Recognition of Patterns of Benthic Diatom Assemblages within a River System to Aid Bioassessment




Abstract
1. Introduction
2. Materials and Methods
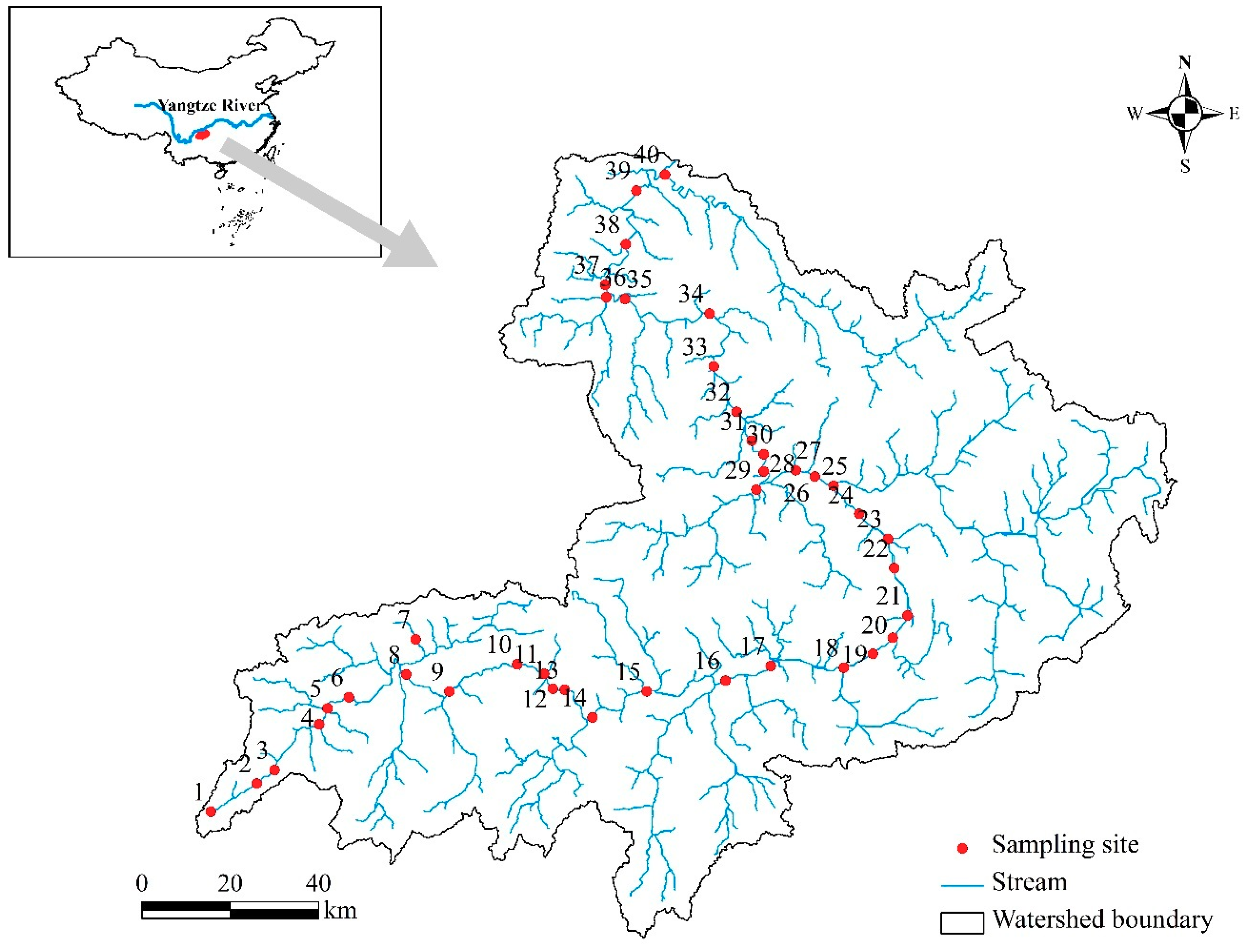
2.1. Study Area
2.2. Benthic Diatom Sampling and Identification and Diatom Index Calculation
2.3. Environmental Variables
2.4. Statistical Analysis
3. Results
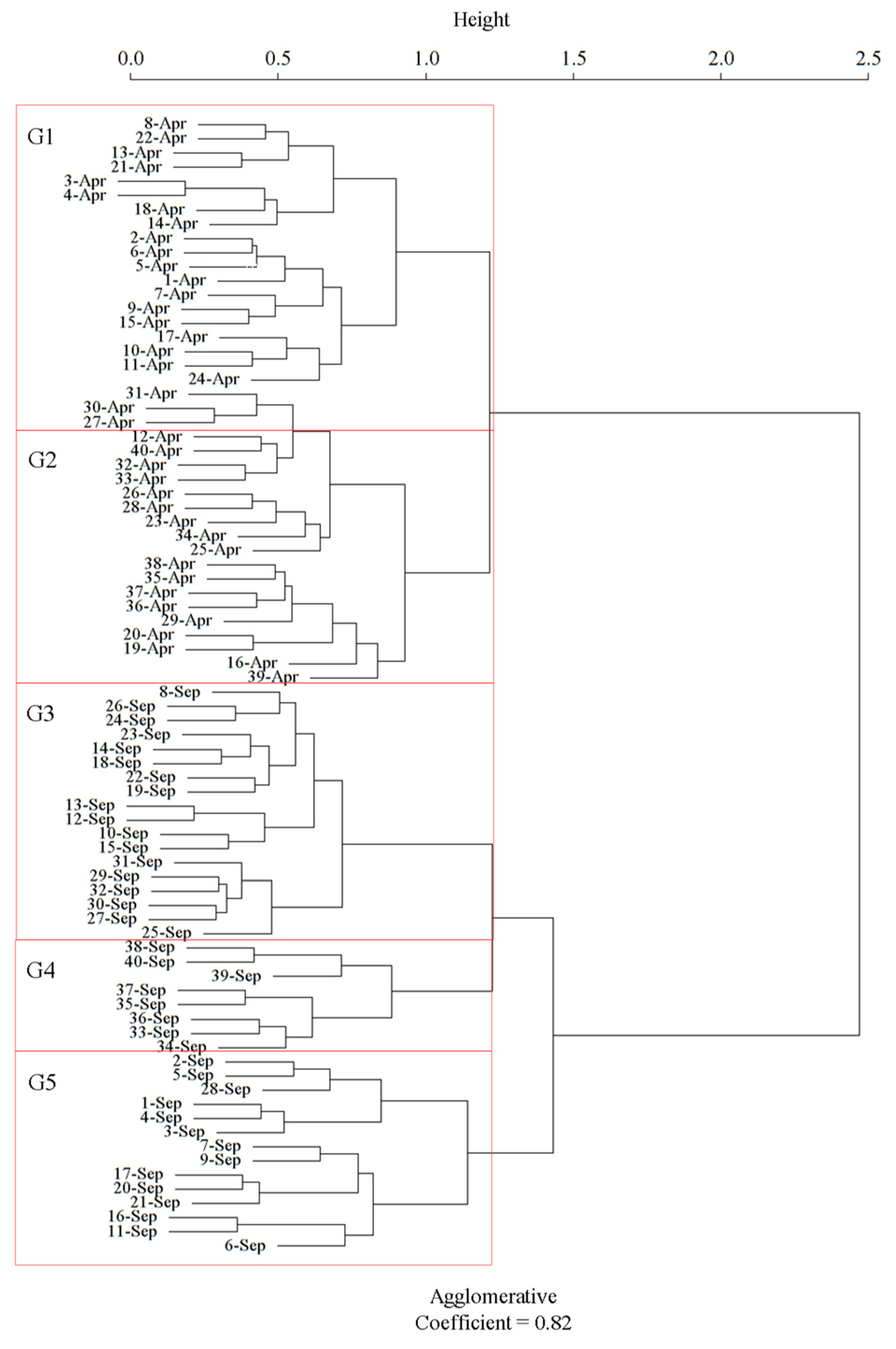
3.1. Site Classification
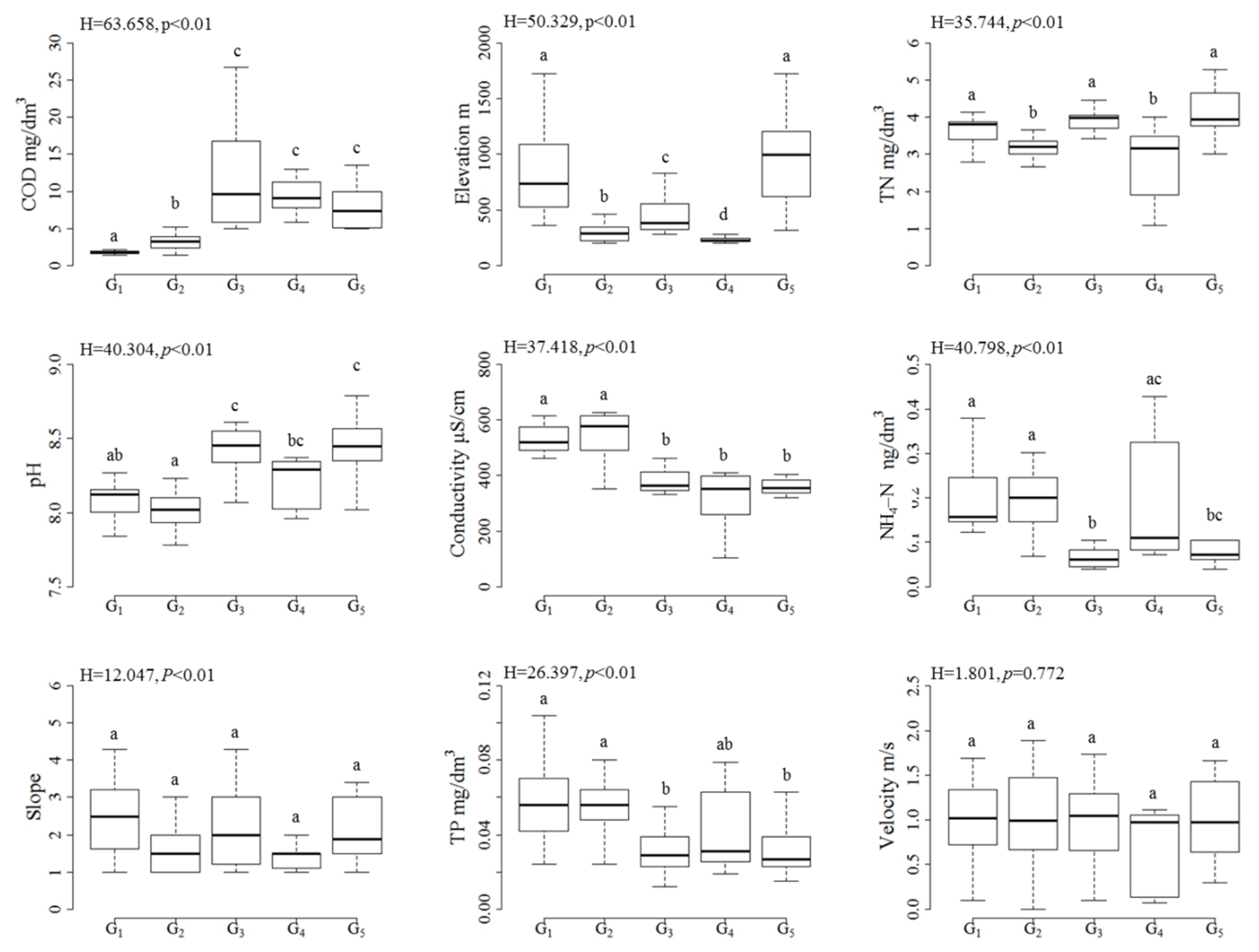
3.2. Differences in Environmental Variables among the Site Groups
3.3. Indicator Taxa for Site Groups
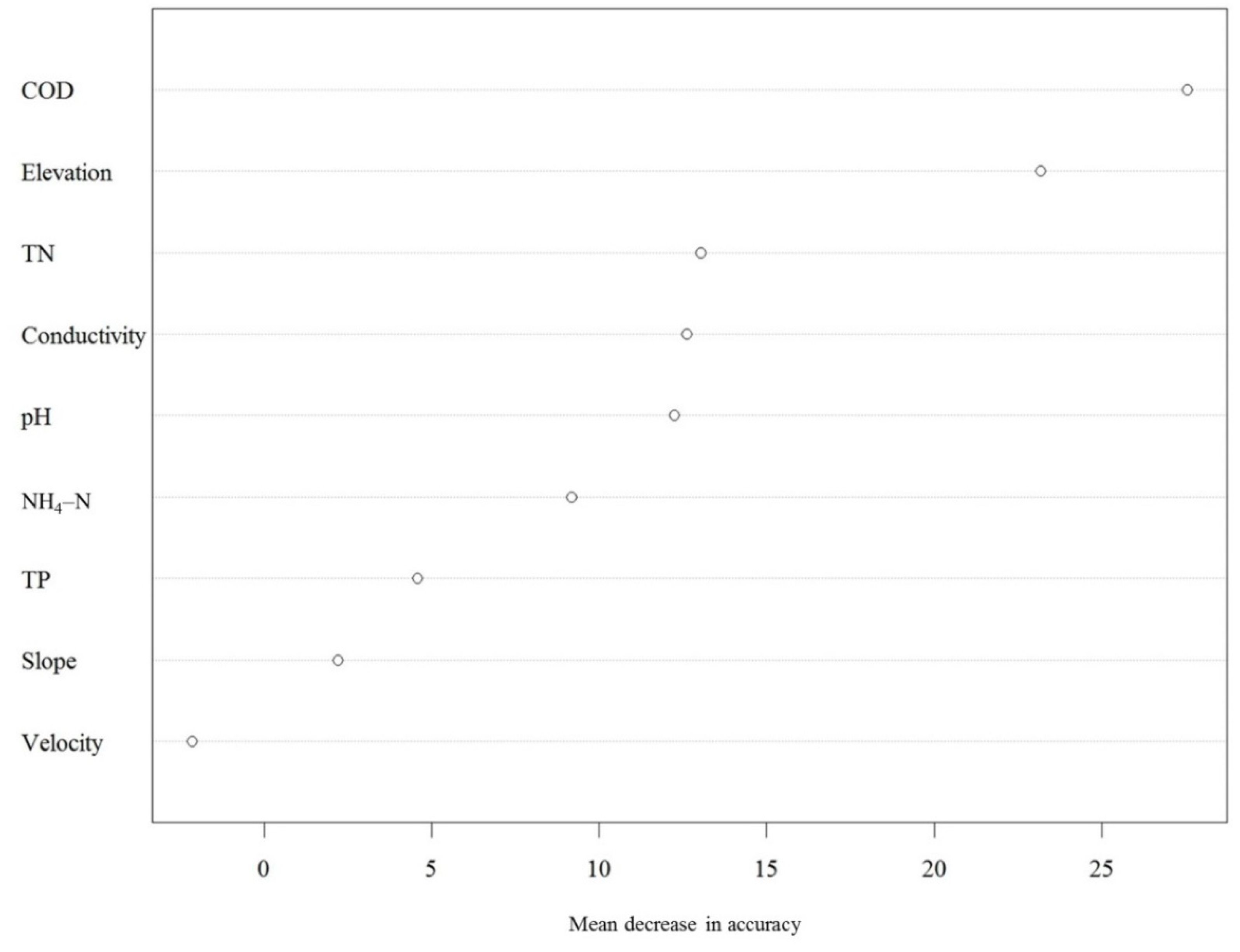
3.4. Environmental Variables Best Explaining the Variation in European Diatom Indices
4. Discussion
4.1. Associations between Algal Patterns and Water Quality Gradients
4.2. Potential Applications
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yuan, L.; Norton, S.B. Assessing the relative severity of stressors at a watershed scale. Environ. Monit. Assess. 2004, 98, 323–349. [Google Scholar] [CrossRef]
- Flotemersch, J.E.; Leibowitz, S.G.; Hill, R.A.; Stoddard, J.L.; Thoms, M.C.; Tharme, R.E. A watershed integrity definition and assessment approach to support strategic management of watersheds. River Res. Appl. 2016, 32, 1654–1671. [Google Scholar] [CrossRef]
- Jähnig, S.; Tonkin, J.; Gies, M.; Domisch, S.; Hering, D.; Haase, P. Severity multipliers as a methodology to explore potential effects of climate change on stream bioassessment programs. Water 2017, 9, 188. [Google Scholar] [CrossRef]
- Zhang, M.; Muñoz-Mas, R.; Martínez-Capel, F.; Qu, X.; Zhang, H.; Peng, W.; Liu, X. Determining the macroinvertebrate community indicators and relevant environmental predictors of the Hun-Tai River Basin (Northeast China): A study based on community patterning. Sci. Total Environ. 2018, 634, 749–759. [Google Scholar] [CrossRef]
