Flow Regime and Nutrient-Loading Trends from the Largest South European Watersheds: Implications for the Productivity of Mediterranean and Black Sea’s Coastal Areas





Abstract
:
1. Introduction
2. Materials and Methods
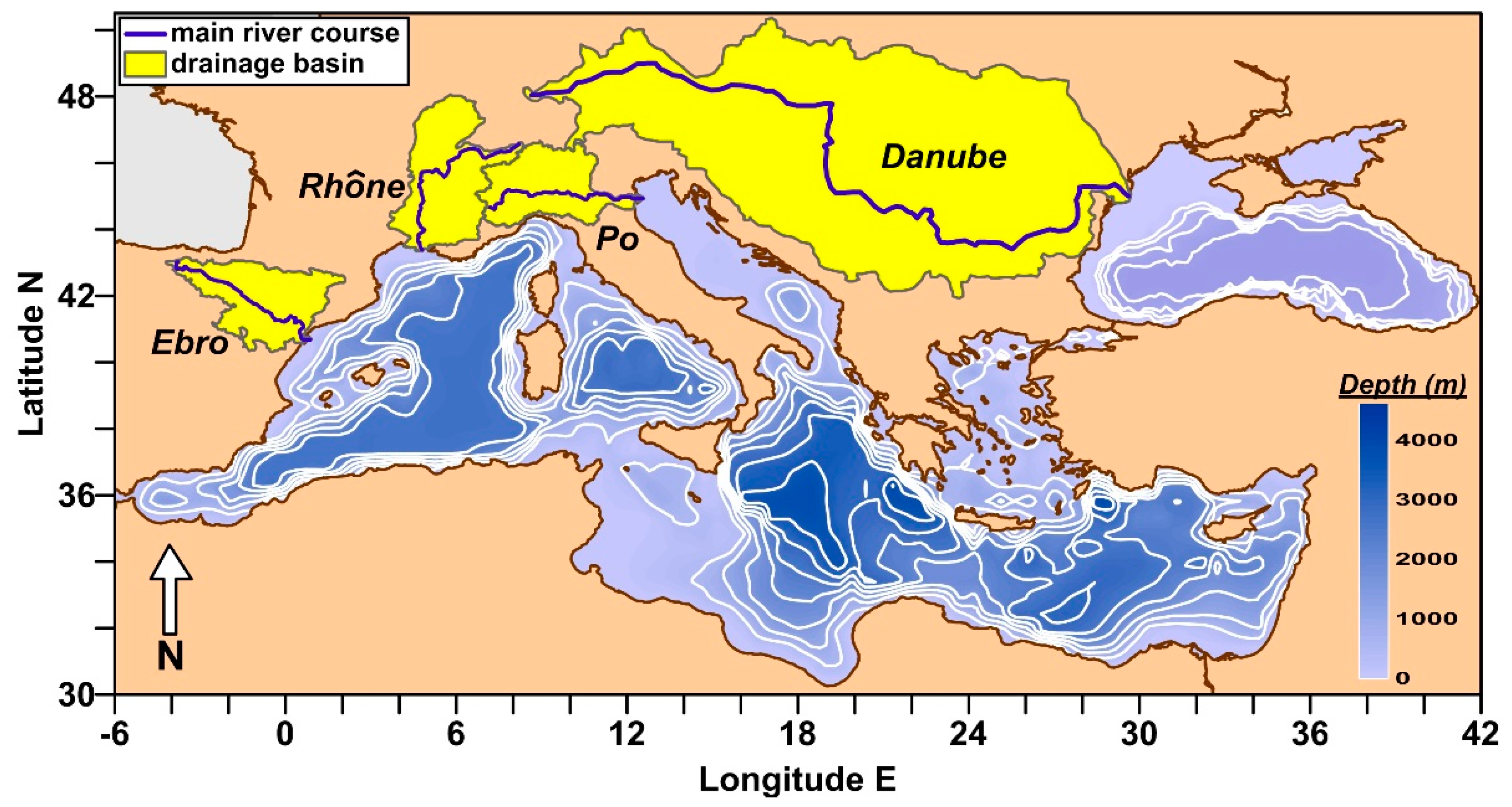
2.1. Geographical Settings of River Basins
2.2. River Flow Data
2.3. Chemical Parameters
2.4. Data Analysis
3. Results
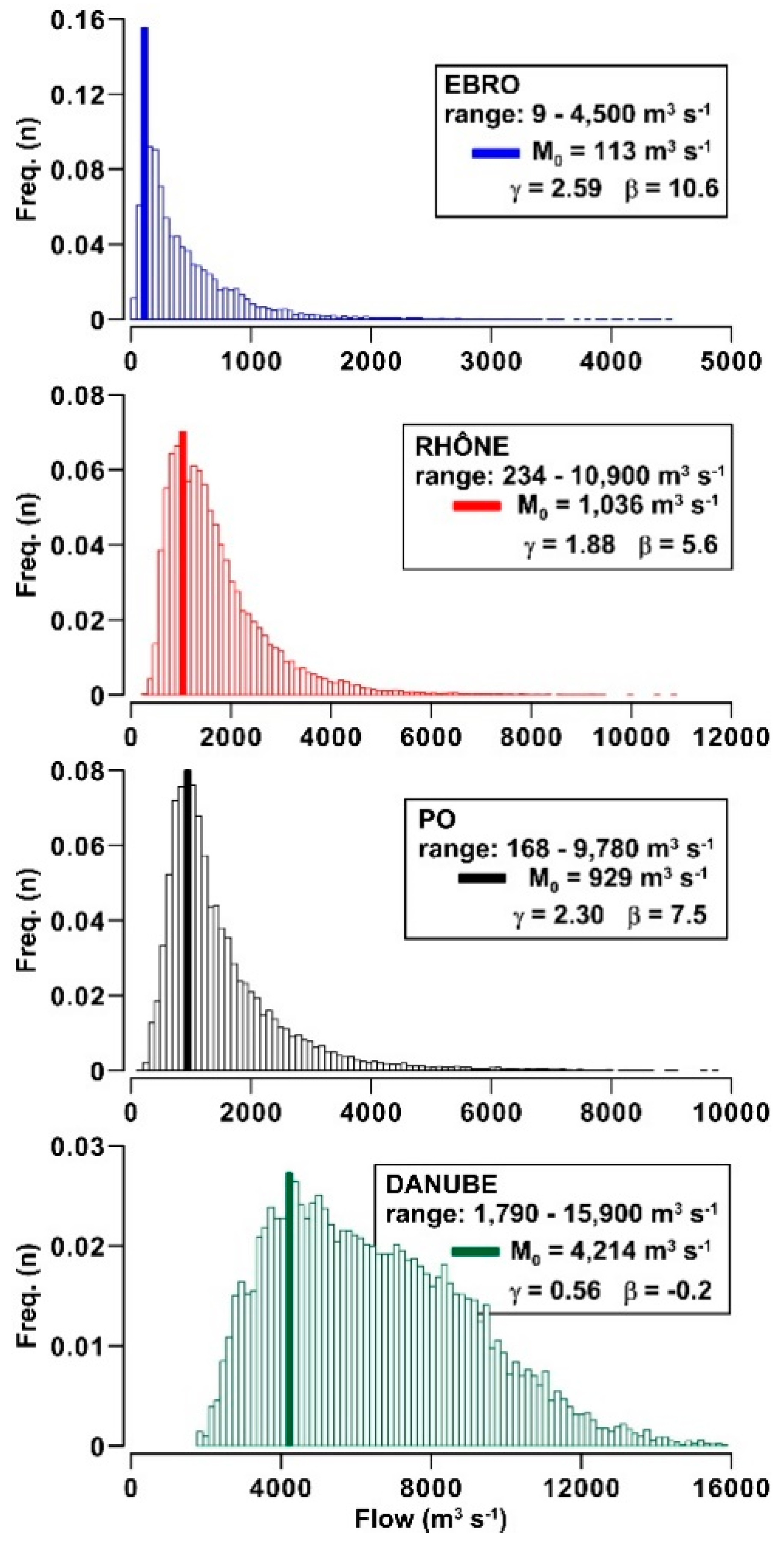
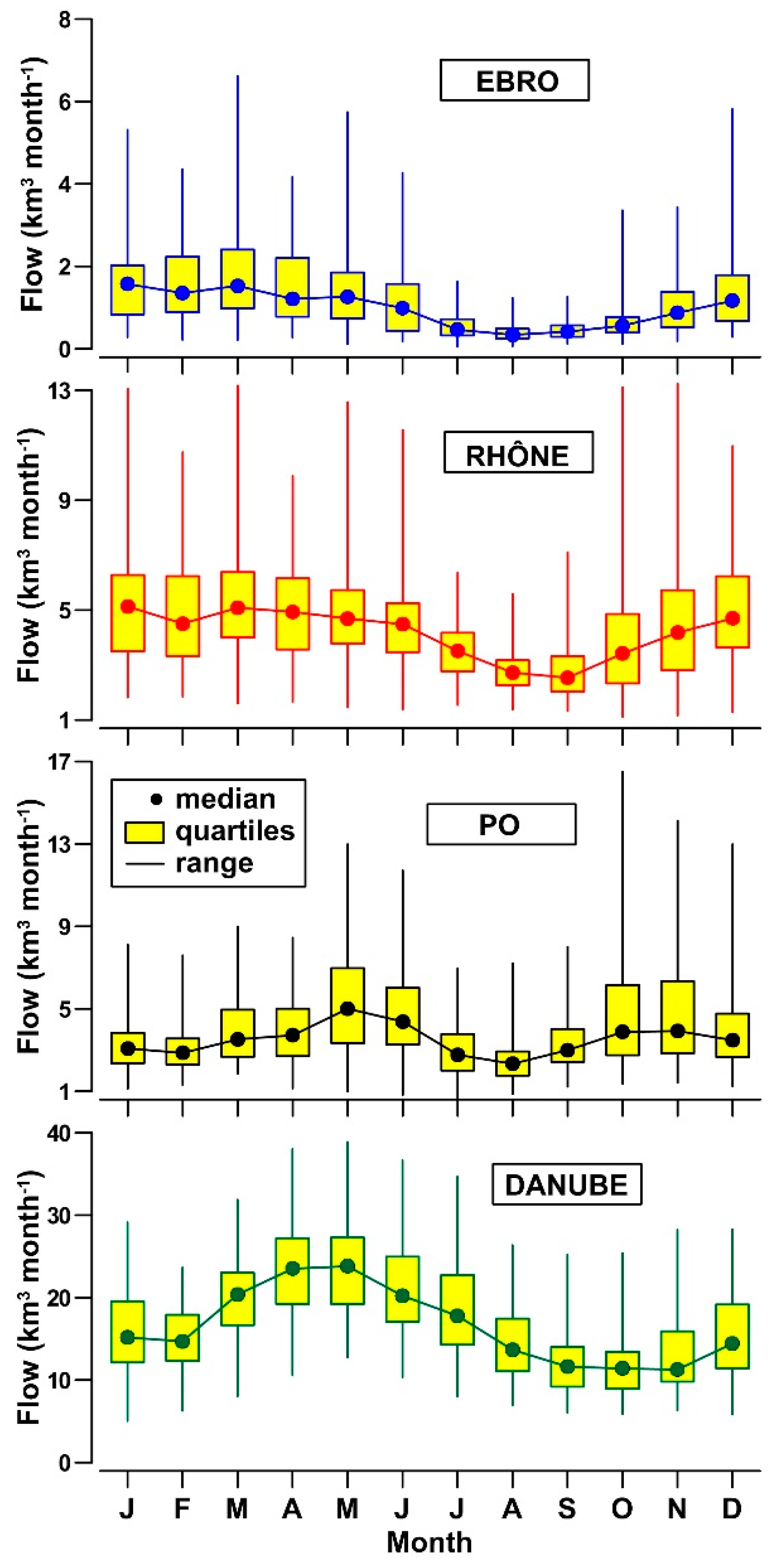
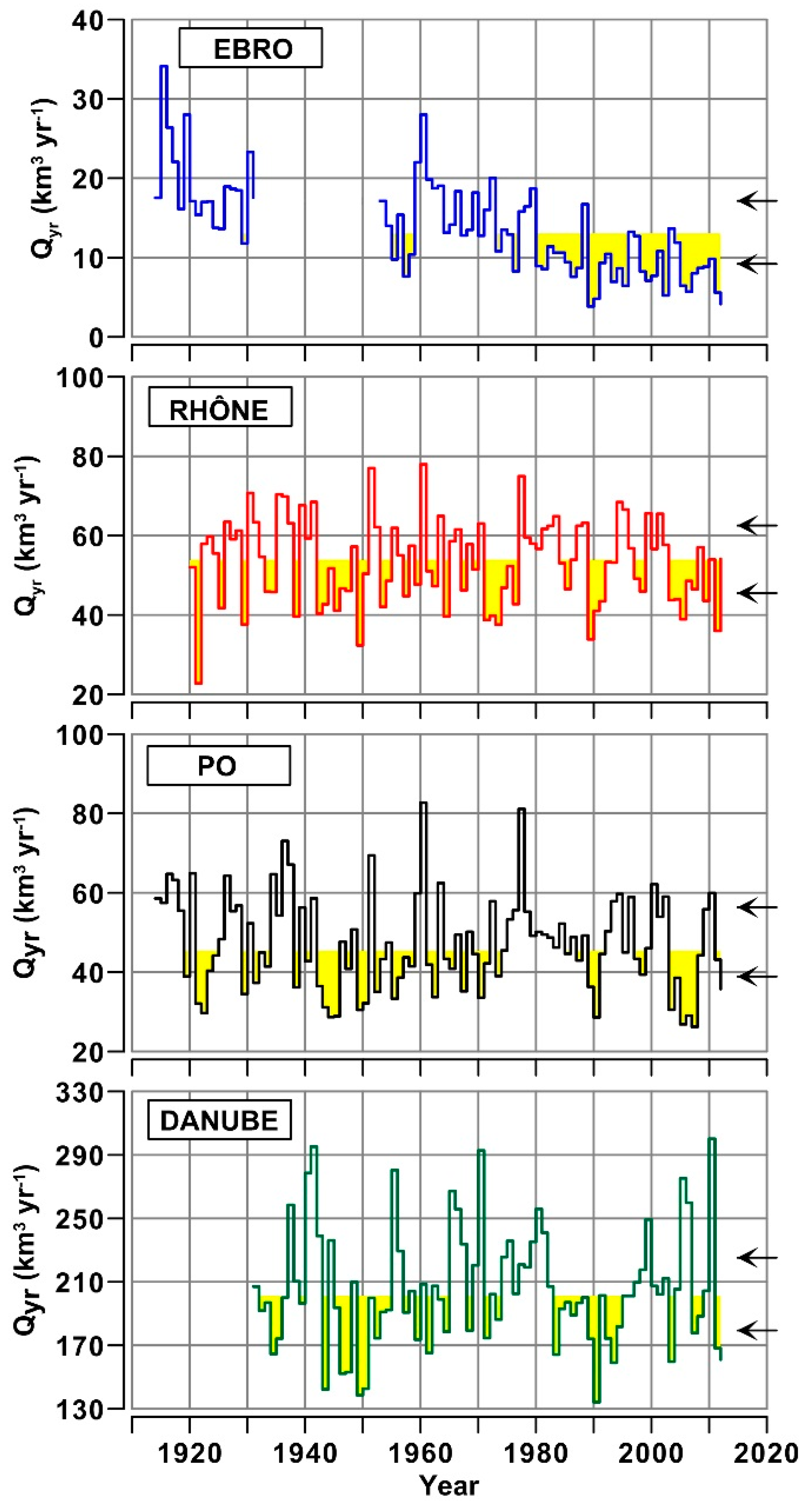
3.1. Daily to Multi-Decadal Variability of River Flows
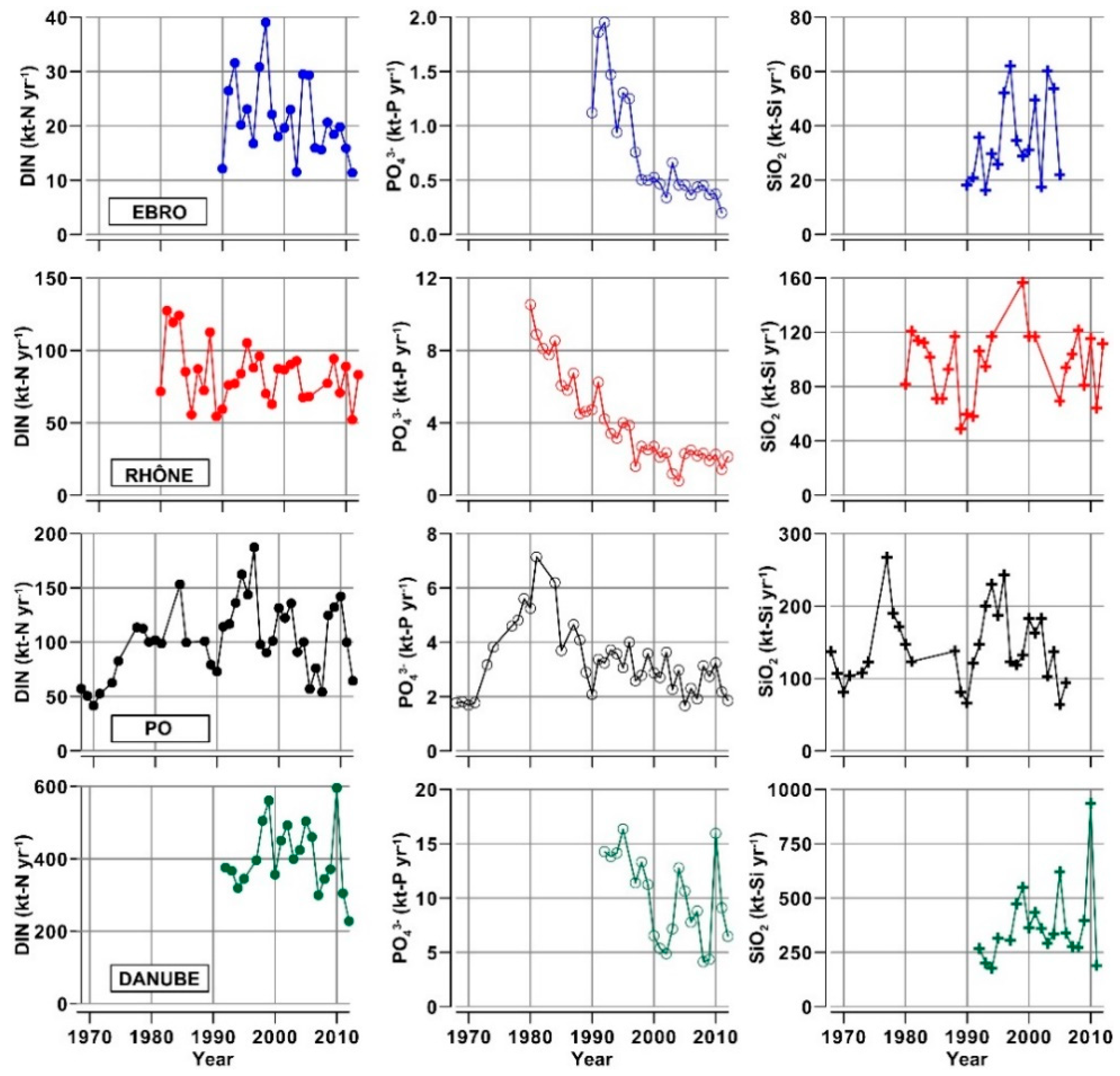
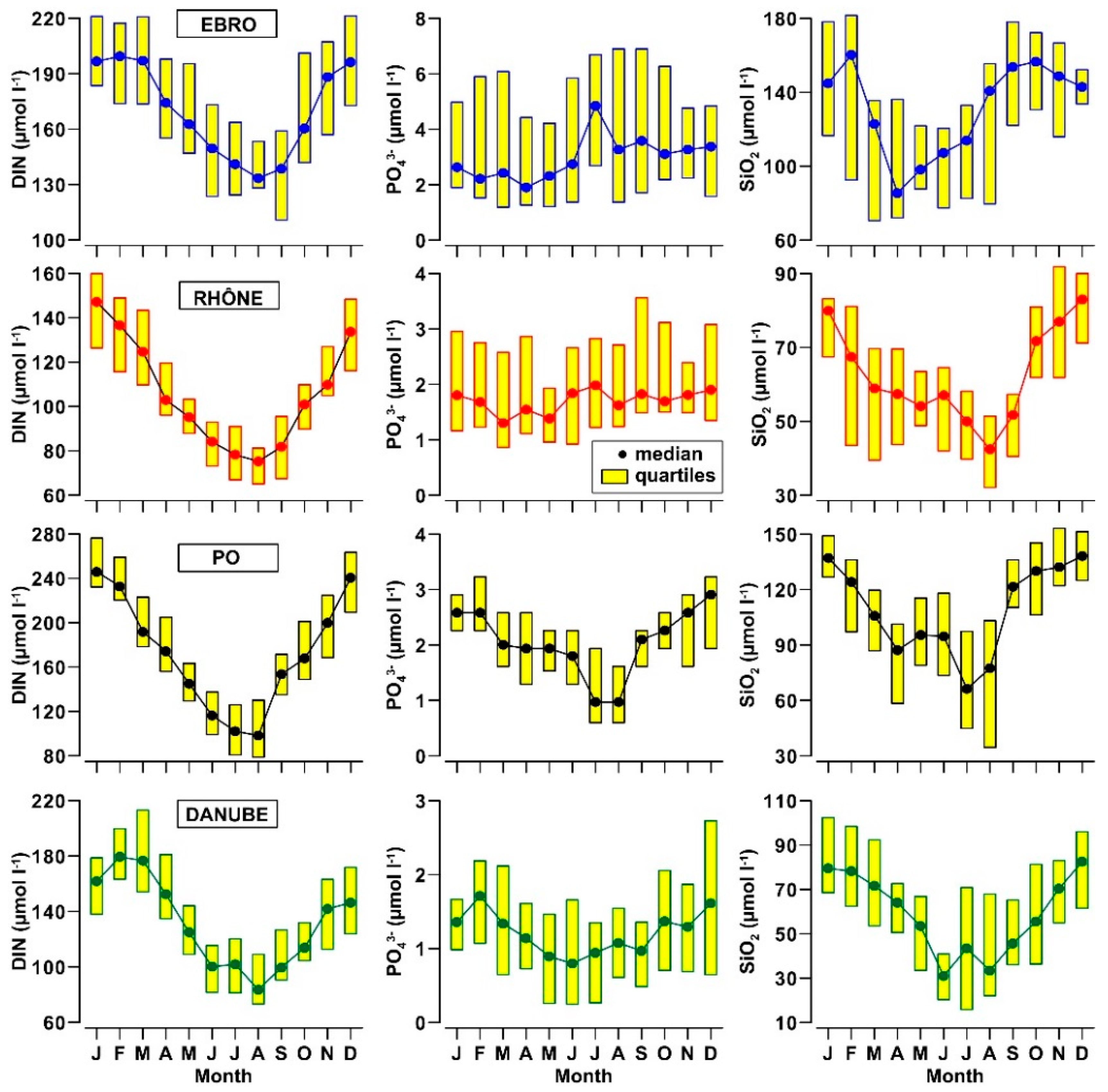
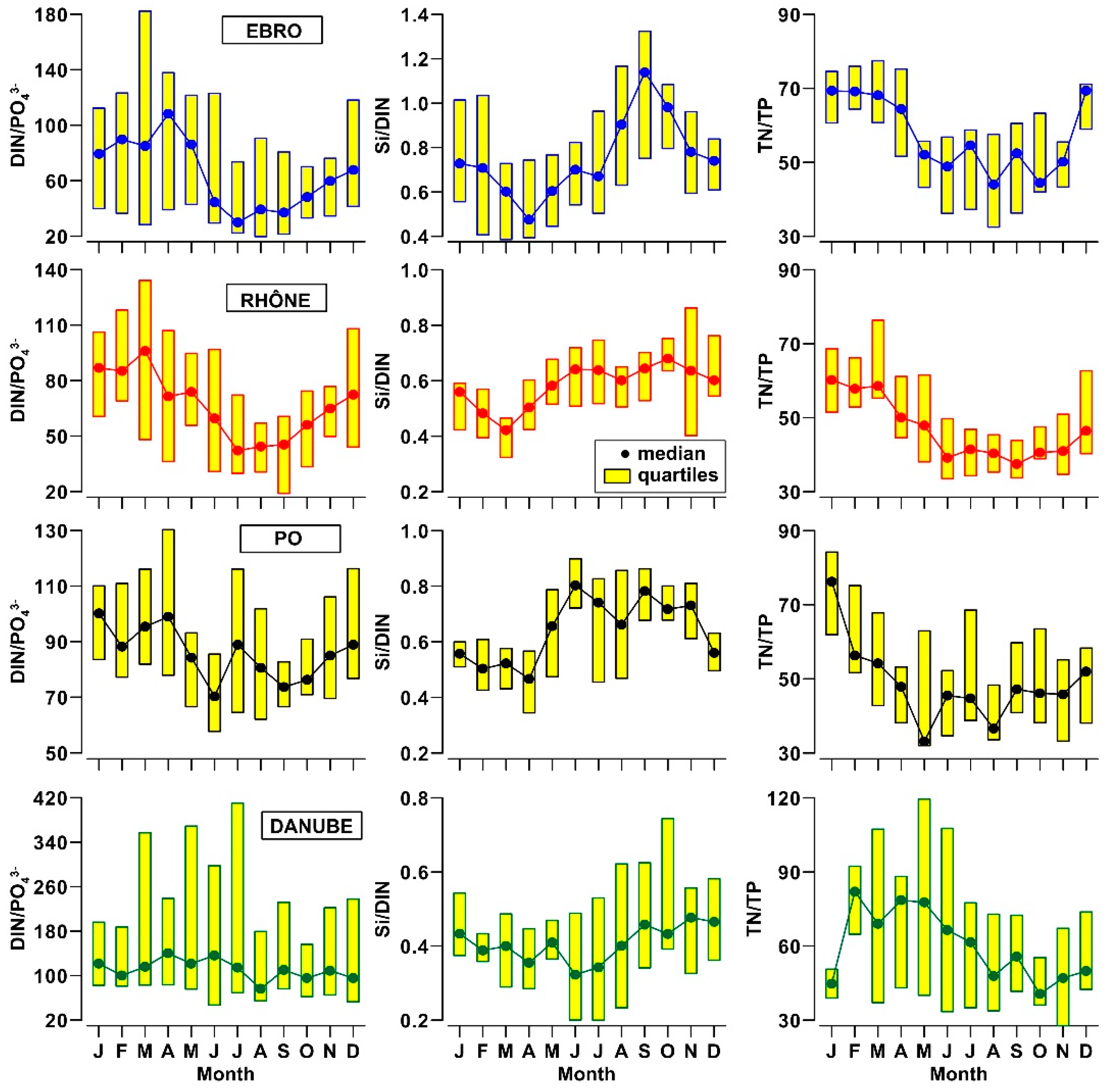
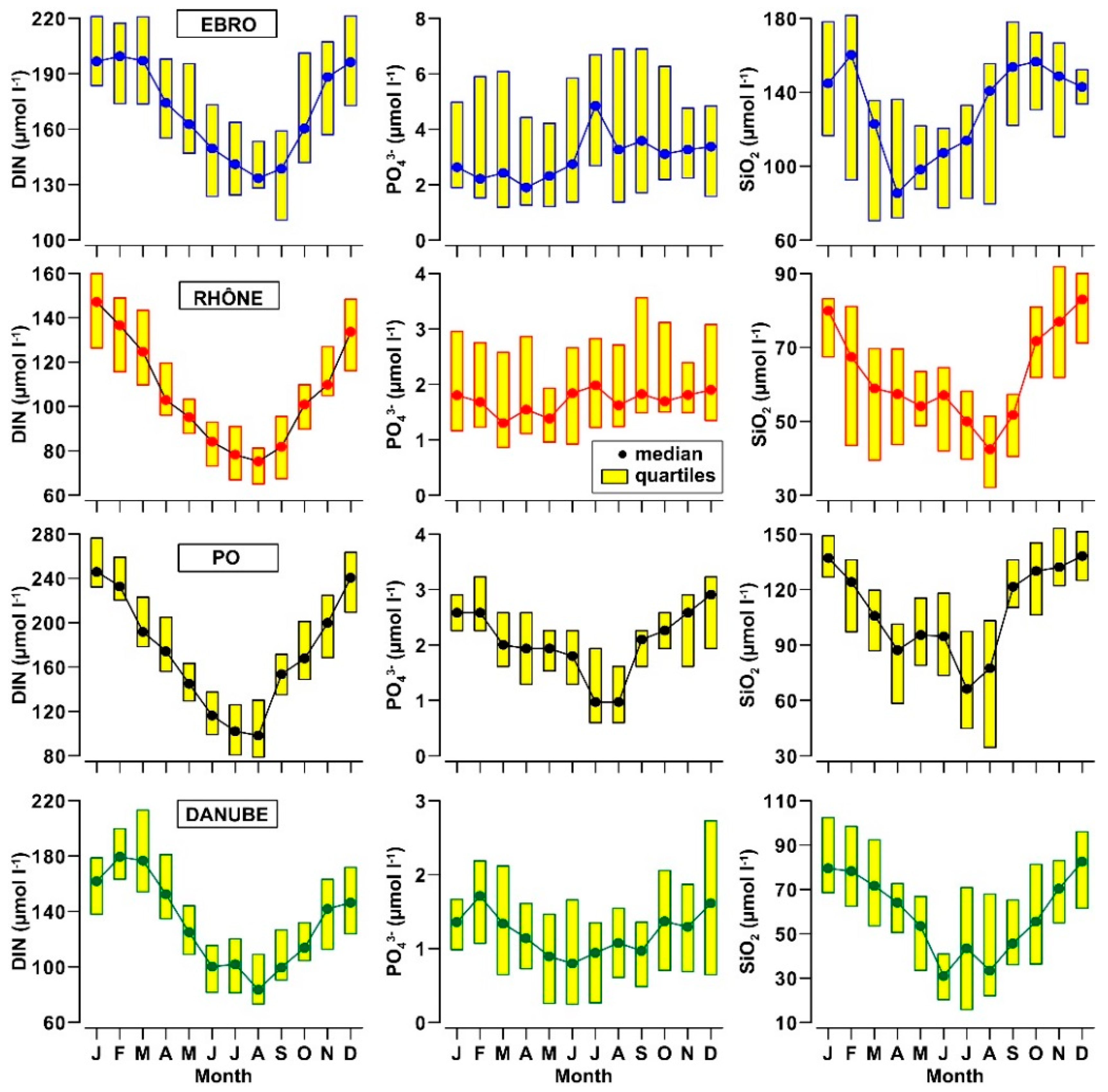
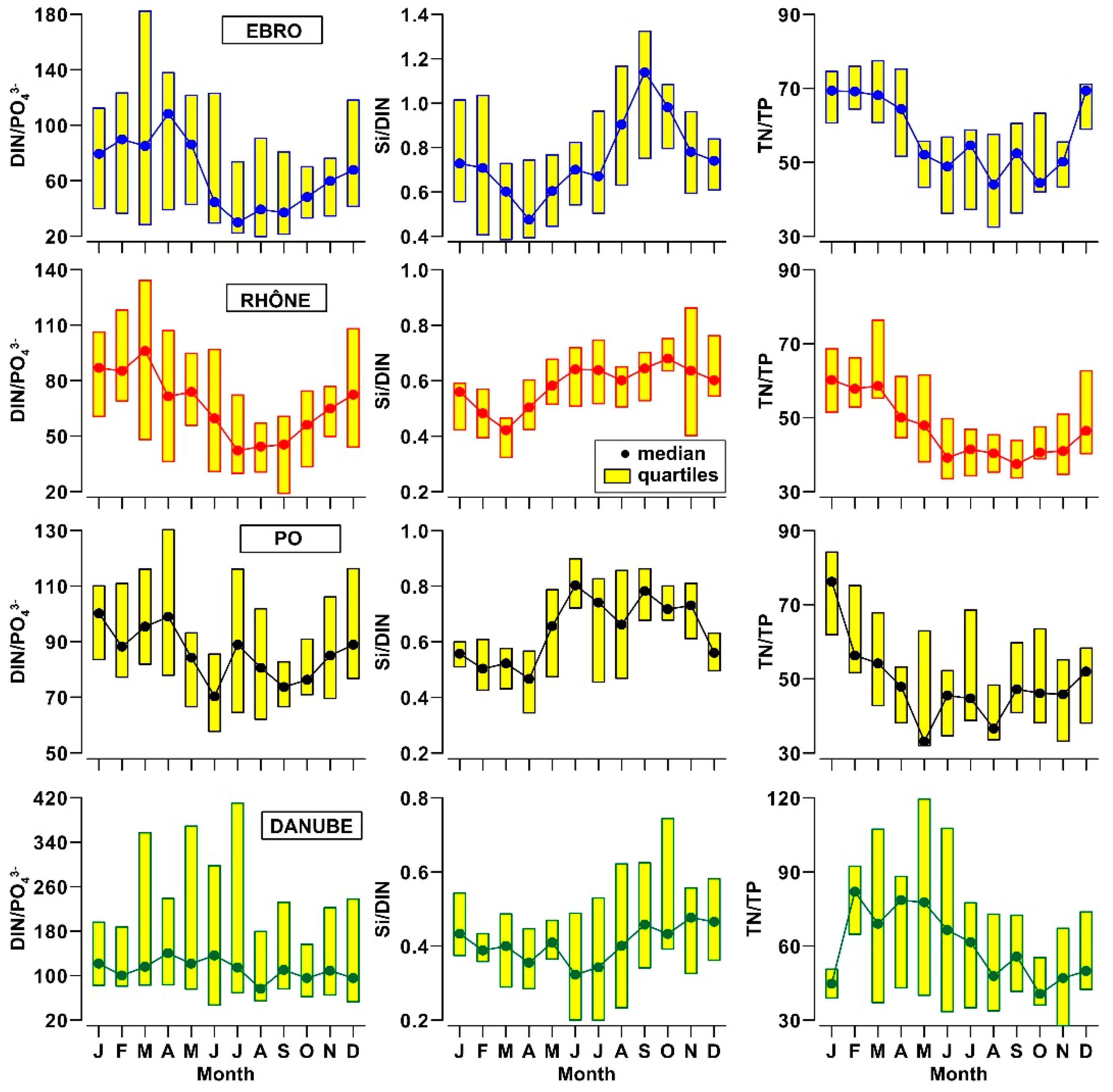
3.2. Nutrients and Organic Matter in River Waters
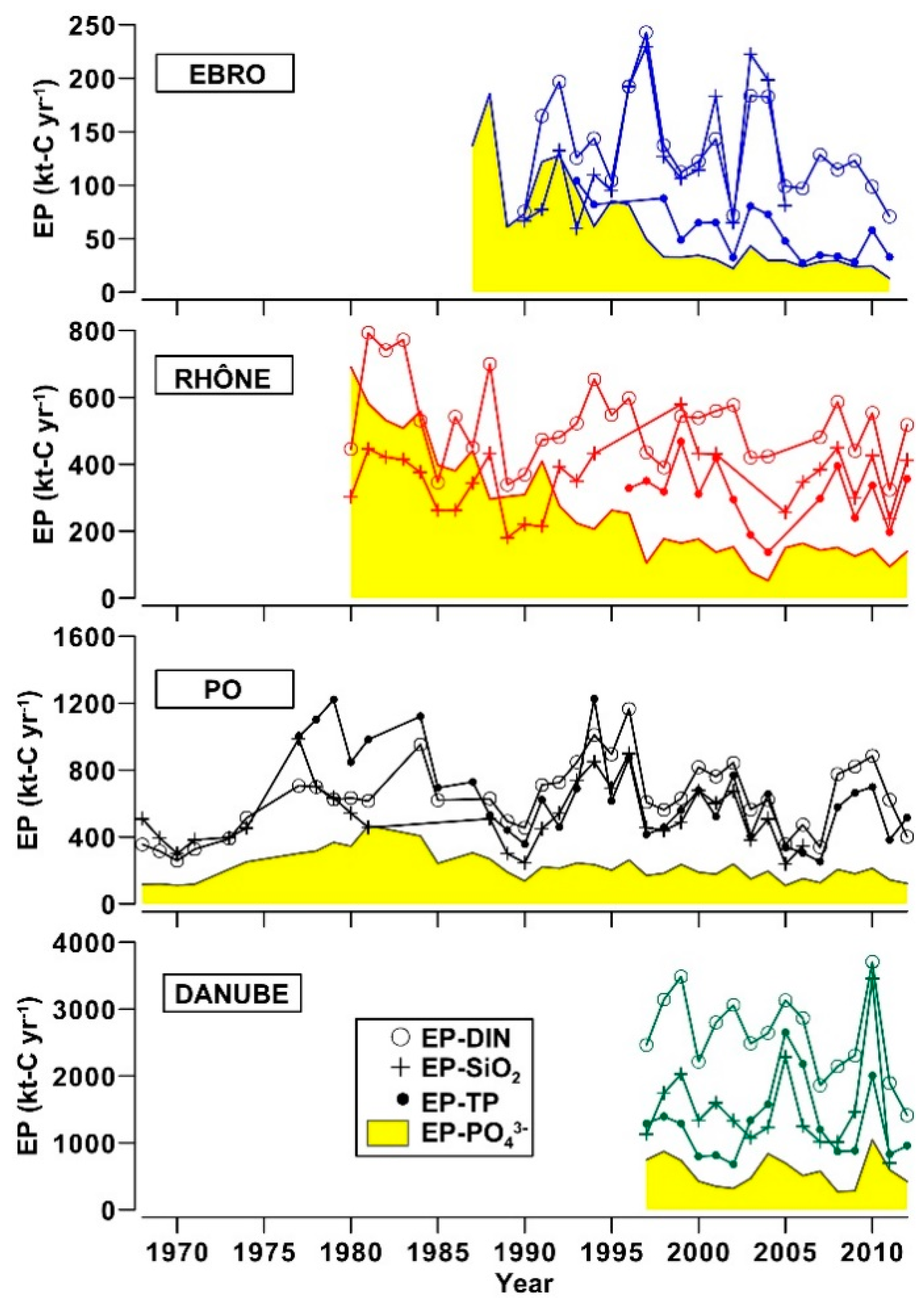
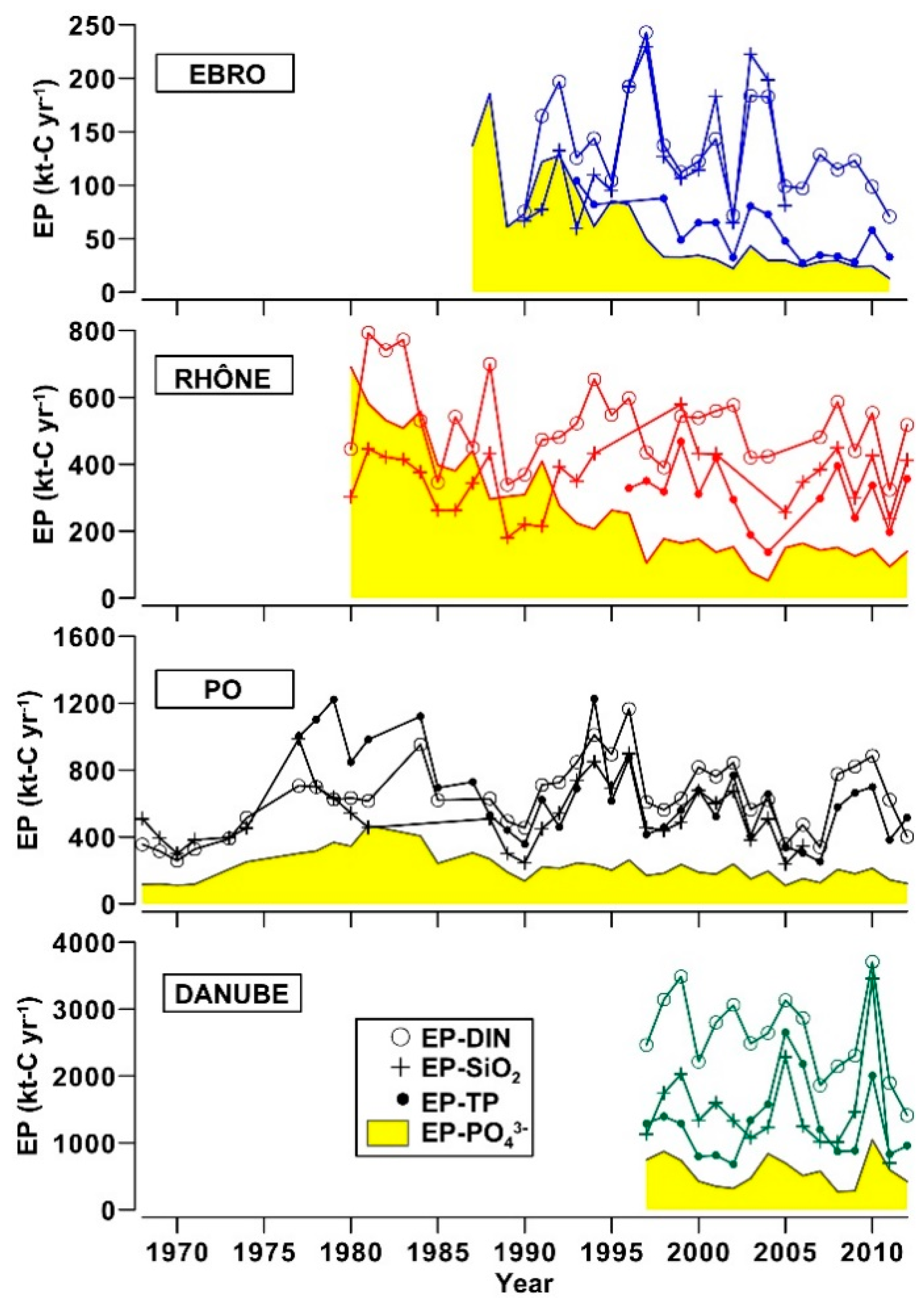
3.3. River Discharges of Biogenic Elements and Eutrophication Potential in Coastal Zones
