Abundance of Biological Ice Nucleating Particles in the Mississippi and Its Major Tributaries


Abstract
1. Introduction
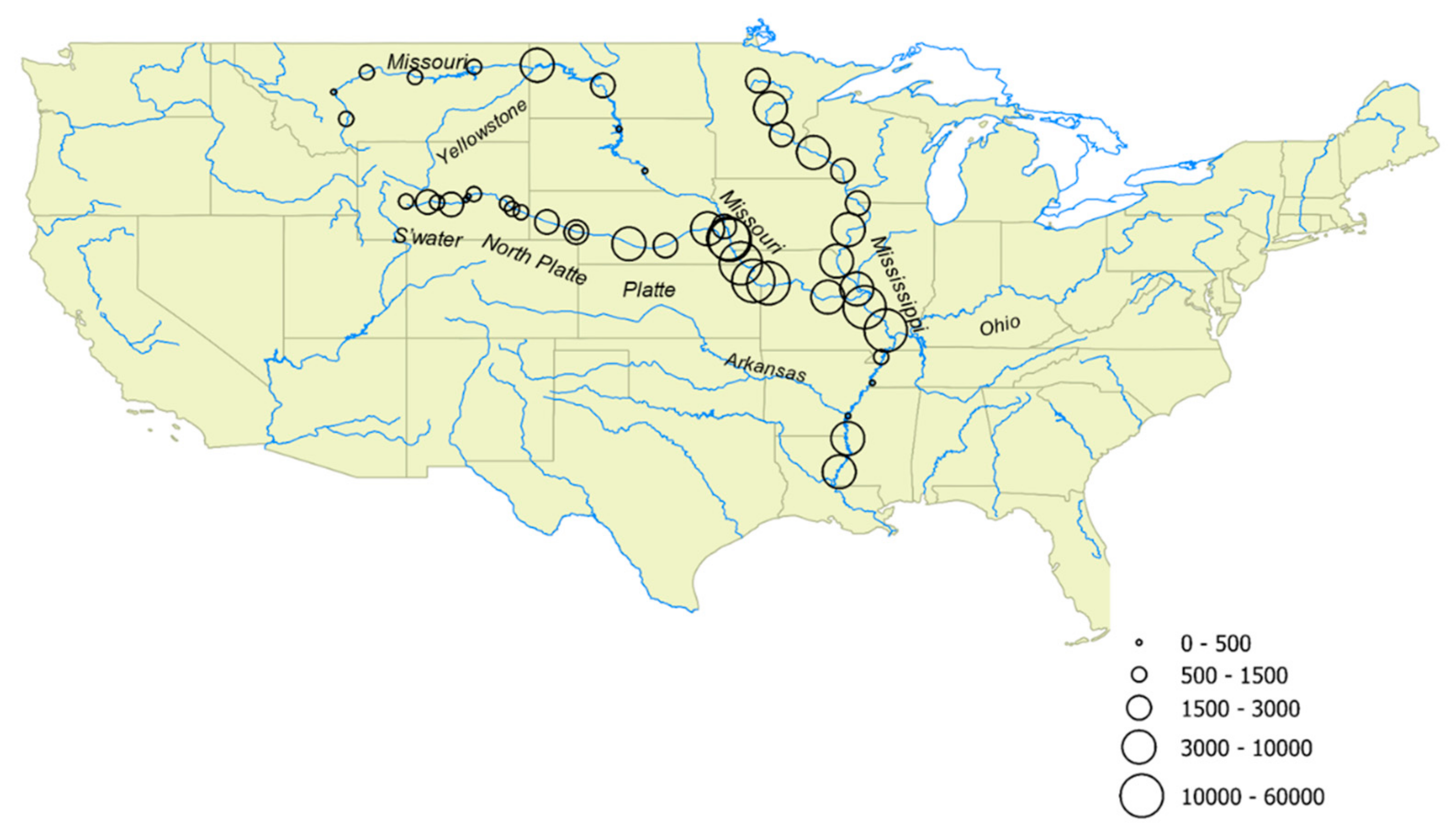
2. Materials and Methods
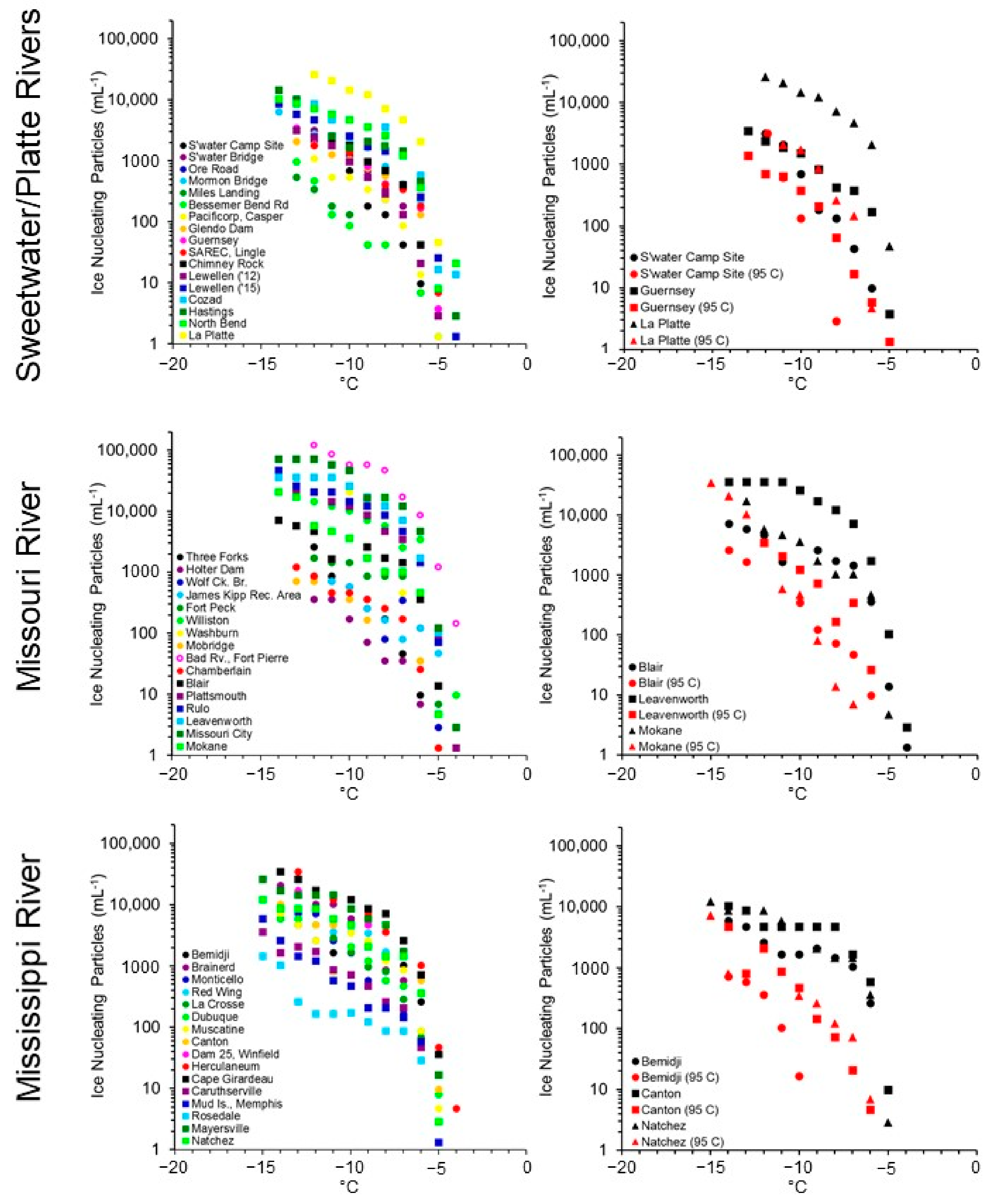
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Petters, M.D.; Wright, T.P. Revisiting ice nucleation from precipitation samples. Geophys. Res. Lett. 2015, 42, 8758–8766. [Google Scholar] [CrossRef]
- Lohmann, U.; Feichter, J. Global indirect aerosol effects: A review. Atmos. Chem. Phys. 2005, 5, 715–737. [Google Scholar] [CrossRef]
- DeMott, P.J.; Prenni, J.A.; Liu, X.; Kreidenweis, S.M.; Petters, M.D.; Twohy, C.H.; Richardson, M.S.; Eidhammer, T.; Rogers, D.C. Predicting global atmospheric ice nuclei distributions and their impacts on climate. Proc. Natl. Acad. Sci. USA 2010, 107, 11217–11222. [Google Scholar] [CrossRef] [PubMed]
- Murray, B.J.; O’Sullivan, D.; Atkinson, J.D.; Webb, M. Ice nucleation by particles immersed in supercooled cloud droplets. Chem. Soc. Rev. 2012, 41, 6519–6554. [Google Scholar] [CrossRef] [PubMed]
- Boucher, O.; Randall, D.; Artaxo, P.; Bretherton, C.; Feingold, G.; Forster, P.; Kerminen, V.-M.; Kondo, Y.; Liao, H.; Lohmann, U.; et al. Clouds and Aerosols. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Hoose, C.; Möhler, O. Heterogeneous ice nucleation on atmospheric aerosols: A review of results from laboratory experiments. Atmos. Chem. Phys. 2012, 12, 9817–9854. [Google Scholar] [CrossRef]
- Niemand, M.; Möhler, O.; Vogel, B.; Vogel, H.; Hoose, C.; Connolly, P.; Klein, H.; Bingemer, H.; DeMott, P.; Skrotzki, J.; et al. Parameterization of immersion freezing on mineral dust particles: An application in a regional scale model. J. Atmos. Sci. 2012, 69, 3077–3092. [Google Scholar] [CrossRef]
- Boose, Y.; Welti, A.; Atkinson, J.; Ramelli, F.; Danielczok, A.; Bingemer, H.G.; Plötze, M.; Sierau, B.; Kanji, Z.A.; Lohmann, U. Heterogeneous ice nucleation on dust particles sourced from nine deserts worldwide—Part 1: Immersion freezing. Atmos. Chem. Phys. 2016, 16, 15075–15095. [Google Scholar] [CrossRef]
- Conen, F.; Morris, C.E.; Leifeld, J.; Yakutin, M.V.; Alewell, C. Biological residues define the ice nucleation properties of soil dust. Atmos. Chem. Phys. 2011, 11, 9643–9648. [Google Scholar] [CrossRef]