- Hering, D.; Borja, A.; Carstensen, J.; Carvalho, L.; Elliott, M.; Feld, C.K.; Heiskanen, A.-S.; Johnson, R.K.; Moe, J.; Pont, D.; et al. The European Water Framework Directive at the age of 10: A critical review of the achievements with recommendations for the future. Sci. Total Environ. 2010, 408, 4007–4019. [Google Scholar] [CrossRef]
- Scrimgeour, G.J.; Hvenegaard, P.J.; Tchir, J. Cumulative industrial activity alters lotic fish assemblages in two boreal forest watersheds of Alberta, Canada. Environ. Manag. 2008, 42, 957–970. [Google Scholar] [CrossRef]
- Mykrä, H.; Saarinen, T.; Tolkkinen, M.; McFarland, B.; Hämäläinen, H.; Martinmäki, K.; Kløve, B. Spatial and temporal variability of diatom and macroinvertebrate communities: How representative are ecological classifications within a river system? Ecol. Indic. 2012, 18, 208–217. [Google Scholar] [CrossRef]
- Herlihy, A.T.; Paulsen, S.G.; Sickle, J.V.; Stoddard, J.L.; Hawkins, C.P.; Yuan, L.L. Striving for consistency in a national assessment: The challenges of applying a reference-condition approach at a continental scale. J. N. Am. Benthol. Soc. 2008, 27, 860–877. [Google Scholar] [CrossRef]
- Hering, D.; Johnson, R.K.; Kramm, S.; Schmutz, S.; Szoszkiewicz, K.; Verdonschot, P.F.M. Assessment of European streams with diatoms, macrophytes, macroinvertebrates and fish: A comparative metric-based analysis of organism response to stress. Freshwat. Biol. 2006, 51, 1757–1785. [Google Scholar] [CrossRef]
- Cao, Y.; Hawkins, C.P.; Olson, J.; Kosterman, M.A. Modeling natural environmental gradients improves the accuracy and precision of diatom-based indicators. J. N. Am. Benthol. Soc. 2007, 26, 566–585. [Google Scholar] [CrossRef]
- Tang, T.; Stevenson, R.J.; Infante, D.M. Accounting for regional variation in both natural environment and human disturbance to improve performance of multimetric indices of lotic benthic diatoms. Sci. Total Environ. 2016, 568, 1124–1134. [Google Scholar] [CrossRef]