4. Discussion
4.1. Seasonal to Decadal Trends of River Flows and Coastal Hydrology
4.2. Loading of Biogenic Elements and Impacts on Fluvial and Coastal Systems
5. Conclusions
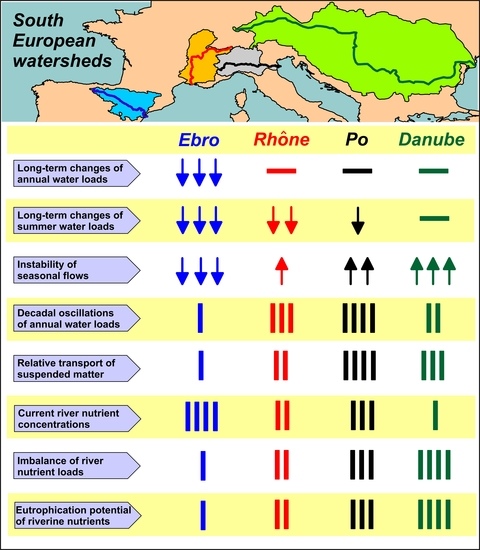
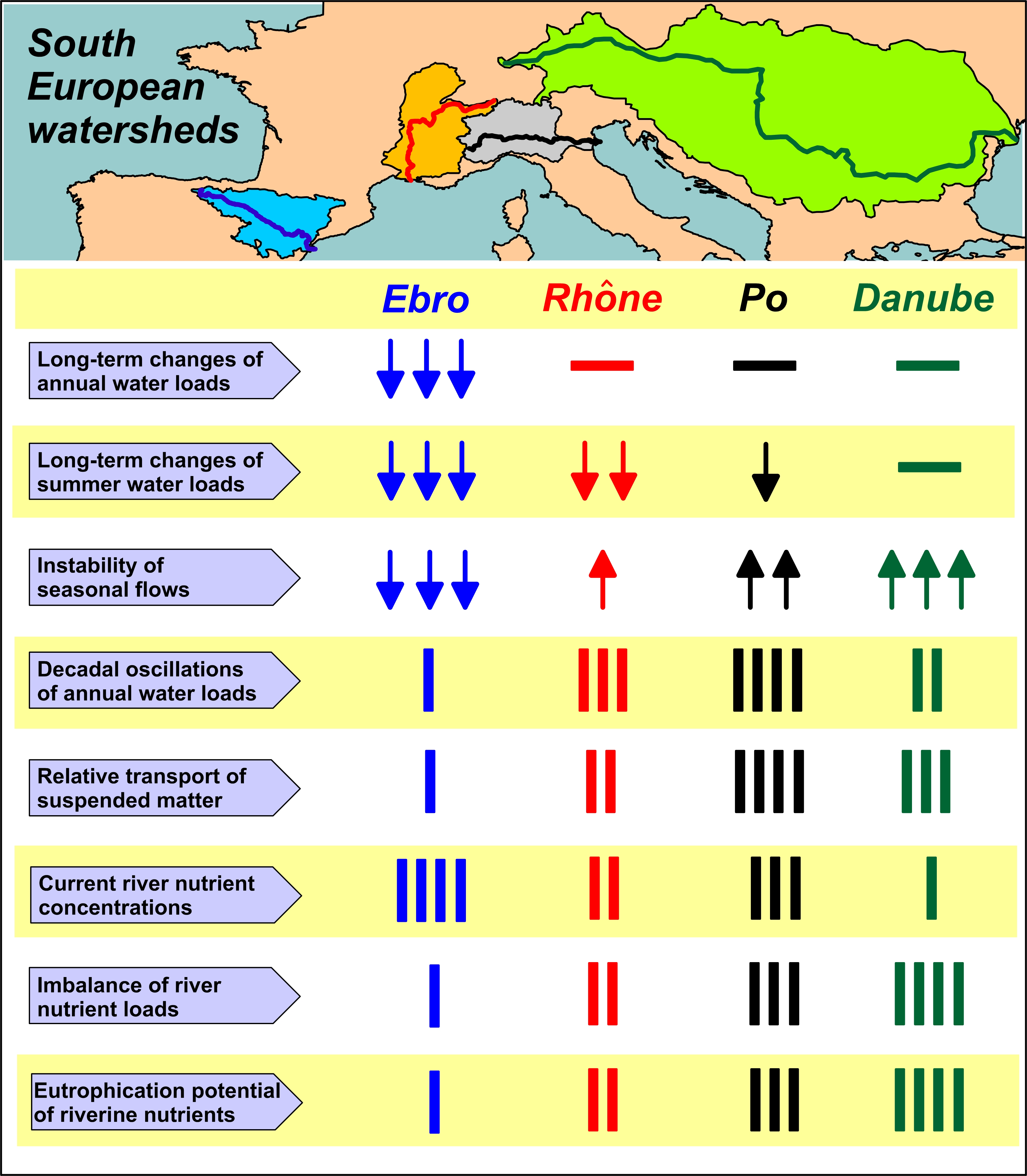
- Flow dynamics of Ebro, Rhône, Po and Danube rivers exhibits a different incidence of freshets and droughts and distinct annual cycles. This feature suggests the importance of regional climatic factors in these drainage basins, despite the widespread presence of flow regulation systems.
- Annual water discharges of the Ebro significantly decreased during the last century, whereas those Rhône, Po and Danube showed multi-decadal oscillations. For the Ebro, this difference is consistent with the rise of anthropogenic usage of freshwater in the drainage basins and with regional climate changes. For the other rivers, interannual variability of water discharge is still prevailing on long-term trends.