- O’Sullivan, D.; Murray, B.J.; Malkin, T.L.; Whale, T.F.; Umo, N.S.; Atkinson, J.D.; Price, H.C.; Baustian, K.J.; Browse, J.; Webb, M.E. Ice nucleation by fertile soil dusts: Relative importance of mineral and biogenic components. Atmos. Chem. Phys. 2014, 14, 1853–1867. [Google Scholar]
- Tobo, Y.; DeMott, P.J.; Hill, T.C.J.; Prenni, A.J.; Swoboda-Colberg, N.G.; Franc, G.D.; Kreidenweis, S.M. Organic matter matters for ice nuclei of agricultural soil origin. Atmos. Chem. Phys. 2014, 14, 8521–8531. [Google Scholar] [CrossRef]
- Hill, T.C.J.; DeMott, P.J.; Tobo, Y.; Fröhlich-Nowoisky, J.; Moffett, B.F.; Franc, G.D.; Kreidenweis, S.M. Sources of organic ice nucleating particles in soils. Atmos. Chem. Phys. 2016, 16, 7195–7211. [Google Scholar] [CrossRef]
- Schnell, R.C.; Vali, G. Biogenic ice nuclei, Part I: Terrestrial and marine sources. J. Atmos. Sci. 1976, 33, 1554–1564. [Google Scholar] [CrossRef]
- Vali, G.; Christensen, M.; Fresh, R.W.; Galyan, E.L.; Maki, L.R.; Schnell, R.C. Biogenic ice nuclei Part II, Bacterial sources. J. Atmos. Sci. 1976, 33, 1565–1570. [Google Scholar]
- Burrows, S.M.; Hoose, C.; Pöschl, U.; Lawrence, M.G. Ice nuclei in marine air: Bioparticles or dust? Atmos. Chem. Phys. 2011, 13, 245–267. [Google Scholar] [CrossRef]
- Wang, X.; Sultana, C.M.; Trueblood, J.; Hill, T.C.J.; Malfatti, F.; Lee, C.; Laskina, O.; Moore, K.A.; Beall, C.M.; McCluskey, C.S.; et al. Microbial control of sea spray aerosol composition: A tale of two blooms. ACS Cent. Sci. 2015, 1, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.W.; Ladino, L.A.; Alpert, P.A.; Breckels, M.N.; Brooks, I.M.; Browse, J.; Burrows, S.M.; Carslaw, K.S.; Huffman, J.A.; Judd, C.; et al. A marine biogenic source of atmospheric ice-nucleating particles. Nature 2015, 525, 234–238. [Google Scholar] [CrossRef] [PubMed]
- DeMott, P.J.; Hill, T.C.J.; McCluskey, C.S.; Prather, K.A.; Collins, D.B.; Sullivan, R.C.; Ruppel, M.J.; Mason, R.H.; Irish, V.E.; Lee, T.; et al. Sea spray aerosol as a unique source of ice nucleating particles. Proc. Natl. Acad. Sci. USA 2016, 113, 5797–5803. [Google Scholar] [CrossRef] [PubMed]