- Olson, J.R.; Hawkins, C.P. Developing site-specific nutrient criteria from empirical models. Freshw. Sci. 2013, 32, 719–740. [Google Scholar] [CrossRef]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Bellinger, B.J.; Angradi, T.R.; Bolgrien, D.W.; Jicha, T.M.; Hill, B.H.; Reavie, E.D. Longitudinal variation and response to anthropogenic stress in diatom assemblages of the Lower Mississippi River, USA. River Syst. 2013, 21, 29–54. [Google Scholar] [CrossRef]
- Ferreira, F.; Petrere, M. The fish zonation of the Itanhaém river basin in the Atlantic Forest of southeast Brazil. Hydrobiologia 2009, 636, 11–34. [Google Scholar] [CrossRef]
- Angradi, T.R.; Pearson, M.S.; Jicha, T.M.; Taylor, D.L.; Bolgrien, D.W.; Moffett, M.F.; Blocksom, K.A.; Hill, B.H. Using stressor gradients to determine reference expectations for great river fish assemblages. Ecol. Indic. 2009, 9, 748–764. [Google Scholar] [CrossRef]
- Jiang, X.; Xiong, J.; Qiu, J.; Wu, J.; Wang, J.; Xie, Z. Structure of macroinvertebrate communities in relation to environmental variables in a Subtropical Asian river system. Int. Rev. Hydrobiol. 2010, 95, 42–57. [Google Scholar] [CrossRef]
- Pan, Y.; Hughes, R.M.; Herlihy, A.T.; Kaufmann, P.R. Non-wadeable river bioassessment: Spatial variation of benthic diatom assemblages in Pacific Northwest rivers, USA. Hydrobiologia 2012, 684, 241–260. [Google Scholar] [CrossRef]
- Wu, N.; Faber, C.; Sun, X.; Qu, Y.; Wang, C.; Ivetic, S.; Riis, T.; Ulrich, U.; Fohrer, N. Importance of sampling frequency when collecting diatoms. Sci. Rep. 2016, 6, 36950. [Google Scholar] [CrossRef]
- Lavoie, I.; Hamilton, P.B.; Wang, Y.K.; Dillon, P. J.; Campeau, S. A. comparison of stream bioassessment in Québec (Canada) using six European and North American diatom-based indices. In Diatom Taxonomy, Ultrastructure and Ecology: Modern Methods and Timeless Questions; Kociolek, J.P., Theriot, E.C., Stevenson, R.J., Eds.; Schweizerbart Science Publishers: Stuttgart, Germany, 2009; pp. 37–56. ISBN 9783443510572. [Google Scholar]