- The decrease of water discharge of the Ebro was concomitant to a reduction of flow variability in all the seasons. For the Rhône, Po and Danube, the decrease of discharges occurred mostly in summer, with a concomitant increase of flow variability that suggests a greater instability of climatic conditions in their regions.
- The concentrations of inorganic nutrients, TN and TP in the waters of Ebro and Po are about 50% higher than in those of Rhône and Danube. This finding suggests that the former two watersheds might be the most impacted ones by nutrient pollution, in a future scenario of reduced runoff, even in the presence of constant inputs due to agricultural, urban and industrial activities.
- The concentrations of DIN and SiO2 show a clear annual cycle in these rivers, with the lowest levels (−50%) being reached in spring and summer. This cycle changes seasonally quantity and composition of the nutrient pool delivered into the receiving coastal water bodies.
- The analysis of nutrient budgets indicated that these rivers have changed from a past condition characterized by large discharges of nutrients, with a rather balanced N:Si:P ratio, to overloads of DIN and SiO2 with respect to PO43−. This process has reduced the eutrophication of rivers, estuaries and coastal marine environments inducing, however, changes of ecological conditions that have to be further assessed.
- Phosphorus scarcity is a common feature of these river and coastal ecosystems, but its potential ability to limit primary production significantly reduces if organic phosphorus is considered at least partially available for the growth of phytoplankton. For this reason, the real bioavailability of riverine organic phosphorus for auto- and hetero-trophs should be better investigated as it could play a key role in the regulation of the productivity and structure of plankton communities.