- McCluskey, C.S.; Hill, T.C.J.; Malfatti, F.; Sultana, C.M.; Lee, C.; Santander, M.V.; Beall, C.M.; Moore, K.A.; Cornwell, G.C.; Collins, D.B.; et al. A dynamic link between ice nucleating particles released in nascent sea spray aerosol and oceanic biological activity during two mesocosm experiments. J. Atmos. Sci. 2017, 74, 151–166. [Google Scholar] [CrossRef]
- McCluskey, C.S.; Hill, T.C.J.; Sultana, C.M.; Laskina, O.; Trueblood, J.; Santander, M.V.; Beall, C.M.; Michaud, J.M.; Kreidenweis, S.M.; Prather, K.A.; et al. A mesocosm double feature: Insights into the chemical make-up of marine ice nucleating particles. J. Atmos. Sci. 2018, 75, 2405–2423. [Google Scholar] [CrossRef]
- Vergara-Temprado, J.; Miltenberger, A.K.; Furtado, K.; Grosvenor, D.P.; Shipway, B.J.; Hill, A.A.; Wilkinson, J.M.; Field, P.R.; Murray, B.J.; Carslaw, K.S. Strong control of Southern Ocean cloud reflectivity by ice-nucleating particles. P. Natl. Acad. Sci. 2018, 115, 2687–2692. [Google Scholar] [CrossRef] [PubMed]
- Bigg, E.K. Ice nucleus concentrations in remote areas. J. Atmos. Sci. 1973, 30, 1153–1157. [Google Scholar] [CrossRef]
- McCluskey, C.S.; Ovadnevaite, J.; Rinaldi, M.; Atkinson, J.; Belosi, F.; Ceburnis, D.; Salvatore, M.; Hill, T.C.J.; Lohmann, U.; Kanji, Z.A.; et al. Marine and terrestrial organic ice-nucleating particles in pristine marine to continentally influenced Northeast Atlantic air masses. J. Geophys. Res. Atmos. 2018, 123. [Google Scholar] [CrossRef]
- Schnell, R.C.; Vali, G. Freezing nuclei in marine waters. Tellus 1975, 27, 321–323. [Google Scholar] [CrossRef]
- Schnell, R. Ice nuclei in seawater, fog water and marine air off the coast of Nova Scotia: Summer 1975. J. Atmos. Sci. 1977, 34, 1299–1305. [Google Scholar] [CrossRef]
- Engel, A.; Bange, H.W.; Cunliffe, M.; Burrows, S.M.; Friedrichs, G.; Galgani, L.; Herrmann, H.; Hertkorn, N.; Johnson, M.; Liss, P.S.; et al. The ocean’s vital skin: Toward an integrated understanding of the sea surface microlayer. Front. Mar. Sci. 2017, 4, 165. [Google Scholar] [CrossRef]