- Bere, T. Challenges of diatom-based biological monitoring and assessment of streams in developing countries. Environ. Sci. Pollut. Res. 2016, 23, 5477–5486. [Google Scholar] [CrossRef]
- Kilgour, B.W.; Stanfield, L.W. Hindcasting reference conditions in streams. In Landscape Influences on Stream Habitats and Biological Assemblages; Hughes, R., Wang, L., Seelbach, P., Eds.; American Fisheries Society Symposium: Bethesda, MD, USA, 2006; pp. 623–639. [Google Scholar]
- Birk, S.; van Kouwen, L.; Willby, N. Harmonising the bioassessment of large rivers in the absence of near-natural reference conditions—A case study of the Danube River. Freshw. Biol. 2012, 57, 1716–1732. [Google Scholar] [CrossRef]
- Hawkins, C.P.; Olson, J.R.; Hill, R.A. The reference condition: Predicting benchmarks for ecological and water-quality assessments. J. N. Am. Benthol. Soc. 2010, 29, 312–343. [Google Scholar] [CrossRef]
- Stevenson, R.; Pan, Y.; Van Dam, H. Assessing environmental conditions in rivers and streams with diatoms. In The Diatoms: Applications for the Environmental and Earth Sciences, 2nd ed.; Smol, J.P., Stoermer, E.F., Eds.; Cambridge University Press: Cambridge, UK, 2010; ISBN 9780521509961. [Google Scholar]
- Huang, Z. On the development and conservation for the resources and ecology of Chishui river basin. Res. Environ. Yangtze Basin 2003, 12, 333–339. (In Chinese) [Google Scholar] [CrossRef]
- Wu, J.; Zhao, H.; Miao, Z.; Chen, Y.; Zhang, F.; Wang, J. Status and conservation of fish resources in the Chishui River. Biodivers. Sci. 2010, 18, 162–168. (In Chinese) [Google Scholar]
- Wang, Z.; Jiang, L.; Huang, M.; Zhang, C.; Yu, X. Biodiversity status and its conservation strategy in the Chishui river basin. Res. Environ. Yangtze Basin 2007, 16, 175–180. (In Chinese) [Google Scholar] [CrossRef]