- In the current post-eutrophic phase, the discharge of riverine TOC to the coastal zone is not negligible with respect to the eutrophication potential of river nutrients.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Ebro | Rhône | Po | Danube |
|---|---|---|---|---|
| km3 month−1 yr−1 | ||||
| Jan. | −0.013 *** | +0.011 | +0.001 | +0.045 |
| Feb. | −0.014 *** | +0.008 | +0.001 | +0.041 * |
| Mar. | −0.020 *** | −0.001 | −0.009 + | −0.005 |
| Apr. | −0.019 *** | −0.002 | −0.007 | +0.025 |
| May | −0.015 *** | −0.010 + | −0.001 | −0.015 |
| Jun. | −0.014 *** | −0.010 + | −0.015 * | −0.052 + |
| Jul. | −0.005 *** | −0.009 * | −0.016 ** | −0.040 |
| Aug. | −0.002 * | −0.007 ** | −0.008 * | −0.014 |
| Sep. | −0.003 *** | −0.004 | +0.001 | +0.011 |
| Oct. | −0.005 *** | −0.001 | −0.004 | +0.020 |
| Nov. | −0.010 *** | −0.001 | −0.005 | +0.006 |
| Dec. | −0.013 *** | +0.012 + | 0.000 | +0.007 |
| Month | Ebro | Rhône | Po | Danube |
|---|---|---|---|---|
| % yr−1 | ||||
| Jan. | −0.116 + | +0.065 | −0.065 | −0.123 * |
| Feb. | −0.211 *** | −0.002 | −0.054 | −0.135 |
| Mar. | −0.060 | +0.019 | −0.106 | +0.028 |
| Apr. | −0.019 | +0.082 | −0.021 | +0.074 + |
| May | +0.063 | +0.047 | +0.007 | +0.201 *** |
| Jun. | −0.040 | +0.105 + | +0.119 * | +0.123 * |
| Jul. | −0.420 *** | +0.279 *** | +0.134 ** | +0.127 * |
| Aug. | −0.417 *** | +0.128 * | −0.012 | +0.091 + |
| Sep. | −0.481 *** | +0.020 | −0.086 | +0.072 |
| Oct. | −0.251** | +0.006 | −0.042 | +0.004 |
| Nov. | −0.222 *** | +0.062 | −0.009 | −0.012 |
| Dec. | −0.165 * | +0.017 | +0.038 | +0.034 |
| Year | F-TSM kt yr−1 | F-NO3− kt-N yr−1 | F-NH4+ kt-N yr−1 | F-NO2− kt-N yr−1 | F-PO43− kt-P yr−1 | F-SiO2 kt-Si yr−1 | F-TOC kt-C yr−1 | F-TN kt-N yr−1 | F-TP kt-P yr−1 |
|---|---|---|---|---|---|---|---|---|---|
| Ebro | |||||||||
| Median | 70.91 | 19.17 | 0.50 | 0.19 | 0.50 | 30.37 | 29.28 | 26.12 | 0.81 |
| 1th Quartile | 51.70 | 15.74 | 0.27 | 0.15 | 0.44 | 21.64 | 25.70 | 21.85 | 0.51 |
| 3rd Quartile | 98.91 | 24.64 | 0.69 | 0.28 | 1.08 | 50.20 | 43.21 | 34.11 | 1.14 |
| Min. | 21.03 | 21.03 | 0.18 | 0.09 | 0.20 | 16.16 | 12.83 | 13.68 | 0.42 |
| Max. | 224.10 | 38.15 | 1.48 | 0.43 | 1.95 | 62.14 | 49.33 | 47.24 | 1.59 |
| Rhône | |||||||||
| Median | 2030.56 | 79.98 | 2.91 | 1.27 | 2.33 | 105.05 | 155.20 | 101.91 | 4.85 |
| 1th Quartile | 1262.63 | 65.71 | 2.39 | 0.98 | 2.11 | 78.11 | 143.92 | 89.27 | 4.08 |
| 3rd Quartile | 3972.30 | 83.93 | 3.43 | 1.53 | 3.28 | 116.62 | 229.02 | 109.66 | 5.39 |
| Min. | 405.14 | 49.67 | 1.16 | 0.44 | 0.79 | 58.02 | 124.25 | 62.79 | 2.09 |
| Max. | 7998.60 | 99.54 | 4.59 | 2.52 | 6.24 | 156.88 | 279.43 | 115.80 | 7.13 |
| Po | |||||||||
| Median | 5546.90 | 102.11 | 3.99 | 1.44 | 2.88 | 137.49 | 246.42 | 156.80 | 8.53 |
| 1th Quartile | 3434.51 | 85.43 | 3.05 | 1.16 | 2.28 | 118.75 | 229.18 | 111.37 | 6.27 |
| 3rd Quartile | 7771.78 | 126.30 | 6.08 | 1.85 | 3.30 | 183.28 | 299.69 | 227.11 | 10.25 |
| Min. | 1030.47 | 51.42 | 1.60 | 0.54 | 1.67 | 64.39 | 225.14 | 94.19 | 3.83 |
| Max. | 16,292.57 | 179.19 | 9.69 | 2.53 | 4.00 | 243.46 | 366.53 | 295.16 | 18.71 |
| Danube | |||||||||
| Median | 6565.15 | 338.77 | 62.72 | 7.95 | 8.28 | 360.13 | 1139.75 | 446.30 | 18.96 |
| 1th Quartile | 4820.07 | 297.08 | 36.09 | 5.67 | 6.19 | 298.29 | 968.46 | 374.27 | 13.17 |
| 3rd Quartile | 10,478.27 | 406.26 | 75.93 | 8.92 | 11.27 | 452.54 | 1311.04 | 587.20 | 21.95 |
| Min. | 2645.97 | 188.40 | 26.08 | 2.14 | 4.13 | 189.11 | 797.17 | 270.00 | 10.39 |
| Max. | 19,077.89 | 535.22 | 89.79 | 12.05 | 15.96 | 935.85 | 1482.33 | 714.05 | 40.47 |
| Parameter | Ebro | Rhône | Po | Danube |
|---|---|---|---|---|
| kt yr−2 | ||||
| F-TSM | −1.2 + | −10.6 | −171 | +190 |
| F-NO3− | −0.23 | −0.11 | −0.35 | −8.31 |
| F-NH4+ | +0.01 | −0.06 | −0.28 *** | −3.58 ** |
| F-NO2− | −0.01 * | −0.04 *** | −0.06 *** | −0.18 |
| F-DIN | −0.36 + | −0.35 | −0.81 | −11.1 |
| F-PO43− | −0.06 *** | −0.22 *** | −0.06 ** | −0.18 |
| F-SiO2 | +1.3 | +0.04 | −1.06 | −7.04 |
| F-TOC | −1.36 + | - | - | - |
| F-TN | −0.22 | −0.52 | −1.07 | −32.5 |
| F-TP | −0.05 ** | −0.09 | −0.12 + | 0.00 |
| F-ON | +0.01 | −1.13 * | +0.09 | −4.30 |
| F-OP | −0.01 | −0.02 | −0.04 | +0.08 |
References
- Smith, V.H. Eutrophication of freshwater and coastal marine ecosystems. A global problem. Environ. Sci. Pollut. Res. 2003, 10, 1–14. [Google Scholar] [CrossRef]
- Garnier, J.; Beusen, A.; Thieu, V.; Billen, G.; Bouwman, L. N:P:Si nutrient export ratios and ecological consequences in coastal seas evaluated by the ICEP approach. Glob. Biogeochem. Cycles 2010, 24, GB0A05. [Google Scholar] [CrossRef]
- Howarth, R.; Chan, F.; Conley, D.J.; Garnier, J.; Doney, S.C.; Marino, R.; Billen, G. Coupled biogeochemical cycles: Eutrophication and hypoxia in temperate estuaries and coastal marine ecosystems. Front. Ecol. Environ. 2011, 9, 18–26. [Google Scholar] [CrossRef]
- Ibáñez, C.; Alcaraz, C.; Caiola, N.; Rovira, A.; Trobajo, R.; Alonso, M.; Duran, C.; Jiménez, P.J.; Munné, A.; Prat, N. Regime shift from phytoplankton to macrophyte dominance in a large river: Top-down versus bottom-up effects. Sci. Total Environ. 2012, 416, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Poulter, B.; Sardans, J.; Ciais, P.; van der Velde, M.; Bopp, L.; Boucher, O.; Godderis, Y.; Hinsinger, P.; Llusia, J.; et al. Human-induced nitrogen–phosphorus imbalances alter natural and managed ecosystems across the globe. Nat. Commun. 2013, 4, 2934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollenweider, R.A.; Marchetti, R.; Viviani, R. Marine Coastal Eutrophication. Proceedings of an International Conference, Bologna, Italy, 21–24 March 1990; Elsevier: Amsterdam, The Netherlands; London, UK; New York, NY, USA; Tokyo, Japan, 1992; p. 1310. ISBN 978-0-444-89990-3. [Google Scholar]
- Howarth, R.; Anderson, D.; Cloern, J.; Elfring, C.; Hopkinson, C.; Lapointe, B.; Malone, T.; Marcus, N.; McGlathery, K.; Sharpley, A.; et al. Nutrient Pollution of Coastal Rivers, Bays, and Seas; Issues in Ecology, Ecological Society of America: Washington, DC, USA, 2000; Volume 7, p. 17. ISSN 1092-8987. [Google Scholar]
- Yunev, O.A.; Carstensen, J.; Moncheva, S.; Khaliulin, A.; Ærtebjerg, G.; Nixon, S. Nutrient and phytoplankton trends on the western Black Sea shelf in response to cultural eutrophication and climate changes. Estuar. Coast. Shelf Sci. 2007, 74, 63–76. [Google Scholar] [CrossRef]
- Rabouille, C.; Conley, D.J.; Dai, M.H.; Cai, W.-J.; Chen, C.T.A.; Lansard, B.; Green, R.; Yin, K.; Harrison, P.J.; Dagg, M.; et al. Comparison of hypoxia among four river-dominated ocean margin: The Changjiang (Yangtze), Mississippi, Pearl, and Rhône rivers. Cont. Shelf Res. 2008, 28, 1527–1537. [Google Scholar] [CrossRef]
- Oguz, T.; Velikova, V. Abrupt transition of the northwestern Black Sea shelf ecosystem from a eutrophic to an alternative pristine state. Mar. Ecol. Prog. Ser. 2010, 405, 231–242. [Google Scholar] [CrossRef] [Green Version]
- Djakovac, T.; Supić, N.; Bernardi-Aubry, F.; Degobbis, D.; Giani, M. Mechanisms of hypoxia frequency changes in the northern Adriatic Sea during the period 1972–2012. J. Mar. Syst. 2014, 141, 179–189. [Google Scholar] [CrossRef]
- Stachowitsch, M. Coastal hypoxia and anoxia: A multi-tiered holistic approach. Biogeosciences 2014, 11, 2281–2285. [Google Scholar] [CrossRef]
- Bianchi, T.S. The role of terrestrially derived organic carbon in the coastal ocean: A changing paradigm and the priming effect. Proc. Natl. Acad. Sci. USA 2011, 108, 19473–19481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asmala, E.; Autio, R.; Kaartokallio, H.; Stedmon, C.A.; Thomas, D.N. Processing of humic-rich riverine dissolved organic matter by estuarine bacteria: Effects of predegradation and inorganic nutrients. Aquat. Sci. 2014, 76, 451–463. [Google Scholar] [CrossRef]
- Durand, N.; Fiandrino, A.; Fraunié, P.; Ouillon, S.; Forget, P.; Naudin, J.J. Suspended matter dispersion in the Ebro ROFI: An integrated approach. Cont. Shelf Res. 2002, 22, 267–284. [Google Scholar] [CrossRef]
- Berlinsky, N.; Bogatova, Y.; Garkavaya, G. Estuary of the Danube. In Handbook of Environmental Chemistry; Water Pollution, 5 (PART H); Wangersky, P.J., Ed.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 5, pp. 233–264. ISBN 978-3-540-00270-3. [Google Scholar]
- Syvitski, J.P.M.; Kettner, A.J.; Overeem, I.; Hutton, E.W.H.; Hannon, M.T.; Brakenridge, G.R.; Day, J.; Vörösmarty, C.; Saito, Y.; Giosan, L.; et al. Sinking deltas due to human activities. Nat. Geosci. 2009, 2, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Markaki, Z.; Loÿe-Pilot, M.D.; Violaki, K.; Benyahya, L.; Mihalopoulos, N. Variability of atmospheric deposition of dissolved nitrogen and phosphorus in the Mediterranean and possible link to the anomalous seawater N/P ratio. Mar. Chem. 2010, 120, 187–194. [Google Scholar] [CrossRef]
- The MerMex Group. Marine ecosystems’ responses to climatic and anthropogenic forcings in the Mediterranean. Prog. Oceanogr. 2011, 91, 97–166. [Google Scholar] [CrossRef]
- Giani, M.; Djakovac, T.; Degobbis, D.; Cozzi, S.; Solidoro, C.; Fonda Umani, S. Recent changes in the marine ecosystems of the northern Adriatic Sea. Estuar. Coast. Shelf Sci. 2012, 115, 1–13. [Google Scholar] [CrossRef]
- Ludwig, W.; Dumont, E.; Meybeck, M.; Heussner, S. River discharges of water and nutrients to the Mediterranean and Black Sea: Major drivers for ecosystem changes during past and future decades? Prog. Oceanogr. 2009, 80, 199–217. [Google Scholar] [CrossRef]
- Huertas, I.E.; Ríos, A.F.; García-Lafuente, J.; Navarro, G.; Makaoui, A.; Sánchez-Román, A.; Rodriguez-Galvez, S.; Orbi, A.; Ruíz, J.; Pérez, F.F. Atlantic forcing of the Mediterranean oligotrophy. Glob. Biogeochem. Cycles 2012, 26, GB2022. [Google Scholar] [CrossRef]
- Kempe, S.; Pettine, M.; Cauwet, G. Biogeochemistry of European Rivers. In Biogeochemistry of Major World Rivers; SCOPE Report 42; Degens, E.T., Kempe, S., Richey, J.E., Eds.; ICSU, UNEP, John Wiley & Sons Ltd.: Chichester, UK, 1991; pp. 169–211. ISSN 1052-7613. [Google Scholar]
- Grizzetti, B.; Bouraoui, F.; Aloe, A. Changes of nitrogen and phosphorus loads to European seas. Glob. Chang. Biol. 2012, 18, 769–782. [Google Scholar] [CrossRef]
- Romero, E.; Garnier, J.; Lassaletta, L.; Billen, G.; Le Gendre, R.; Riou, P.; Cugier, P. Large-scale patterns of river inputs in southwestern Europe: Seasonal and interannual variations and potential eutrophication effects at the coastal zone. Biogeochemistry 2013, 113, 481–505. [Google Scholar] [CrossRef]
- DANUBS Final Report. Nutrient Management in the Danube Basin and Its Impact on the Black Sea. EVK1-CT-2000-00051. 2005, p. 78. Available online: http://danubs.tuwien.ac.at (accessed on 12 October 2017).