- Morris, C.E.; Sands, D.C.; Vinatzer, B.A.; Glaux, C.; Guilbaud, C.; Buffière, A.; Yan, S.; Dominguez, H.; Thompson, B.M. The life history of the plant pathogen Pseudomonas syringae is linked to the water cycle. Int. Soc. Microb. Ecol. 2008, 2, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.E.; Sands, D.C.; Vanneste, J.L.; Montarry, J.; Oakley, B.; Guilbaud, C.; Glaux, C. Inferring the evolutionary history of the plant pathogen Pseudomonas syringae from its biogeography in headwaters of rivers in North America, Europe, and New Zealand. Am. Soc. Microbol. 2010, 1, 1–11. [Google Scholar] [CrossRef] [PubMed]
- D’souza, N.A.; Kawarasaki, Y.; Gantz, J.D.; Lee, R.D., Jr.; Beall, B.F.N.; Shtarkman, Y.M.; Koçer, Z.A.; Rogers, S.O.; Wildschutte, H.; Bullerjahn, G.S.; et al. Diatom assemblages promote ice formation in large lakes. Int. Soc. Microb. Ecol. 2013, 7, 1632–1640. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, R.B.; David, R.F.; Marr, L.C.; Vinatzer, B.; Schmale, D.G., III. Aerosolization of two strains (Ice+ and Ice–) of Pseudomonas syringae in a collision nebulizer at different temperatures. Aerosol Sci. Technol. 2015, 49, 159–166. [Google Scholar] [CrossRef]
- Moffett, B.F. Fresh water ice nuclei. Fundam. Appl. Limnol. 2016, 188, 19–23. [Google Scholar] [CrossRef]
- Larsen, J.A.; Conen, F.; Alewell, C. Export of ice nucleating particles from a watershed. R. Soc. Open Sci. 2017, 4, 170213. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.C.J.; Georgakopoulos, D.G.; DeMott, P.J.; Stump, W.L.; Franc, G.D. Measurement of ice nucleation-active bacteria on plants and in precipitation by quantitative PCR. Appl. Environ. Microbiol. 2014, 80, 256–1267. [Google Scholar] [CrossRef] [PubMed]
- Vali, G. Quantitative evaluation of experimental results on the heterogeneous freezing nucleation of supercooled liquids. J. Atmos. Sci. 1971, 28, 402–409. [Google Scholar] [CrossRef]
- Vali, G. Sizes of atmospheric ice nuclei. Nature 1966, 212, 384–385. [Google Scholar] [CrossRef]
- Rangel-Alvarado, R.B.; Nazarenko, Y.; Ariya, P.A. Snow-borne nanosized particles: Abundance, distribution, composition, and significance in ice nucleation processes. J. Geophys. Res. Atmos. 2015, 120, 11760–11774. [Google Scholar] [CrossRef]
- Šantl-Temkiv, T.; Sahyoun, M.; Finster, K.; Hartmann, S.; Augustin, S.; Stratmann, F.; Wex, H.; Clauss, T.; Nielsen, N.W.; Sørensen, J.H.; et al. Characterization of airborne ice-nucleation-active bacteria and bacterial fragments. Atmos. Environ. 2015, 109, 105–117. [Google Scholar] [CrossRef]