- Chi, S.; Gong, Y.; Wang, H.; Zheng, J.; Hu, J.; Hu, J.; Dong, F. A pilot macroinvertebrate-based multimetric index (MMI-CS) for assessing the ecological status of the Chishui River basin, China. Ecol. Indic. 2017, 83, 84–95. [Google Scholar] [CrossRef]
- Tang, T.; Tang, T.; Tan, L.; Gu, Y.; Jiang, W.; Cai, Q. Identifying community thresholds for lotic benthic diatoms in response to human disturbance. Sci. Rep. 2017, 7, 4134. [Google Scholar] [CrossRef]
- Tang, T.; Jia, X.; Jiang, W.; Cai, Q. Multi-scale temporal dynamics of epilithic algal assemblages: Evidence from a Chinese subtropical mountain river network. Hydrobiologia 2016, 770, 289–299. [Google Scholar] [CrossRef]
- Hu, H.; Wei, Y. The Freshwater Algae of China: Systematics, Taxonomy and Ecology; Science Press: Beijing, China, 2006; ISBN 7030166337. [Google Scholar]
- Qi, Y. Flora Algarum Sinicarum Aquae Dulcis. Vol. 4. Bacillariophyta Centreae; Science Press: Beijing, China, 1995; ISBN 7030044223. [Google Scholar]
- Shi, Z. Flora Algarum Sinicarum Aquae Dulcis, Tomus 12, Bacillariophyta-Ghophonemaceae; Science Press: Beijing, China, 2004; ISBN 9787030133366. (In Chinese) [Google Scholar]
- Krammer, K. Diatoms of Europe: Diatoms of the European Inland Waters and Comparable Habitats. vol. 1: The Genus Pinnularia; A.R.G.Gantner Verlag, K.G.: Ruggell, Liechtenstein, 2000; ISBN 978-3-904144-24-7. [Google Scholar]
- Krammer, K. Diatoms of Europe: Diatoms of the European Inland Waters and Comparable Habitats. Vol. 3: Cymbella; A.R.G.Gantner Verlag, K.G.: Ruggell, Liechtenstein, 2002; ISBN 978-3-904144-84-1. [Google Scholar]
- Krammer, K. Diatoms of Europe: Diatoms of the European Inland Waters and Comparable Habitats. Vol. 4: Cymbopleura, Delicata, Navicymbula, Gomphocymbellopsis, Afrocymbella; A.R.G. Gantner Verlag, K.G: Ruggell, Liechtenstein, 2003; ISBN 978-3-904144-99-5. [Google Scholar]
