macroinvertebrate biotic integrity index (MBII) for regionally assessing mid–Atlantic highlands streams. Environ. Manag. 2003, 31, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Vignati, D.A.; Ferrari, B.J.; Dominik, J. Laboratory–to–field extrapolation in aquatic sciences. Environ. Sci. Technol. 2007, 41, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Bednarska, A.J.; Jevtić, D.M.; Laskowski, R. More ecological ERA: Incorporating natural environmental factors and animal behavior. Integr. Environ. Assess. Manag. 2013, 9, e39–e46. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Tennekes, H.A. Environmental risk assessment of agrochemicals—A critical appraisal of current approaches. In Toxicity and Hazard of Agrochemicals; Larramendy, M., Soloneski, S., Eds.; InTechOpen: London, UK, 2015; Available online: https://www.intechopen.com/books/toxicity-and-hazard-of-agrochemicals/environmental-risk-assessment-of-agrochemicals-a-critical-appraisal-of-current-approaches (accessed on 1 October 2019). [CrossRef]
- Buchwalter, D.B.; Clements, W.H.; Luoma, S.N. Modernizing water quality criteria in the United States: A need to expand the definition of acceptable data. Environ. Toxicol. Chem. 2017, 36, 285–291. [Google Scholar] [CrossRef]
- EFSA Panel on Plant Protection Products and their Residues (PPR). Guidance on tiered risk assessment for plant protection products for aquatic organisms in edge-of-field surface waters. EFSA J. 2013, 11, 3290. [Google Scholar] [CrossRef]
- Stehle, S.; Schulz, R. Pesticide authorization in the EU—Environment unprotected? Environ. Sci. Pollut. Res. 2015, 22, 19632–19647. [Google Scholar] [CrossRef]
- Liess, M.; von der Ohe, P.C. Analyzing effects of pesticides on invertebrate communities in streams. Environ. Toxicol. Chem. 2005, 24, 954–965. [Google Scholar] [CrossRef]
- Schäfer, R.B.; von der Ohe, P.C.; Rasmussen, J.; Kefford, B.J.; Beketov, M.A.; Schulz, R.; Liess, M. Thresholds for the effects of pesticides on invertebrate communities and leaf breakdown in stream ecosystems. Environ. Sci. Technol. 2012, 46, 5134–5142. [Google Scholar] [CrossRef]
- Beketov, M.A.; Kefford, B.J.; Schäfer, R.B.; Liess, M. Pesticides reduce regional biodiversity of stream invertebrates. Proc. Natl. Acad. Sci. USA 2013, 110, 11039–11043. [Google Scholar] [CrossRef] [PubMed]
- Szöcs, E.; Brinke, M.; Karaoglan, B.; Schäfer, R.B. Large scale risks from agricultural pesticides in small streams. Environ. Sci. Technol. 2017, 51, 7378–7385. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Becker, J.M.; Liess, M. Sequential exposure to low levels of pesticides and temperature stress increase toxicological sensitivity of crustaceans. Sci. Total Environ. 2018, 610, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Liess, M. Population response to toxicants is altered by intraspecific interaction. Environ. Toxicol. Chem. 2002, 21, 138–142. [Google Scholar] [CrossRef]
- Holmstrup, M.; Bindesbøl, A.M.; Oostingh, G.J.; Duschl, A.; Scheil, V.; Köhler, H.R.; Loureiro, S.; Soares, A.M.V.M.; Ferreira, A.L.G.; Kienle, C.; et al. Interactions between effects of environmental chemicals and natural stressors: A review. Sci. Total Environ. 2010, 408, 3746–3762. [Google Scholar] [CrossRef]
- Laskowski, R.; Bednarska, A.J.; Kramarz, P.E.; Loureiro, S.; Scheil, V.; Kudłek, J.; Holmstrup, M. Interactions between toxic chemicals and natural environmental factors—A meta–analysis and case studies. Sci. Total Environ. 2010, 408, 3763–3774. [Google Scholar] [CrossRef]