- Fröhlich-Nowoisky, J.; Hill, T.C.J.; Pummer, B.G.; Yordanova, P.; Franc, G.D.; Pöschl, U. Ice nucleation activity in the widespread soil fungus Mortierella alpina. Biogeosciences 2015, 12, 1057–1071. [Google Scholar] [CrossRef]
- Pummer, B.G.; Budke, C.; Augustin-Bauditz, S.; Niedermeier, D.; Felgitsch, L.; Kampf, C.J.; Huber, R.G.; Liedl, K.R.; Loerting, T.; Moschen, T.; et al. Ice nucleation by water-soluble macromolecules. Atmos. Chem. Phys. 2015, 15, 4077–4091. [Google Scholar] [CrossRef]
- O’Sullivan, D.; Murray, B.J.; Ross, J.; Webb, M.E. The adsorption of fungal ice-nucleating proteins on mineral dusts: A terrestrial reservoir of atmospheric ice-nucleating particles. Atmos. Chem. Phys. 2016, 16, 7879–7887. [Google Scholar]
- DeLeon-Rodriguez, N.; Lathem, T.L.; Rodriguez-R, L.M.; Barazesh, J.M.; Anderson, B.E.; Beyersdorf, A.J.; Ziemba, L.D.; Bergin, M.; Nenes, A.; Konstantinidis, K.T. Microbiome of the upper troposphere: Species composition and prevalence, effects of tropical storms, and atmospheric implications. Proc. Natl. Acad. Sci. USA 2013, 110, 2575–2580. [Google Scholar] [CrossRef] [PubMed]
- Maki, T.; Hara, K.; Iwata, A.; Lee, K.C.; Kawai, K.; Kai, K.; Kobayashi, F.; Pointing, S.B.; Archer, S.; Hasegawa, H.; et al. Variations in airborne bacterial communities at high altitudes over the Noto Peninsula (Japan) in response to Asian dust events. Atmos. Chem. Phys. 2017, 17, 11877–11897. [Google Scholar] [CrossRef]
- Pruppacher, H.R.; Klett, J.D. Microphysics of Clouds and Precipitation; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1997. [Google Scholar]
- Leifer, I.; Caulliez, G.; de Leeuw, G. Bubbles generated from wind-steepened breaking waves: 2. Bubble plumes, bubbles, and wave characteristics. J. Geophys. Res. Oceans 2006, 111, C06021. [Google Scholar] [CrossRef]
- Axson, J.L.; May, N.W.; Coloón-Bernal, I.D.; Pratt, K.A.; Ault, A.P. Lake spray aerosol: A chemical signature from individual ambient particles. Environ. Sci. Technol. 2016, 50, 9835–9845. [Google Scholar] [CrossRef] [PubMed]