- Lange-Bertalot, H.; Bak, M.; Witkowski, A.; Tagliaventi, N. Diatoms of Europe: Diatoms of the European Inland Waters and Comparable Habitats. Vol. 2: Navicula Sensu Stricto,10 Genera Separated from Navicula Sensu Lato, Frustulia; A.R.G. Gantner Verlag, K.G.: Ruggell, Liechtenstein, 2001; ISBN 978-3-904144-78-0. [Google Scholar]
- Lecointe, C.; Coste, M.; Prygiel, J. “Omnidia”: Software for taxonomy, calculation of diatom indices and inventories management. Hydrobiologia 1993, 269, 509–513. [Google Scholar] [CrossRef]
- Prygiel, J.; Coste, M.; Bukowska, J. Review of the major diatom-based techniques for the quality assessment of rivers-State of the art in Europe. In Use of Algae for Monitoring Rivers III; Prygiel, J., Whitton, B.A., Buckowska, J., Eds.; Agence de I’Eau Artois-Picar-die Press: Douai Cedex, France, 1999; ISBN 9782950208354. [Google Scholar]
- Wei, F. Monitoring and Analysis Methods of Water and Wastewater; Chinese Environmental Sciences Publishment: Beijing, China, 2002. [Google Scholar]
- Tornés, E.; Leira, M.; Sabater, S. Is the biological classification of benthic diatom communities concordant with ecotypes? Hydrobiologia 2012, 695, 43–55. [Google Scholar] [CrossRef]
- Hawkins, C.P.; Vinson, M.R. Weak correspondence between landscape classifications and stream invertebrate assemblages: Implications for bioassessment. J. N. Am. Benthol. Soc. 2000, 19, 501–517. [Google Scholar] [CrossRef]
- Cutler, D.R.; Edwards, T.C., Jr.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef]
- Snelder, T.; Ortiz, J.; Booker, D.; Lamouroux, N.; Pella, H.; Shankar, U. Can bottom-up procedures improve the performance of stream classifications? Aquat. Sci. 2012, 74, 45–59. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Stevenson, R.J.; Bahls, L.L. Periphyton Protocols; US Environmental Protection Agency: Washington, DC, USA, 1999; pp. 6–22.