- Heugens, E.H.; Hendriks, A.J.; Dekker, T.; Straalen, N.M.V.; Admiraal, W. A review of the effects of multiple stressors on aquatic organisms and analysis of uncertainty factors for use in risk assessment. Crit. Rev. Toxicol. 2001, 31, 247–284. [Google Scholar] [CrossRef]
- Macaulay, S.J.; Buchwalter, D.B.; Matthaei, C.D. Water temperature interacts with the insecticide imidacloprid to increase lethal and sublethal toxicity to mayfly larvae. N. Z. J. Mar. Freshw. Res. 2019. [Google Scholar] [CrossRef]
- Hopkins, W.A.; Snodgrass, J.W.; Roe, J.H.; Staub, B.P.; Jackson, B.P.; Congdon, J.D. Effects of food ration on survival and sublethal responses of lake chubsuckers (Erimyzon sucetta) exposed to coal combustion wastes. Aquat. Toxicol. 2002, 57, 191–202. [Google Scholar] [CrossRef]
- Metcalfe, N.B.; Monaghan, P. Compensation for a bad start: Grow now, pay later? Trends Ecol. Evol. 2001, 16, 254–260. [Google Scholar] [CrossRef]
- Storey, K.B. Regulation of hypometabolism: Insights into epigenetic controls. J. Exp. Biol. 2015, 218, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Bridges, T.S.; Farrar, J.D.; Duke, B.M. The influence of food ration on sediment toxicity in Neanthes arenaceodentata (Annelida: Polychaeta). Environ. Toxicol. Chem. 1997, 16, 1659–1665. [Google Scholar] [CrossRef]
- Mommaerts, V.; Reynders, S.; Boulet, J.; Besard, L.; Sterk, G.; Smagghe, G. Risk assessment for side–effects of neonicotinoids against bumblebees with and without impairing foraging behavior. Ecotoxicology 2010, 19, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Foit, K.; Kaske, O.; Liess, M. Competition increases toxicant sensitivity and delays the recovery of two interacting populations. Aquat. Toxicol. 2012, 106, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Liess, M.; Foit, K.; Becker, A.; Hassold, E.; Dolciotti, I.; Kattwinkel, M.; Duquesne, S. Culmination of low–dose pesticide effects. Environ. Sci. Technol. 2013, 47, 8862–8868. [Google Scholar] [CrossRef]
- McIntosh, A.R.; Townsend, C.R. Impacts of an introduced predatory fish on mayfly grazing in New Zealand streams. Limnol. Oceanogr. 1995, 40, 1508–1512. [Google Scholar] [CrossRef]
- Song, M.Y.; Stark, J.D.; Brown, J.J. Comparative toxicity of four insecticides, including imidacloprid and tebufenozide, to four aquatic arthropods. Environ. Toxicol. Chem. 1997, 16, 2494–2500. [Google Scholar] [CrossRef]
- Gurevitch, J.; Morrow, L.L.; Wallace, A.; Walsh, J.S. A metaanalysis of competition in field experiments. Am. Nat. 1992, 140, 539–572. [Google Scholar] [CrossRef]
- Peckarsky, B.L.; McIntosh, A.R. Fitness and community consequences of avoiding multiple predators. Oecologia 1998, 113, 565–576. [Google Scholar] [CrossRef]
- Beketov, M.; Liess, M. Potential of 11 pesticides to initiate downstream drift of stream macroinvertebrates. Arch. Environ. Contam. Toxicol. 2008, 55, 247–253. [Google Scholar] [CrossRef]
- Hunn, J.G.; (University of Otago, Dunedin, NZ). Personal Communication, 2016.
- Hunn, J.G. Imidacloprid Effects on New Zealand Mayfly Nymphs: Acute and Chronic Exposure and Interactions with Natural Stressors. Bachelor Thesis, University of Otago, Dunedin, New Zealand, October 2016. [Google Scholar]
- Macaulay, S.J. Insecticide Contamination of Streams in a Warming Climate. Ph.D. Thesis, University of Otago, Dunedin, New Zealand, June 2019. [Google Scholar]
- Garson, D. Multivariate GLM, MANOVA, and MANCOVA 2015 Edition. Available online: www.statisticalassociates.com (accessed on 5 October 2019).