- Ibáñez, C.; Prat, N.; Duran, C.; Pardos, M.; Munné, A.; Andreu, R.; Caiola, N.; Cid, N.; Hampel, H.; Sánchez, R.; Trobajo, R. Changes in dissolved nutrients in the lower Ebro river: Causes and consequences. Limnetica 2008, 27, 131–142. [Google Scholar]
- Ollivier, P.; Radakovitch, O.; Hamelin, B. Major and trace element partition and fluxes in the Rhône River. Chem. Geol. 2011, 285, 15–31. [Google Scholar] [CrossRef]
- Viaroli, P.; Soana, E.; Pecora, S.; Laini, A.; Naldi, M.; Fano, E.A.; Nizzoli, D. Space and time variations of watershed N and P budgets and their relationships with reactive N and P loadings in a heavily impacted river basin (Po river, Northern Italy). Sci. Total Environ. 2018, 639, 1574–1587. [Google Scholar] [CrossRef] [PubMed]
- Cruzado, A.; Velásquez, Z.; Pérez, M.D.C.; Bahamón, N.; Grimaldo, N.S.; Ridolfi, F. Nutrient fluxes from the Ebro River and subsequent across-shelf dispersion. Cont. Shelf Res. 2002, 22, 349–360. [Google Scholar] [CrossRef]
- Alvisi, F.; Cozzi, S. Seasonal dynamics and long-term trends of hypoxia in the coastal zone of Emilia Romagna (NW Adriatic Sea, Italy). Sci. Total Environ. 2016, 541, 1448–1462. [Google Scholar] [CrossRef] [PubMed]
- Kovats, R.S.; Valentini, R.; Bouwer, L.M.; Georgopoulou, E.; Jacob, D.; Martin, E.; Rounsevell, M.; Soussana, J.-F. Europe. In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of IPCC; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 1267–1326. ISBN 978-1-107-05816-3. [Google Scholar]
- Díaz, F.; Naudin, J.J.; Courties, C.; Rimmelin, P.; Oriol, L. Biogeochemical and ecological functioning of the low-salinity water lenses in the region of the Rhone River freshwater influence, NW Mediterranean Sea. Cont. Shelf Res. 2008, 28, 1511–1526. [Google Scholar] [CrossRef]
- Cozzi, S.; Giani, M. River water and nutrient discharges in the Northern Adriatic Sea: Current importance and long-term changes. Cont. Shelf Res. 2011, 31, 1881–1893. [Google Scholar] [CrossRef]
- Tuǧrul, S.; Murray, J.W.; Friederich, G.E.; Salihoǧlu, T. Spatial and temporal variability in the chemical properties of the oxic and suboxic layers of the Black Sea. J. Mar. Syst. 2014, 135, 29–43. [Google Scholar] [CrossRef]
- Lutz, S.R.; Mallucci, S.; Diamantini, E.; Majone, B.; Bellin, A.; Merz, R. Hydroclimatic and water quality trends across three Mediterranean river basins. Sci. Total Environ. 2016, 571, 1392–1406. [Google Scholar] [CrossRef]
- Zampieri, M.; Scoccimarro, E.; Gualdi, S.; Navarra, A. Observed shift towards earlier spring discharge in the main Alpine rivers. Sci. Total Environ. 2015, 503–504, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, C.; Prat, N. The environmental impact of the Spanish National Hydrological Plan on the lower Ebro River and Delta. Int. J. Water Resour. Dev. 2003, 19, 485–500. [Google Scholar] [CrossRef]
- Falco, S.; Niencheski, L.F.; Rodilla, M.; Romero, I.; González del Río, J.; Sierra, J.P.; Mösso, C. Nutrient flux and budget in the Ebro Estuary. Estuar. Coast. Shelf Sci. 2010, 87, 92–102. [Google Scholar] [CrossRef]
- Torrecilla, N.J.; Galve, J.P.; Zaera, L.G.; Retamar, J.F.; Álvarez, A.N.A. Nutrient sources and dynamics in a Mediterranean fluvial regime (Ebro River, NE Spain) and their implications for water management. J. Hydrol. 2005, 304, 166–182. [Google Scholar] [CrossRef]
- Tornés, E.; Pérez, M.C.; Durán, C.; Sabater, S. Reservoirs override seasonal variability of phytoplankton communities in a regulated Mediterranean river. Sci. Total Environ. 2014, 475, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Higueras, M.; Kerhervé, P.; Sanchez-Vidal, A.; Calafat, A.; Ludwig, W.; Verdoit-Jarraya, M.; Heussner, S.; Canals, M. Biogeochemical characterization of the riverine particulate organic matter transferred to the NW Mediterranean Sea. Biogeosciences 2014, 11, 157–172. [Google Scholar] [CrossRef] [Green Version]
- Denant, V.; Saliot, A. Seasonal variations of nutrients (NO3−, NO2−, NH4+, PO43− and Si(OH)4) and suspended matter in the Rhône delta, France. Oceanol. Acta 1990, 13, 47–52. [Google Scholar]
- Moutin, T.; Raimbault, P.; Golterman, H.L.; Coste, B. The input of nutrients by the Rhône River into the Mediterranean Sea: Recent observations and comparison with earlier data. Hydrobiologia 1998, 373/374, 237–246. [Google Scholar] [CrossRef]
- Fruget, J.-F.; Centofanti, M.; Dessaix, J.; Olivier, J.-M.; Druart, J.-C.; Martinez, P.-J. Temporal and spatial dynamics in large rivers: Example of a long-term monitoring of the middle Rhône River. Ann. Limnol. 2001, 37, 237–251. [Google Scholar] [CrossRef]
- Basin Authority of Po River. Characteristics of Po River Basin and Preliminary Assessment of the Environmental Impact of Human Activities on Hydrological Resources. Parma, Italy, 2006; p. 643, [Italian]. Available online: http://www.adbpo.gov.it/it/distretto-del-po/alcuni-dati (accessed on 29 January 2017).
- Copetti, D.; Carniato, L.; Crise, A.; Guyennon, N.; Palmeri, L.; Pisacane, G.; Struglia, M.V.; Tartari, G. Impacts of Climate Change on Water Quality. In Regional Assessment of Climate Change in the Mediterranean; Advances in Global Change Research; Navarra, A., Tubiana, L., Eds.; Springer: Dordrecht, The Netherlands, 2013; Volume 50, pp. 307–332. ISBN 978-94-007-5780-6. [Google Scholar]
- Tesi, T.; Miserocchi, S.; Acri, F.; Langone, L.; Boldrin, A.; Hatten, J.A.; Albertazzi, S. Flood-driven transport of sediment, particulate organic matter, and nutrients from the Po River watershed to the Mediterranean Sea. J. Hydrol. 2013, 498, 144–152. [Google Scholar] [CrossRef]
- Humborg, C.; Ittekkot, V.; Cociasu, A.; Bodungen, B.V. Effect of Danube River dam on Black Sea biogeochemistry and ecosystem structure. Nature 1997, 386, 385–388. [Google Scholar] [CrossRef]
- Rice, E.W.; Baird, R.B.; Eaton, A.D.; Clesceri, L.S. Standard Methods for the Examination of Water and Wastewater, 22nd ed.; American Public Health Association: Washington, DC, USA, 2012; p. 1360. ISBN 978-087553-013-0. [Google Scholar]
- Grasshoff, K.; Kremling, K.; Ehrhardt, M. Methods of Seawater Analysis. Third Completely Revised and Extended Edition; Wiley-VCH Verlag GmbH: Weinheim, Germany, 1999; p. 632. ISBN 978-3-527-61399-1. [Google Scholar]
- Ott, W.R. Environmental Statistics and Data Analysis; Lewis Publishers: Boca Raton, FL, USA; London, UK; New York, NY, USA; Washington, DC, USA, 1994; p. 336. ISBN 9780873718486. [Google Scholar]
- Chen, Z.; Grasby, S.E. Impact of decadal and century-scale oscillations on hydroclimate trend analyses. J. Hydrol. 2009, 365, 122–133. [Google Scholar] [CrossRef]
- Rodionov, S.N. A sequential method of detecting abrupt changes in the correlation coefficient and its application to Bering Sea Climate. Climate 2015, 3, 474–491. [Google Scholar] [CrossRef]
- Pujo-Pay, M.; Conan, P.; Oriol, L.; Cornet-Barthaux, V.; Falco, C.; Ghiglione, J.-F.; Goyet, C.; Moutin, T.; Prieur, L. Integrated survey of elemental stoichiometry (C, N, P) from the western to eastern Mediterranean Sea. Biogeosciences 2011, 8, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Dafner, E.V.; Boscolo, R.; Bryden, H.L. The N:Si:P molar ratio in the Strait of Gibraltar. Geophys. Res. Lett. 2003, 30, 1506. [Google Scholar] [CrossRef]