- Zhang, C.; Wang, Y.; Qiao, H.; Sun, L.; Dong, L. The impacts of liquor enterprises on water environment of Chishui River Basin. Water Resour. Protect. 2015, 31, 62–66. (In Chinese) [Google Scholar] [CrossRef]
- Teng, Z.; Ding, A.; Li, Y.; Jing, R.; Liu, Y. Sources of water pollution and their spatio temporal variations in the upper reach of the Chishui River. J. Beijng Norm. Univ. (Nat. Sci.) 2016, 52, 3222–3327. (In Chinese) [Google Scholar] [CrossRef]
- Soininen, J. Responses of epilithic diatom communities to environmental gradients in some Finnish rivers. Int. Rev. Hydrobiol. 2002, 87, 11–24. [Google Scholar] [CrossRef]
- Tan, X.; Xia, X.; Zhao, Q.; Zhang, Q. Temporal variations of benthic diatom community and its main influencing factors in a subtropical river, China. Environ. Sci. Pollut. R. 2014, 21, 434–444. [Google Scholar] [CrossRef]
- Soininen, J. Environmental and spatial control of freshwater diatoms—A review. Diatom Res. 2007, 22, 473–490. [Google Scholar] [CrossRef]
- Damanik-Ambarita, M.; Everaert, G.; Forio, M.; Nguyen, T.; Lock, K.; Musonge, P.; Suhareva, N.; Dominguez-Granda, L.; Bennetsen, E.; Boets, P.; et al. Generalized linear models to identify key hydromorphological and chemical variables determining the occurrence of macroinvertebrates in the Guayas River Basin (Ecuador). Water 2016, 8, 297. [Google Scholar] [CrossRef]
- Tonkin, J.D.; Shah, T.R.D.; Shah, D.N.; Hoppeler, F.; Jähnig, S.C.; Pauls, S.U. Metacommunity structuring in Himalayan streams over large elevational gradients: The role of dispersal routes and niche characteristics. J. Biogeogr. 2017, 44, 62–74. [Google Scholar] [CrossRef]
- Van Dam, H.; Mertens, A.; Sinkeldam, J. A coded checklist and ecological indicator values of freshwater diatoms from the Netherlands. Neth. J. Aquat. Ecol. 1994, 28, 117–133. [Google Scholar] [CrossRef]
- Smucker, N.J.; Detenbeck, N.E.; Morrison, A.C. Diatom responses to watershed development and potential moderating effects of near-stream forest and wetland cover. Freshw. Sci. 2013, 32, 230–249. [Google Scholar] [CrossRef]
- Tan, X.; Zhang, Q.; Burford, M.A.; Sheldon, F.; Bunn, S.E. Benthic diatom based indices for water quality assessment in two subtropical streams. Front. Microbiol. 2017, 8, 601. [Google Scholar] [CrossRef]
- Huang, Q.; Gao, J.; Cai, Y.; Yin, H.; Gao, Y.; Zhao, J.; Liu, L.; Huang, J. Development and application of benthic macroinvertebrate-based multimetric indices for the assessment of streams and rivers in the Taihu Basin, China. Ecol. Indic. 2015, 48, 649–659. [Google Scholar] [CrossRef]
- Allan, J.D. Landscapes and riverscapes: The influence of land use on stream ecosystems. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 257–284. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | A | B | lndVal | p-Value |
|---|---|---|---|---|
| G1 (N = 2) | ||||
| Rossithidium linearis (Smith) Round et Bukhtiyarova | 0.743 | 1.000 | 0.862 | 0.005 |
| Nitzschia acicularis (Kützing) Smith | 0.645 | 0.421 | 0.521 | 0.020 |
| G2 (N = 22) | ||||
| Encyonema muelleri (Hustedt) Mann | 0.970 | 0.571 | 0.744 | 0.005 |
| Melosira granulata (Ehrenberg) Ralfs | 0.966 | 0.571 | 0.743 | 0.005 |
| Ulnaria ulna (Nitzsch) Compère | 0.635 | 0.857 | 0.738 | 0.005 |
| Nitzschia pseudofonticola Hustedt | 0.757 | 0.714 | 0.735 | 0.005 |
| Gomphonema clavatum Agardh | 0.806 | 0.667 | 0.733 | 0.005 |
| Diatoma vulgare Bory | 0.730 | 0.714 | 0.722 | 0.010 |
| Halamphora coffeaeformis (Agardh) Levkov | 0.890 | 0.524 | 0.683 | 0.005 |
| Navicula cryptocephala Kützing | 0.478 | 0.952 | 0.674 | 0.005 |
| Cymbella tumidula Grunow | 0.945 | 0.476 | 0.671 | 0.005 |
| Melosira varians Agardh | 0.435 | 1.000 | 0.659 | 0.010 |
| Achnanthes rossii Hustedt | 0.977 | 0.429 | 0.647 | 0.005 |
| Diatoma moniliformis (Kützing) Williams | 0.786 | 0.524 | 0.642 | 0.005 |
| Amphora pediculus (Kützing) Grunow | 0.706 | 0.571 | 0.635 | 0.005 |
| Encyonopsis behrei (Foged) Krammer & Metzeltin | 0.719 | 0.524 | 0.614 | 0.010 |
| Cocconeis pediculus Ehrenberg | 0.395 | 0.952 | 0.613 | 0.010 |