- Nakagawa, S. A farewell to Bonferroni: The problems of low statistical power and publication bias. Behav. Ecol. 2004, 15, 1044–1045. [Google Scholar] [CrossRef]
- Nakagawa, S.; Cuthill, I.C. Effect size, confidence interval and statistical significance: A practical guide for biologists. Biol. Rev. 2007, 82, 591–605. [Google Scholar] [CrossRef] [PubMed]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Tennekes, H.A.; Sánchez-Bayo, F. The molecular basis of simple relationships between exposure concentration and toxic effects with time. Toxicology 2013, 309, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, S.D.; Lapied, B.; le Corronc, H.L.; Sattelle, F. Imidacloprid actions on insect neuronal acetylcholine receptors. J. Exp. Biol. 1997, 200, 2685–2692. [Google Scholar] [PubMed]
- Zhang, A.; Kaiser, H.; Maienfisch, P.; Casida, J.E. Insect nicotinic acetylcholine receptor: Conserved neonicotinoid specificity of [3H] imidacloprid binding site. J. Neurochem. 2000, 75, 1294–1303. [Google Scholar] [CrossRef] [PubMed]
- Chapman, P.M. Integrating toxicology and ecology: Putting the “eco” into ecotoxicology. Mar. Pollut. Bull. 2002, 44, 7–15. [Google Scholar] [CrossRef]
- Gessner, M.O.; Tlili, A. Fostering integration of freshwater ecology with ecotoxicology. Freshw. Biol. 2016, 61, 1991–2001. [Google Scholar] [CrossRef]
- Maloney, E.M. How do we take the pulse of an aquatic ecosystem? Current and historical approaches to measuring ecosystem integrity. Environ. Toxicol. Chem. 2019, 38, 289–301. [Google Scholar] [CrossRef]
- Dinh, K.V.; Janssens, L.; Stoks, R. Exposure to a heat wave under food limitation makes an agricultural insecticide lethal: A mechanistic laboratory experiment. Glob. Chang. Biol. 2016, 22, 3361–3372. [Google Scholar] [CrossRef]
- Janssens, L.; Stoks, R. Fitness effects of chlorpyrifos in the damselfly Enallagma cyathigerum strongly depend upon temperature, food level and can bridge metamorphosis. PLoS ONE 2013, 8, e68107. [Google Scholar] [CrossRef]
- Mydlarz, L.D.; Jones, L.E.; Harvell, C.D. Innate immunity, environmental drivers, and disease ecology of marine and freshwater invertebrates. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 251–288. [Google Scholar] [CrossRef]
- DeBlock, M.; Stoks, R. Short-term larval food stress and associated compensatory growth reduce adult immune function in a damselfly. Ecol. Entomol. 2008, 33, 796–801. [Google Scholar] [CrossRef]
- Muturi, E.J.; Costanzo, K.; Kesavaraju, B.; Lampman, R.; Alto, B.W. Interaction of a pesticide and larval competition on life history traits of Culex pipiens. Acta Trop. 2010, 116, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Alexander, A.C.; Culp, J.M.; Baird, D.J.; Cessna, A.J. Nutrient–insecticide interactions decouple density–dependent predation pressure in aquatic insects. Freshw. Biol. 2016, 61, 2090–2101. [Google Scholar] [CrossRef]
- Effenberger, M.; Sailer, G.; Townsend, C.R.; Matthaei, C.D. Local disturbance history and habitat parameters influence the microdistribution of stream invertebrates. Freshw. Biol. 2006, 51, 312–332. [Google Scholar] [CrossRef]
- Camp, A.A.; Buchwalter, D.B. Can’t take the heat: Temperature-enhanced toxicity in the mayfly Isonychia bicolor exposed to the neonicotinoid insecticide imidacloprid. Aquat. Toxicol. 2016, 178, 49–57. [Google Scholar] [CrossRef]