- Knackstedt, K.A.; Moffett, B.F.; Hartmann, S.; Wex, H.; Hill, T.; Glasgo, E.; Reitz, L.; Augustiin-Bauditz, S.; Deustcher, W.H.; Beall, B.; et al. A terrestrial origin for abundant riverine nanoscale ice-nucleating particles. Environ. Sci. Technol. 2018. submitted. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| River | Location | Date | Lat. | Long. | Water Temp. (°C) | pH | Onset of Freezing (°C) | INPs Active at −7 °C (mL−1) | INPs Active at −10 °C (mL−1) | INPs Heat Labile at −10 °C (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| Sweetwater | Sweetwater Camp Site | 8/15/2012 | 42.564837 | −109.061902 | 11.0 | 6.8 | −5 | 43 | 690 | 81 |
| Sweetwater | Sweetwater Bridge | 8/15/2012 | 42.541721 | −108.18018 | 17.0 | 7.7 | −6 | 180 | 1600 | 99 |
| Sweetwater | Ore Road | 8/15/2012 | 42.524983 | −107.798983 | 21.0 | 8.0 | −6 | 130 | 1200 | 66 |
| Sweetwater | Mormon Bridge | 8/14/2012 | 42.442715 | −107.221725 | - | - | −5 | 420 | 2100 | 78 |
| North Platte | Miles Landing | 8/16/2012 | 42.638502 | −106.619033 | 17.0 | 7.3 | −6 | >69 | 130 | 67 |
| North Platte | Bessemer Bend Rd | 8/16/2012 | 42.771948 | −106.530563 | 17.0 | 7.7 | −5 | >69 | 87 | 89 |
| North Platte | Pacificorp, Casper | 8/16/2012 | 42.865464 | −106.297231 | 19.0 | 8.4 | −4 | 87 | 540 | 76 |
| North Platte | Glendo Dam | 8/17/2012 | 42.478084 | −104.951656 | 17.0 | 7.3 | −4 | 370 | 1100 | 66 |
| North Platte | Guernsey | 8/17/2012 | 42.259881 | −104.74157 | 20.0 | 8.6 | −5 | 370 | 1500 | 75 |
| North Platte | SAREC, Lingle | 8/23/2012 | 42.125186 | −104.378219 | 19.0 | 7.6 | −5 | 340 | 1400 | 87 |
| North Platte | Chimney Rock | 8/24/2012 | 41.729665 | −103.324214 | 18.0 | 7.5 | −5 | 410 | 1800 | 95 |
| North Platte | Lewellen | 8/24/2012 | 41.316284 | −102.126333 | 21.0 | 7.5 | −5 | 130 | 980 | 91 |
| North Platte | Lewellen | 6/14/2015 | 41.316284 | −102.126333 | 21.5 | 8.0 | −4 | 1500 | 2600 | 69 |
| Platte | Cozad | 6/14/2015 | 40.837433 | −99.98696 | 22.5 | 8.0 | −5 | 1500 | 4700 | 99 |
| Platte | Hastings | 6/14/2015 | 40.77126 | −98.492202 | 24.0 | 8.1 | −5 | 1500 | 1700 | 94 |
| Platte | North Bend | 6/14/2015 | 41.452678 | −96.776912 | 24.5 | 8.3 | −5 | 1200 | 4700 | 97 |
| Platte | La Platte | 6/15/2015 | 41.058924 | −95.928276 | - | - | −5 | 4700 | 15,000 | 88 |