| Delicata gadjiana (Maillard) Krammer | 0.943 | 0.381 | 0.599 | 0.005 |
| Cyclotella meneghiniana Kützing | 0.894 | 0.381 | 0.583 | 0.020 |
| Encyonema silesiacum (Bleisch) Mann | 0.884 | 0.381 | 0.580 | 0.010 |
| Fragilaria vaucheriae (Grunow) Jørgensen | 1.000 | 0.333 | 0.577 | 0.010 |
| Cyclotella sp. | 1.000 | 0.286 | 0.535 | 0.010 |
| Gomphonema olivaceum (Hornemann) Ehrenberg | 0.594 | 0.476 | 0.532 | 0.010 |
| Achnanthes impexiformis Lange-Bertalot | 0.892 | 0.286 | 0.505 | 0.015 |
| G3 (N = 16) | ||||
| Luticola mutica (Kützing) Mann | 0.929 | 0.944 | 0.937 | 0.005 |
| Cymbella tumida (Brébisson ex Kützing) Van Heurck | 0.913 | 0.944 | 0.929 | 0.005 |
| Cymbella turgidula Grunow | 0.862 | 0.889 | 0.875 | 0.005 |
| Cymbella peraffinis Tynni | 0.866 | 0.778 | 0.821 | 0.005 |
| Encyonema lacustre (Agardh) Mann | 0.804 | 0.778 | 0.791 | 0.005 |
| Cymbella saxicola Bíly & Marvan | 1.000 | 0.611 | 0.782 | 0.005 |
| Navicula antonii Lange-Bertalot & Rumrich | 0.551 | 1.000 | 0.742 | 0.005 |
| Luticola stigma (Patrick) Johansen | 0.990 | 0.556 | 0.742 | 0.005 |
| Nitzschia dissipata (Kützing) Rabenhorst | 0.538 | 1.000 | 0.733 | 0.005 |
| Amphiprora paludosa Smith | 0.942 | 0.500 | 0.686 | 0.005 |
| Gomphonema subclavatum (Grunow) Grunow | 0.533 | 0.722 | 0.621 | 0.015 |
| Gyrosigma scalproides (Rabenhorst) Cleve | 0.510 | 0.722 | 0.607 | 0.030 |
| Amphora bullatoides Hohn & Hellerman | 0.702 | 0.500 | 0.592 | 0.020 |
| Navicula cryptotenella Lange-Bertalot | 0.560 | 0.556 | 0.558 | 0.020 |
| Navicula cryptofallax Lange-Bertalot & Hofmann | 0.632 | 0.444 | 0.530 | 0.015 |
| Navicula rostellata Kützing | 0.903 | 0.278 | 0.501 | 0.010 |
| G4 (N = 8) | ||||
| Planothidium lanceolatum (Brébisson) Lange-Bertalot | 0.658 | 1.000 | 0.811 | 0.005 |
| Halamphora montana (Krasske) Levkov | 0.910 | 0.625 | 0.754 | 0.005 |
| Navicula reichardtiana Lange-Bertalot | 0.638 | 0.875 | 0.747 | 0.005 |
| Nitzschia palea (Kützing) Smith | 0.624 | 0.875 | 0.739 | 0.010 |
| Navicula symmetrica Patrick | 0.658 | 0.750 | 0.703 | 0.005 |
| Amphora cymbamphora Cholnoky | 0.708 | 0.625 | 0.665 | 0.005 |
| Nitzschia valdecostata Lange-Bertalot & Simonsen | 0.822 | 0.375 | 0.555 | 0.005 |
| Nitzschia minuta Bleisch | 0.606 | 0.500 | 0.550 | 0.020 |
| G5 (N = 1) | ||||
| Cocconeis placentula Ehrenberg | 0.348 | 1.000 | 0.590 | 0.010 |
| Month | Diatom Indices | Predictors | Coeff. | p-Value | F Statistic | p-Value | R2adj |
|---|---|---|---|---|---|---|---|
| April | SLA | Intercept | 11.422 | <0.0001 | 5.624 | 0.0074 | 0.192 |
| NH4–N * | −5.793 | 0.0071 | |||||
| Velocity * | 1.578 | 0.0268 | |||||
| IBD | Intercept | 4.219 | 0.0002 | 9.875 | 0.0032 | 0.185 | |
| Elevation | 0.004 | 0.0032 | |||||
| TDI | Intercept | 58.140 | <0.0001 | 7.029 | 0.0116 | 0.134 | |
| Elevation | −0.012 | 0.0116 | |||||
| September | SLA | Intercept | 23.440 | 0.0002 | 8.325 | <0.0001 | 0.429 |
| TP * | −27.330 | <0.0001 | |||||
| pH | −1.438 | 0.0353 | |||||
| Elevation | −0.001 | 0.0215 | |||||
| Velocity * | 3.852 | 0.0068 | |||||
| TDI | Intercept | 54.574 | <0.0001 | 4.107 | 0.0498 | 0.074 | |
| Elevation | −0.008 | 0.0498 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, S.; Guo, S.; Deng, H.; Xie, Z.; Tang, T. Recognition of Patterns of Benthic Diatom Assemblages within a River System to Aid Bioassessment. Water 2018, 10, 1559. https://doi.org/10.3390/w10111559
Mao S, Guo S, Deng H, Xie Z, Tang T. Recognition of Patterns of Benthic Diatom Assemblages within a River System to Aid Bioassessment. Water. 2018; 10(11):1559. https://doi.org/10.3390/w10111559
Chicago/Turabian StyleMao, Shuxin, Shuhan Guo, Hongbing Deng, Zhicai Xie, and Tao Tang. 2018. "Recognition of Patterns of Benthic Diatom Assemblages within a River System to Aid Bioassessment" Water 10, no. 11: 1559. https://doi.org/10.3390/w10111559
APA StyleMao, S., Guo, S., Deng, H., Xie, Z., & Tang, T. (2018). Recognition of Patterns of Benthic Diatom Assemblages within a River System to Aid Bioassessment. Water, 10(11), 1559. https://doi.org/10.3390/w10111559