- Van den Brink, P.J.; Van Smeden, J.M.; Bekele, R.S.; Dierick, W.; De Gelder, D.M.; Noteboom, M.; Roessink, I. Acute and chronic toxicity of neonicotinoids to nymphs of a mayfly species and some notes on seasonal differences. Environ. Toxicol. Chem. 2016, 35, 128–133. [Google Scholar] [CrossRef]
- Raby, M.; Zhao, X.M.; Hao, C.Y.; Poirier, D.G.; Sibley, P.K. Chronic effects of an environmentally–relevant, short–term neonicotinoid insecticide pulse on four aquatic invertebrates. Sci. Total Environ. 2018, 639, 1543–1552. [Google Scholar] [CrossRef]
- Raby, M.; Nowierski, M.; Perlov, D.; Zhao, X.; Hao, C.; Poirier, D.G.; Sibley, P.K. Acute toxicity of 6 neonicotinoid insecticides to freshwater invertebrates. Environ. Toxicol. Chem. 2018, 37, 1430–1445. [Google Scholar] [CrossRef]
- Raby, M.; Maloney, E.; Poirier, D.G.; Sibley, P.K. Acute effects of binary mixtures of imidacloprid and tebuconazole on 4 freshwater invertebrates. Environ. Toxicol. Chem. 2019, 38, 1093–1103. [Google Scholar] [CrossRef]
- Macaulay, S.J.; Hageman, K.J.; Alumbaugh, R.E.; Lyons, S.M.; Piggott, J.J.; Matthaei, C.D. Chronic toxicities of neonicotinoids to nymphs of the common New Zealand mayfly Deleatidium spp. Environ Toxicol. Chem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Diamond, J.M.; Winchester, E.L.; Mackler, D.G.; Gruber, D. Use of the mayfly Stenonema modestum (Heptageniidae) in subacute toxicity assessments. Environ. Toxicol. Chem. 1992, 11, 415–425. [Google Scholar] [CrossRef]
- Camp, A.; Funk, D.; Buchwalter, D.B. A stressful shortness of breath: Molting disrupts breathing in the mayfly Cloeon dipterum. Freshw. Sci. 2014, 33, 695–699. [Google Scholar] [CrossRef]
- Magbanua, F.S.; Townsend, C.R.; Hageman, K.J.; Lange, K.; Lear, G.; Lewis, G.D.; Matthaei, C.D. Understanding the combined influence of fine sediment and glyphosate herbicide on stream periphyton communities. Water Res. 2013, 47, 5110–5120. [Google Scholar] [CrossRef]
- Magbanua, F.S.; Townsend, C.R.; Hageman, K.J.; Matthaei, C.D. Individual and combined effects of fine sediment and the herbicide glyphosate on benthic macroinvertebrates and stream ecosystem function. Freshw. Biol. 2013, 58, 1729–1744. [Google Scholar] [CrossRef]
- Chapman, R.B. A Review of Insecticide Use on Pastures and Forage Crops in New Zealand. Available online: http://agpest.co.nz/wp-content/uploads/2013/06/A-review-of-insecticide-use-on-pastures-and-forage-crops-in-New-Zealand.pdf (accessed on 2 October 2019).
- Hageman, K.J.; Aebig, C.H.F.; Luong, K.H.; Kaserzon, S.L.; Wong, C.S.; Reeks, T.; Greenwood, M.; Macaulay, S.; Matthaei, C.D. Current–use pesticides in New Zealand streams: Comparing results from grab samples and three types of passive samplers. Environ. Pollut. 2019, 254, 112973. [Google Scholar] [CrossRef]
- Moreau, M.; Hadfield, J.; Hughey, J.; Sanders, F.; Lapworth, D.J.; White, D.; Civil, W. A baseline assessment of emerging organic contaminants in New Zealand groundwater. Sci. Total Environ. 2019, 686, 425–439. [Google Scholar] [CrossRef]
- Magbanua, F.S.; Townsend, C.R.; Blackwell, G.L.; Matthaei, C.D. Responses of stream macroinvertebrates and ecosystem function to conventional, integrated and organic farming. J. Appl. Ecol. 2010, 47, 1014–1025. [Google Scholar] [CrossRef]