| Missouri | Three Forks/Headwaters St. Pk. | 6/28/2015 | 45.926835 | −111.50578 | 22.1 | 8.6 | −6 | 47 | 590 | - |
| Missouri | Wolf Creek Br./Holter Dam | 6/27/2015 | 47.020074 | −112.011922 | 17.3 | 9.0 | −6 | 36 | 170 | - |
| Missouri | Old Fort Benton | 6/27/2015 | 47.820359 | −110.663314 | 24.3 | 8.9 | −4 | 350 | 590 | - |
| Missouri | James Kipp Recreation Area | 6/27/2015 | 47.627895 | −108.697475 | 27.5 | 8.8 | −5 | 81 | 590 | - |
| Missouri | Fort Peck | 6/26/2015 | 48.039842 | −106.290658 | 12.9 | 8.3 | −5 | 870 | 1500 | - |
| Missouri | Williston | 6/26/2015 | 48.110958 | −103.717126 | 21.5 | 8.1 | −4 | 2600 | 10,000 | - |
| Missouri | Washburn | 6/26/2015 | 47.288804 | −101.042415 | 11.0 | 8.4 | −5 | 470 | 2100 | - |
| Missouri | Mobridge | 6/25/2015 | 45.512826 | −100.382602 | 24.2 | 7.3 | −5 | 170 | 360 | - |
| Missouri | Bad River near Fort Pierre | 6/25/2015 | 44.353082 | −100.369376 | 23.2 | 7.8 | −5 | 17,000 | 59,000 | - |
| Missouri | Chamberlain | 6/25/2015 | 43.815016 | −99.326346 | 21.3 | 8.5 | −5 | 170 | 470 | - |
| Missouri | Blair | 6/14/2015 | 41.548381 | −96.097105 | 22.3 | 8.4 | −4 | 1500 | 3000 | 90 |
| Missouri | Plattsmouth | 6/15/2015 | 41.000821 | −95.866735 | 23.2 | 7.9 | −4 | 3500 | 12,000 | 95 |
| Missouri | Rulo | 6/15/2015 | 40.049925 | −95.421274 | 23.0 | 7.9 | −5 | 4700 | 15,000 | 93 |
| Missouri | Leavenworth | 6/15/2015 | 39.330756 | −94.910311 | 23.5 | 7.7 | −4 | 7200 | 26,000 | 95 |
| Missouri | Missouri City | 6/15/2015 | 39.236795 | −94.29664 | 22.7 | 7.7 | −4 | 12,000 | 47,000 | 95 |
| Missouri | Mokane | 6/17/2015 | 38.670188 | −91.874438 | 20.8 | 7.9 | −5 | 1000 | 3600 | 90 |
| Mississippi | Bemidji | 6/23/2015 | 47.483244 | −94.727823 | 20.3 | 8.0 | −6 | 1000 | 1700 | 99 |
| Mississippi | Brainerd | 6/23/2015 | 46.34847 | −94.20721 | 22.7 | 7.9 | −5 | 590 | 5900 | 99 |
| Mississippi | Monticello | 6/22/2015 | 45.294468 | −93.757178 | 25.2 | 8.2 | −5 | 170 | 1700 | 97 |
| Mississippi | Red Wing | 6/22/2015 | 44.562502 | −92.461088 | 23.9 | 8.4 | −5 | 1500 | 3600 | 96 |
| Mississippi | Pettibone Pk, La Crosse | 6/22/2015 | 43.812197 | −91.260732 | 24.7 | 8.4 | −6 | 290 | 1700 | 95 |
| Mississippi | Dubuque | 6/22/2015 | 42.495042 | −90.65702 | 23.2 | 8.1 | −6 | 470 | 2100 | 93 |