- Ren, L.; Wang, P.; Wang, C.; Chen, J.; Hou, J.; Qian, J. Algal growth and utilization of phosphorus studied by combined mono-culture and co-culture experiments. Environ. Pollut. 2017, 220, 274–285. [Google Scholar] [CrossRef]
- Ivančić, I.; Godrijan, J.; Pfannkuchen, M.; Marić, D.; Gašparović, B.; Djakovac, T.; Najdeka, M. Survival mechanisms of phytoplankton in conditions of stratification-induced deprivation of orthophosphate: Northern Adriatic case study. Limnol. Oceanogr. 2012, 57, 1721–1731. [Google Scholar] [CrossRef]
- Ursella, L.; Poulain, P.M.; Signell, R.P. Surface drifter derived circulation in the northern and middle Adriatic Sea: Response to wind regime and season. J. Geophys. Res. Oceans 2007, 112, C03S04. [Google Scholar] [CrossRef]
- Naudin, J.J.; Cauwet, G.; Fajon, C.; Oriol, L.; Terzic, S.; Devenon, J.-L.; Broche, P. Effect of mixing on microbial communities in the Rhone River plume. J. Mar. Syst. 2001, 28, 203–227. [Google Scholar] [CrossRef]
- Turco, M.; Quintana-Seguí, P.; Llasat, M.C.; Herrera, S.; Gutiérrez, J.M. Testing MOS precipitation downscaling for ENSEMBLES regional climate models over Spain. J. Geophys. Res. 2011, 116, D18109. [Google Scholar] [CrossRef]
- Stagl, J.C.; Hattermann, F.F. Impacts of Climate Change on the Hydrological Regime of the Danube River and Its Tributaries Using an Ensemble of Climate Scenarios. Water 2015, 7, 6139–6172. [Google Scholar] [CrossRef] [Green Version]
- Lionello, P.; Malanotte-Rizzoli, P.; Boscolo, R.; Alpert, P.; Artale, V.; Li, L.; Luterbacher, J.; May, W.; Trigo, R.; Tsimplis, M.; et al. The Mediterranean climate: An overview of the main characteristics and issues. Dev. Earth Environ. Sci. 2006, 4, 1–26. [Google Scholar] [CrossRef]
- Schubert, S.D.; Stewart, R.E.; Wang, H.; Barlow, M.; Berbery, E.H.; Cai, W.; Hoerling, M.P.; Kanikicharla, K.K.; Koster, R.D.; Lyon, B.; et al. Global meteorological drought: A synthesis of current understanding with a focus on SST drivers of precipitation deficits. J. Clim. 2016, 29, 3989–4019. [Google Scholar] [CrossRef]
- Forget, P.; André, G. Can Satellite-derived Chlorophyll Imagery Be Used to Trace Surface Dynamics in Coastal Zone? A Case Study in the Northwestern Mediterranean Sea. Sensors 2007, 7, 884–904. [Google Scholar] [CrossRef] [Green Version]
- D’Ortenzio, F.; Ribera d’Alcalà, M. On the trophic regimes of the Mediterranean Sea. Biogeosciences 2009, 6, 139–148. [Google Scholar] [CrossRef]
- Alekseenko, E.; Raybaud, V.; Espinasse, B.; Carlotti, F.; Queguiner, B.; Thouvenin, B.; Garreau, P.; Baklouti, M. Seasonal dynamics and stoichiometry of the planktonic community in the NW Mediterranean Sea: A 3D modeling approach. Ocean Dyn. 2014. [Google Scholar] [CrossRef]
- Harding, L.W.; Degobbis, D.; Precali, R. Production and fate of phytoplankton: Annual cycles and interannual variability. In Ecosystems at the Land-Sea Margin: Drainage Basin to Coastal Sea; Coastal and Estuarine Studies; Malone, T.C., Malej, A., Harding, L.W., Smodlaka, N., Turner, R.E., Eds.; American Geophysical Union: Washington, DC, USA, 1999; Volume 55, pp. 131–172. ISBN 0-87590-269-3. [Google Scholar]
- Melin, F.; Vantrepotte, V.; Clerici, M.; D’Alimonte, D.; Zibordi, G.; Berthon, J.-F.; Canuti, E. Multi-sensor satellite time series of optical properties and chlorophyll-a concentration in the Adriatic Sea. Prog. Oceanogr. 2011, 91, 229–244. [Google Scholar] [CrossRef]
- Díaz, F.; Raimbault, P.; Boudjellal, B.; Garcia, N.; Moutin, T. Early phosphorus limitation during spring in the Gulf of Lions. Mar. Ecol. Prog. Ser. 2001, 211, 51–62. [Google Scholar] [CrossRef]
- Cristofor, S.; Vadineanu, A.; Ignat, G. Importance of flood zones for nitrogen and phosphorus dynamics in the Danube Delta. Hydrobiologia 1993, 251, 143–148. [Google Scholar] [CrossRef]
- Raimbault, P.; Durrieu de Madron, X. Research activities in the Gulf of Lion (NW Mediterranean) within the 1997–2001 PNEC project. Oceanol. Acta 2003, 26, 291–298. [Google Scholar] [CrossRef]
- Nebra, A.; Alcaraz, C.; Caiola, N.; Muñoz-Camarillo, G.; Ibáñez, C. Benthic macrofaunal dynamics and environmental stress across a salt wedge Mediterranean estuary. Mar. Environ. Res. 2016, 117, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Fonda Umani, S. Pelagic production and biomass in the Adriatic Sea. Sci. Mar. 1996, 60, 65–77. [Google Scholar]
- Giani, M.; Rinaldi, A.; Degobbis, D. Mucilages in the Adriatic and Tyrrhenian Sea: An introduction. Sci. Total Environ. 2005, 353, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Aubrey, D.; Moncheva, S.; Demirov, E.; Diaconu, V.; Dimitrov, A. Environmental changes in the western Black Sea related to anthropogenic and natural conditions. J. Mar. Syst. 1996, 7, 411–425. [Google Scholar] [CrossRef]
- Daskalov, G.M.; Boicenco, L.; Grishin, A.N.; Lazar, L.; Mihneva, V.; Shlyakhov, V.A.; Zengin, M. Architecture of collapse: Regime shift and recovery in an hierarchically structured marine ecosystem. Glob. Chang. Biol. 2017, 23, 1486–1498. [Google Scholar] [CrossRef] [PubMed]
- Bostan, V.; Dominik, J.; Bostina, M.; Pardos, M. Forms of particulate phosphorus in suspension and in bottom sediment in the Danube Delta. Lakes Reserv. Res. Manag. 2000, 5, 105–110. [Google Scholar] [CrossRef]
- Ptacnik, R.; Andersen, T.; Tamminen, T. Performance of the Redfield Ratio and a family of nutrient limitation indicators as thresholds for phytoplankton N vs. P limitation. Ecosystems 2010, 13, 1201–1214. [Google Scholar] [CrossRef]
- Lazǎr, L.; Gomoiu, M.-T.; Boicenco, L.; Vasiliu, D. Total Organic Carbon (TOC) of the surface layer sediments covering the seafloor of the Romanian Black Sea coast. Geoecomarina 2012, 18, 121–132. [Google Scholar]
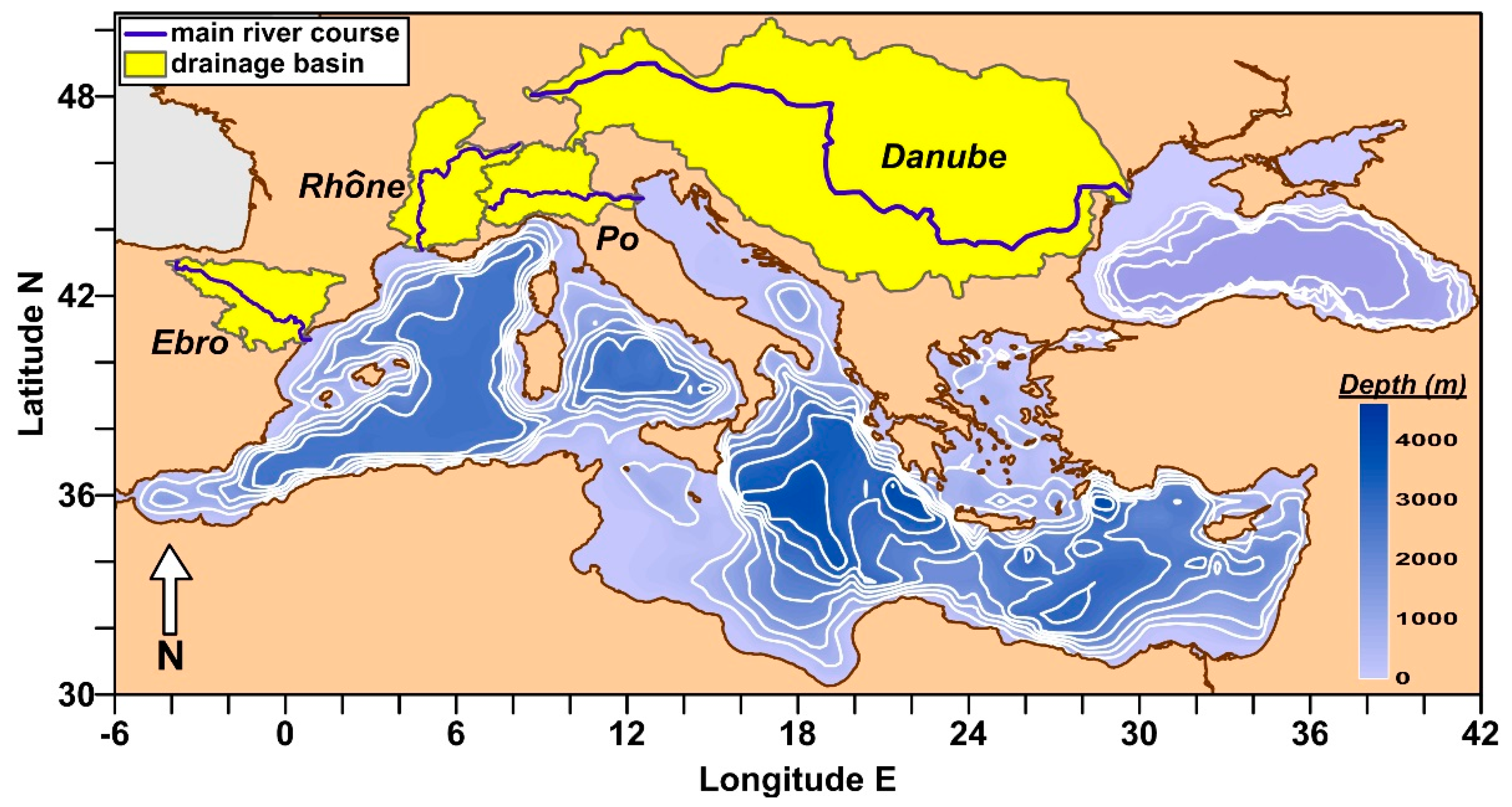
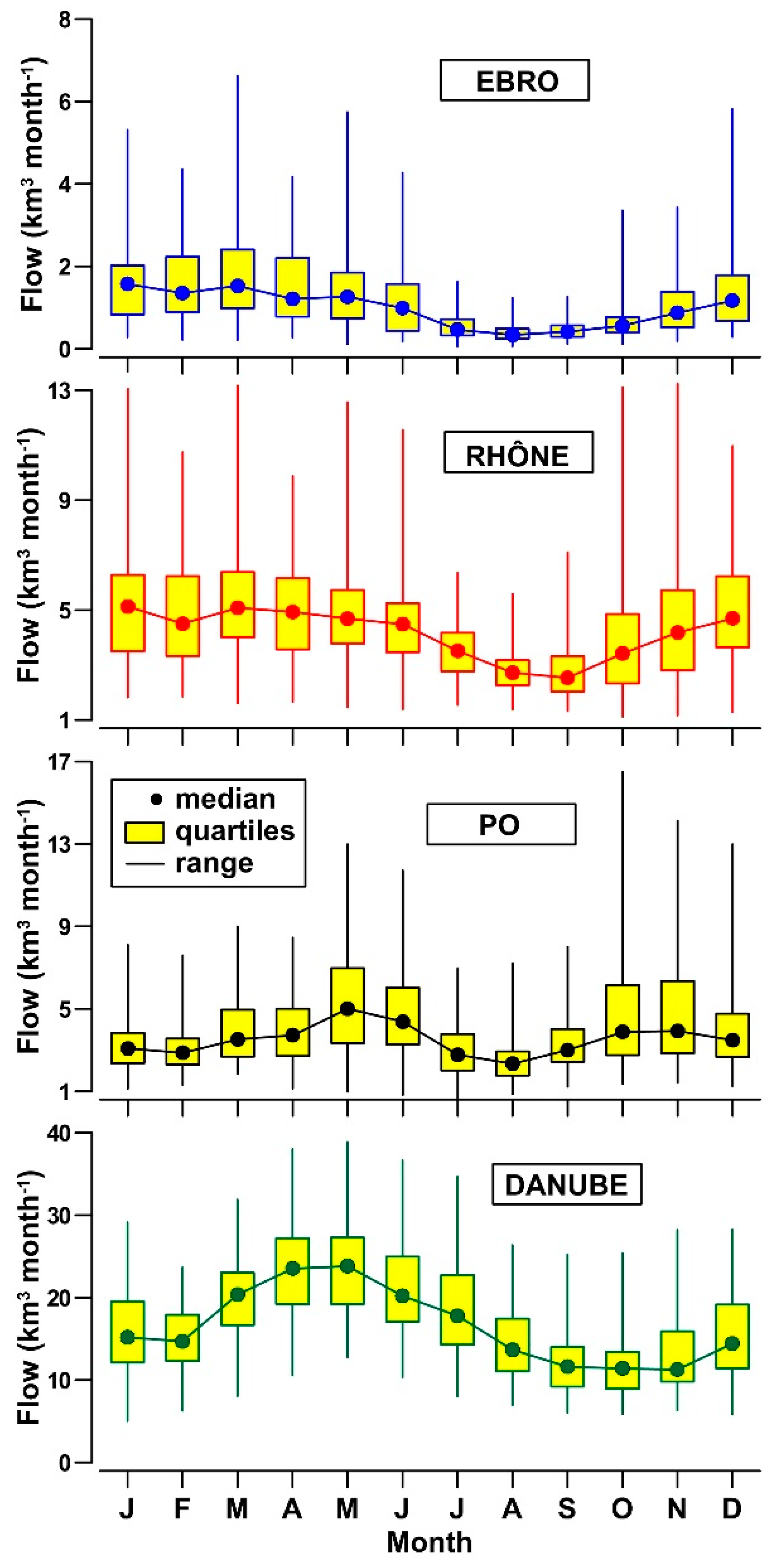
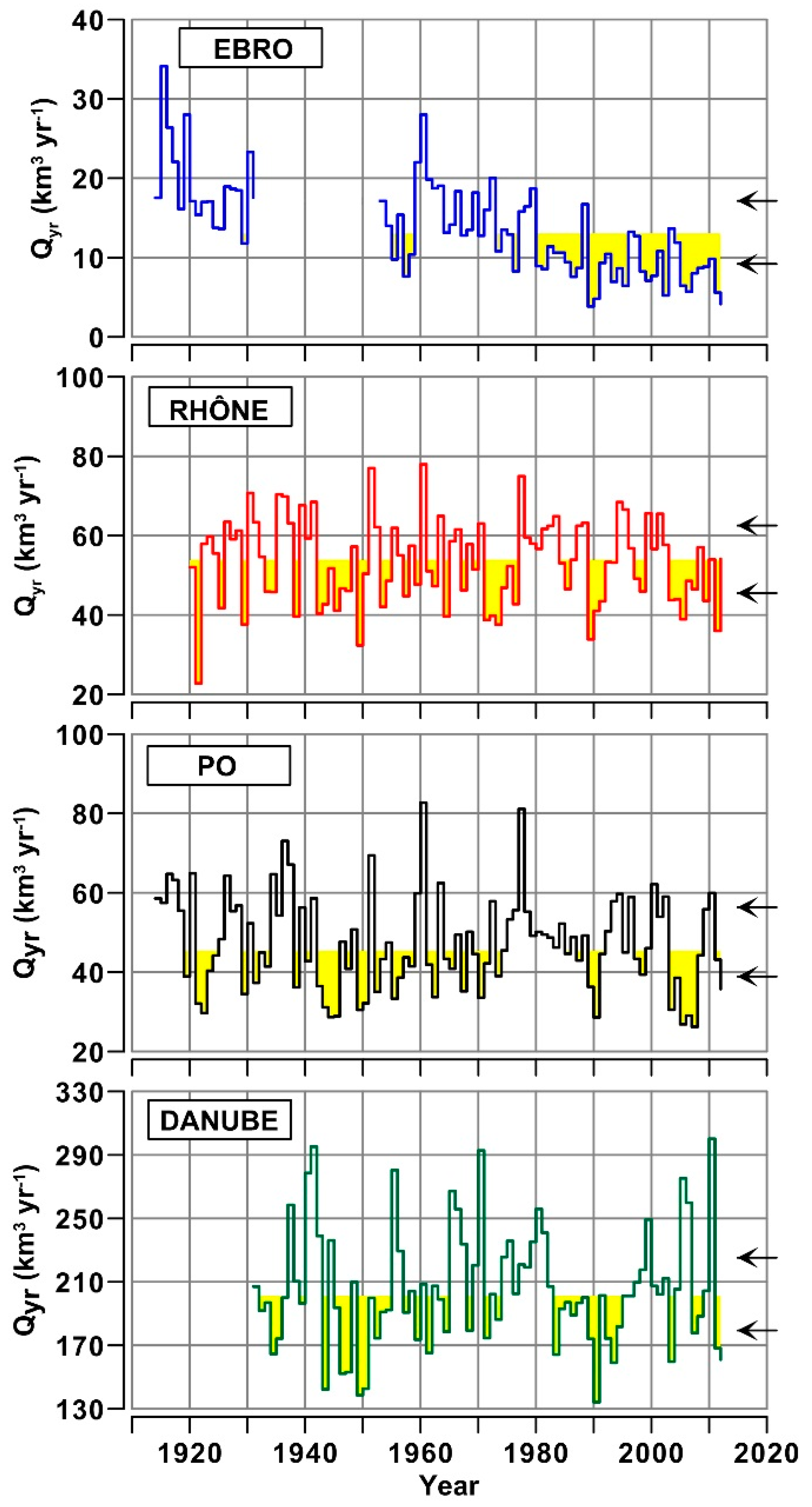
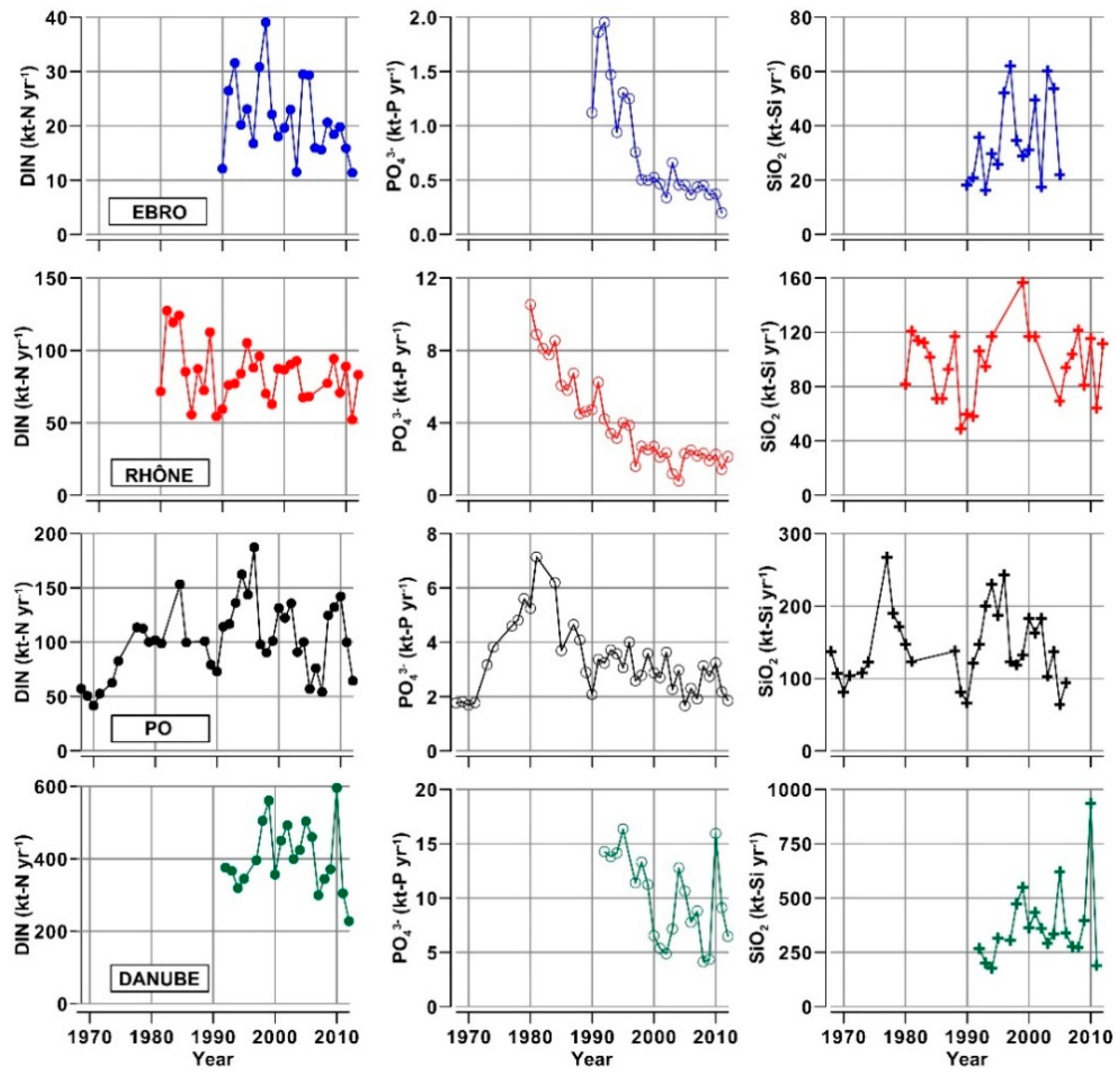
| River (Gauging Station) | Drainage Basin † (km2) | River Length ‡ (km) | Inhabitants (n · 106) | Water Flow Data Series (yr) | Annual Water Flow (km3 yr−1) Median 1th–3rd Quartile Range | ||
|---|---|---|---|---|---|---|---|
| Ebro (Tortosa) | 86,800 | 930 | 3 | 1914–1931, 1951–2012 | 13.0 | 8.7–17.1 | 3.8–34.1 |
| Rhône (Beaucaire) | 157,950 | 813 | 18 | 1920–2012 | 53.9 | 45.8–61.7 | 22.8–78.0 |
| Po (Pontelagoscuro) | 74,000 | 682 | 16 | 1914–2012 | 45.5 | 38.8–55.7 | 26.2–82.7 |
| Danube (Ceatal Izmail) | 801,500 | 2857 | 82 | 1931–2012 | 201.2 | 178.6–221.0 | 134.2–300.2 |
| Total | 1,120,250 | 5282 | 120 | 1951–2012 | 316.8 | 297.9–344.9 | 208.6–424.0 |
| River | Period | Mean Flow km3 yr−1 (m3 s−1) | Duration yr | Year of Change yr | Δ-Flow km3 yr−1 | Confidence α |
|---|---|---|---|---|---|---|
| 1914–1979 | 16.5 (539) | 66 | ||||
| Ebro | 1980–2010 | 9.1 (292) | 31 | 1980 | −7.4 | 0.0001 |
| 2011–2012 | 4.9 (154) | 2 | 2011 | −4.2 | 0.0209 | |
| Rhône | 1920–1941 | 55.9 (1772) | 22 | |||
| 1942–1976 | 50.9 (1614) | 35 | 1942 | −5.0 | 0.1292 | |
| 1977–2002 | 56.7 (1797) | 26 | 1977 | +5.8 | 0.0284 | |
| 2003–2012 | 46.6 (1478) | 10 | 2003 | −10.1 | 0.0018 | |
| Po | 1914–1941 | 51.3 (1593) | 28 | |||