- Shahpoury, P.; Hageman, K.J.; Matthaei, C.D.; Magbanua, F.S. Chlorinated pesticides in stream sediments from organic, integrated and conventional farms. Environ. Pollut. 2013, 181, 219–225. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Conc. (μg L−1) | Starved Y/N | Density | Day 3 | Day 6 | Day 9 |
|---|---|---|---|---|---|
| 0.0 | N | 10 | 3 | 3 | 3 |
| 0.0 | N | 20 | 3 | 3 | 3 |
| 0.0 | Y | 10 | 3 | 3 | 3 |
| 0.0 | Y | 20 | 3 | 3 | 3 |
| 0.97 | N | 10 | 3 | 3 | 2 |
| 0.97 | N | 20 | 3 | 3 | 3 |
| 0.97 | Y | 10 | 3 | 3 | 2 |
| 0.97 | Y | 20 | 3 | 3 | 3 |
| 2.67 | N | 10 | 3 | 2 | 0 |
| 2.67 | N | 20 | 3 | 3 | 1 |
| 2.67 | Y | 10 | 3 | 3 | 1 |
| 2.67 | Y | 20 | 3 | 3 | 2 |
| Total | - | - | 36 | 35 | 26 |
| Sampling Day | Imidacloprid Concentration (IMI) p (ES) Ranking | Starvation p(ES) | Density p | Starved × IMI p (ES) | Density × IMI p (ES) | Starved × Density p (ES) | Starved × Density × IMI p |
|---|---|---|---|---|---|---|---|
| Day 3 | 0.01 (0.32) 0.97 > 2.67 | 0.34 | 0.48 | 0.001(0.43) | 0.14 | 0.16 | 0.96 |
| Day 6 | 0.007 (0.34) (0 = 0.97) > 2.67 | 0.27 | 0.94 | <0.001(0.49) | 0.11 | 0.01 (0.25) | 0.84 |
| Day 9 | <0.001 (0.67) 0 > 0.097 > 2.67 | 0.01 (0.27) | 0.97 | 0.03 (0.25) | 0.03 (0.25) | 0.01 (0.27) | 0.15 |
| Sampling Day | Imidacloprid Concentration p (ES) Ranking | Starvation p (ES) | Density p (ES) | Starved × IMI p (ES) | Density × IMI p (ES) | Starved × Density p | Starved × Density × IMI p |
|---|---|---|---|---|---|---|---|
| Day 3 | <0.001 (0.80) (0 = 0.97) < 2.67 | 0.001 (0.36) | 0.113 | <0.001(0.47) | 0.21 | 0.84 | 0.93 |
| Day 6 | <0.001 (0.91) 0 < 0.97 < 2.67 | 0.01 (0.26) | 0.22 | 0.16 | 0.12 | 0.18 | 0.13 |
| Day 9 | <0.001 (0.76) 0 < (0.97 = 2.67) | 0.40 | 0.01 (0.35) | 0.002 (0.56) | 0.03(0.38) | 0.80 | 0.80 |
| Sampling Day | Imidacloprid Concentration p (ES) Ranking | Starvation p (ES) | Density p | Starved × IMI p (ES) | Density × IMI p | Starved × Density p | Starved × Density × IMI p |
|---|---|---|---|---|---|---|---|
| Day 3 | 0.16 | 0.87 | 0.17 | 0.97 | 0.16 | 0.87 | 0.97 |
| 0.60 | 0.61 | 0.50 | 0.76 | 0.47 | 0.87 | 0.38 | |
| Day 6 | <0.001 (0.54) (0 = 0.97) < 2.67 | 0.004 (0.31) | 0.81 | 0.01(0.34) | 0.32 | 0.16 | 0.58 |
| 0.56 | 0.54 | 0.78 | 0.93 | 0.78 | 0.31 | 0.29 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hunn, J.G.; Macaulay, S.J.; Matthaei, C.D. Food Shortage Amplifies Negative Sublethal Impacts of Low-Level Exposure to the Neonicotinoid Insecticide Imidacloprid on Stream Mayfly Nymphs. Water 2019, 11, 2142. https://doi.org/10.3390/w11102142
Hunn JG, Macaulay SJ, Matthaei CD. Food Shortage Amplifies Negative Sublethal Impacts of Low-Level Exposure to the Neonicotinoid Insecticide Imidacloprid on Stream Mayfly Nymphs. Water. 2019; 11(10):2142. https://doi.org/10.3390/w11102142
Chicago/Turabian StyleHunn, Julia G., Samuel J. Macaulay, and Christoph D. Matthaei. 2019. "Food Shortage Amplifies Negative Sublethal Impacts of Low-Level Exposure to the Neonicotinoid Insecticide Imidacloprid on Stream Mayfly Nymphs" Water 11, no. 10: 2142. https://doi.org/10.3390/w11102142
APA StyleHunn, J. G., Macaulay, S. J., & Matthaei, C. D. (2019). Food Shortage Amplifies Negative Sublethal Impacts of Low-Level Exposure to the Neonicotinoid Insecticide Imidacloprid on Stream Mayfly Nymphs. Water, 11(10), 2142. https://doi.org/10.3390/w11102142