| Mississippi | Muscatine | 6/21/2015 | 41.419779 | −91.043757 | 24.4 | 7.9 | −5 | 870 | 3500 | 93 |
| Mississippi | Canton | 6/21/2015 | 40.139674 | −91.515404 | 23.9 | 8.0 | −5 | 1700 | 4700 | 90 |
| Mississippi | Dam 25, Winfield | 6/21/2015 | 39.0054 | −90.691467 | 24.0 | 8.0 | −5 | 2600 | 8700 | 99 |
| Mississippi | Herculaneum | 6/17/2015 | 38.258919 | −90.376007 | 22.8 | 7.7 | −4 | 2600 | 12,000 | 99 |
| Mississippi | Cape Girardeau | 6/17/2015 | 37.31516 | −89.512691 | 25.8 | 7.7 | −5 | 2600 | 12,000 | 97 |
| Mississippi | Caruthersville | 6/17/2015 | 36.232633 | −89.702174 | 28.0 | 8.6 | −5 | 210 | 720 | 76 |
| Mississippi | Mud Island, Memphis | 6/18/2015 | 35.18217 | −90.057939 | 28.6 | 7.4 | −5 | 150 | 470 | 92 |
| Mississippi | Rosedale | 6/18/2015 | 33.827979 | −91.053996 | 27.1 | 7.8 | −6 | 87 | 170 | 90 |
| Mississippi | Mayersville | 6/18/2015 | 32.917046 | −91.061286 | 26.8 | 7.7 | −5 | 1700 | 8700 | 96 |
| Mississippi | Natchez | 6/18/2015 | 31.558579 | −91.413607 | 26.6 | 7.7 | −5 | 1500 | 4700 | 93 |
| Samples | INPs < 0.22 µm Active at −7 °C (%) | INPs < 0.22 µm Active at −10 °C (%) |
|---|---|---|
| Sweetwater (Campsite, Bridge, Ore Road) | 100 | 97 |
| North Platte (Glendo Dam, Guernsey) | 31 | 32 |
| North Platte (SAREC, Lingle) | 44 | 24 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moffett, B.F.; Hill, T.C.J.; DeMott, P.J. Abundance of Biological Ice Nucleating Particles in the Mississippi and Its Major Tributaries. Atmosphere 2018, 9, 307. https://doi.org/10.3390/atmos9080307
Moffett BF, Hill TCJ, DeMott PJ. Abundance of Biological Ice Nucleating Particles in the Mississippi and Its Major Tributaries. Atmosphere. 2018; 9(8):307. https://doi.org/10.3390/atmos9080307
Chicago/Turabian StyleMoffett, Bruce F., Tom C. J. Hill, and Paul J. DeMott. 2018. "Abundance of Biological Ice Nucleating Particles in the Mississippi and Its Major Tributaries" Atmosphere 9, no. 8: 307. https://doi.org/10.3390/atmos9080307
APA StyleMoffett, B. F., Hill, T. C. J., & DeMott, P. J. (2018). Abundance of Biological Ice Nucleating Particles in the Mississippi and Its Major Tributaries. Atmosphere, 9(8), 307. https://doi.org/10.3390/atmos9080307