| 1942–1974 | 43.7 (1390) | 33 | 1942 | −7.6 | 0.0163 | |
| 1975–2002 | 50.4 (1626) | 28 | 1975 | +6.7 | 0.0183 | |
| 2003–2008 | 32.6 (1058) | 6 | 2003 | −17.8 | 0.0006 | |
| 2009–2012 | 48.7 (1551) | 4 | 2009 | +16.1 | 0.0548 | |
| Danube | 1931–1945 | 212.4 (6729) | 15 | |||
| 1946–1964 | 189.0 (5990) | 19 | 1946 | −23.4 | 0.0910 | |
| 1965–1982 | 225.3 (7141) | 18 | 1965 | +36.3 | 0.0016 | |
| 1983–2010 | 201.5 (6381) | 28 | 1983 | −23.8 | 0.0208 | |
| 2011–2012 | 164.6 (5213) | 2 | 2011 | −36.9 | 0.0003 |
| River (Station) | Ebro (Tortosa) | Rhône (Arles) | Po (Polesella, Serravalle, Pontelagoscuro) | Danube (Reni, Sulina) |
|---|---|---|---|---|
| TSM (mg L−1) | 7.5 (5.0–11.0) | 12.3 (7.4–29) | 46.8 (28.4–110) | 26.5 (15.0–44) |
| NO3− (µmol L−1) | 165 (142–194) | 97 (76–119) | 161 (124–202) | 111 (82–141) |
| NH4+ (µmol L−1) | 2.8 (2.2–5.5) | 3.3 (1.5–5.6) | 5.3 (2.6–10.2) | 15.0 (9.1–25) |
| NO2− (µmol L−1) | 1.5 (1.1–2.0) | 1.6 (1.1–2.0) | 2.0 (1.3–3.0) | 2.1 (1.4–3.4) |
| DIN (µmol L−1) | 171 (146–201) | 102 (84–125) | 168 (302–216) | 132 (105–163) |
| PO43− (µmol L−1) | 2.3 (1.5–4.5) | 1.5 (1.1–1.9) | 1.9 (1.6–2.6) | 1.3 (0.6–2.0) |
| SiO2 (µmol L−1) | 130 (93–160) † | 64 (51–78) | 113 (85–134) | 61 (37–82) |
| TN (µmol L−1) | 211 (180–234) † | 123 (101–148) * | 249 (197–304) + | 147 (121–187) °° |
| TP (µmol L−1) | 2.9 (2.6–3.6) | 2.6 (2.2–3.2) * | 4.8 (4.2–6.1) | 2.8 (1.9–4.0) |
| TOC (µmol L−1) | 270 (208–341) † | 241 (199–304) | 340 (268–460) ° | 398 (294–569) ** |
| ON (µmol L−1) | 28.1 (22–46) † | 18.2 (12.7–29) * | 76 (41–145) | 25.3 (19.2–33) °° |
| OP (µmol L−1) | 1.0 (0.4–1.7) | 1.2 (0.8–1.8) * | 2.9 (1.9–4.1) | 1.2 (0.6–2.1) |
| DIN/PO43− (molar) | 72 (36–118) | 73 (52–100) | 85 (61–107) | 94 (62–228) |
| Si/DIN (molar) | 0.7 (0.5–1.0) † | 0.6 (0.5–0.7) | 0.6 (0.5–0.8) | 0.5 (0.4–0.8) °° |
| TN/TP (molar) | 58 (44–70) † | 45 (37–59) * | 49 (37–64) | 55 (38–82) °° |
| ON/OP (molar) | 20 (11–31) | 15 (10–22) * | 25 (16–46) | 17 (9–41) °° |
| ON/TN (%) | 15 (11–21) † | 16 (10–22) * | 34 (20–49) | 16 (13–22) °° |
| OP/TP (%) | 32 (13–57) | 49 (35–63) * | 60 (47–72) | 49 (30–72) |
| Descriptor | Ebro NE Shelf of Spain | Rhône Gulf of Lion | Po NW Adriatic Sea | Danube NW Black Sea | Environmental Impacts |
|---|---|---|---|---|---|
| Annual freshwater discharge | Long-term decrease. | Interannual to multi-decadal oscillations increased recently. | Interannual to multi-decadal oscillations increased recently. | Interannual to multi-decadal oscillations. | Marked regional differences of river discharges. Increased droughts in summer more marked in SW Europe than in SE Europe. Increased oscillations of summer runoff in SE Europe. |
| Long-term trend of monthly flows | Decrease in all months. | Decrease in late spring and summer. | Decrease in summer. | Small decrease in summer. | |
| Long-term trend of flow oscillations | Decrease in all months. | Increase in summer. | Increase in summer. | Increase in spring and summer. | |
| Dry seasons | Summer. | Summer. | Winter and summer. | Winter and summer. | |
| Flow regime | Low with a high incidence of freshets. | High with low incidence of freshets. | Intermediate with a high incidence of freshets. | Very high with low incidence of freshets. | |
| Concentrations of TSM and nutrients | Low TSM, high nutrients. | Low TSM, low nutrients. | High TSM, high nutrients. | Medium TSM, medium nutrients. | Impact of the decrease of TSM transport on estuarine and coastal areas. Distinct impacts of seasonal changes of nutrients on riverine, estuarine and coastal ecosystems. |
| Variability of nutrient concentrations | Seasonal cycle (except PO43−). | Seasonal cycle (for PO43− only since 2007). | Seasonal cycle. | Seasonal cycle | |
| Variability of DIN/PO43−, TN/TP and Si/DIN ratios | Seasonal oscillations (low N/P in summer, low Si/DIN in spring). | Seasonal oscillations (low N/P in summer, low Si/DIN in spring). | Seasonal oscillations (low N/P in summer, low Si/DIN in spring). | Rather constant DIN/PO43− through the year, high TN/TP in spring. | |
| Incidence of organic nitrogen and phosphorus on TN and TP pools | ON: low OP: low | ON: low OP: medium | ON: high OP: high | ON: low OP: high | Potential growth of marine plankton species able to utilize riverine ON and OP. |
| Recent trends of annual loads of TSM, nutrients and OM | Decreases of TSM, N, P and TOC transport since the 1990s. Oscillations of SiO2. | Decrease of PO43− and ON transport since the 1980s. Oscillations of SiO2 and DIN. | Decreases of NH4+, NO2− and PO43− and TP transport since the 1980s. Oscillations of SiO2. | Decreased transport of NH4+ since the 1990s. Oscillating N, P transports since the 2000s. | Reduction of PO43− transport leading to phytoplankton biomass reduction. Oscillation of nutrient loads linked to runoff variability. |
| Marine region of freshwater influence | Small, limited by continental shelf orography. | Medium, limited by continental shelf orography. | Large, enhanced by continental shelf orography. | Very large, enhanced by continental shelf orography. | Larger impacts in the coastal zones of Po and Danube, even if river loads are reduced. |
| Eutrophication potential of river nutrient loads in the receiving coastal zones | Until 1995, high and balanced nutrient loads. Afterwards, excesses of DIN and SiO2 over PO43−. Low weight of OP in TP. | Until 1990, high and balanced nutrient loads. Afterwards, excesses of DIN and SiO2, over PO43−. Medium weight of OP in TP. | Until 1990, high and balanced nutrient loads, with a surplus of OP. Afterwards, excesses of DIN and SiO2, over PO43−. Persistent high weight of OP in TP. | Since 2000s, excesses of DIN and SiO2, over PO43−. Medium weight of OP in TP. | Shift form eutrophic conditions to oligotrophic, but still degraded, conditions around the 1990s. PO43− scarcity sometime potentially compensated by OP bioavailability. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cozzi, S.; Ibáñez, C.; Lazar, L.; Raimbault, P.; Giani, M. Flow Regime and Nutrient-Loading Trends from the Largest South European Watersheds: Implications for the Productivity of Mediterranean and Black Sea’s Coastal Areas. Water 2019, 11, 1. https://doi.org/10.3390/w11010001
Cozzi S, Ibáñez C, Lazar L, Raimbault P, Giani M. Flow Regime and Nutrient-Loading Trends from the Largest South European Watersheds: Implications for the Productivity of Mediterranean and Black Sea’s Coastal Areas. Water. 2019; 11(1):1. https://doi.org/10.3390/w11010001
Chicago/Turabian StyleCozzi, Stefano, Carles Ibáñez, Luminita Lazar, Patrick Raimbault, and Michele Giani. 2019. "Flow Regime and Nutrient-Loading Trends from the Largest South European Watersheds: Implications for the Productivity of Mediterranean and Black Sea’s Coastal Areas" Water 11, no. 1: 1. https://doi.org/10.3390/w11010001
APA StyleCozzi, S., Ibáñez, C., Lazar, L., Raimbault, P., & Giani, M. (2019). Flow Regime and Nutrient-Loading Trends from the Largest South European Watersheds: Implications for the Productivity of Mediterranean and Black Sea’s Coastal Areas. Water, 11(1), 1. https://doi.org/10.3390/w11010001