
Assessment of Suppressive Effects of Negative Air Ions on Fungal Growth, Sporulation and Airborne Viral Load

 ,
,  , , , , , , , , ,
, , , , , , , , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Evaluation of the Effect of NAIs on Airborne Fungal Spores
2.1.1. Air Sampling
2.1.2. Determination of Spore Counts in ITS and NTS Air Samples and Evaluation of NAIs Effects on Fungal Growth and Sporulation
2.2. Evaluation of the Effect of NAIs on EHV-1
2.2.1. Experimental Setup
2.2.2. DNA Extraction, EHV-1 Detection and Quantitation
3. Statistical Analysis
4. Results
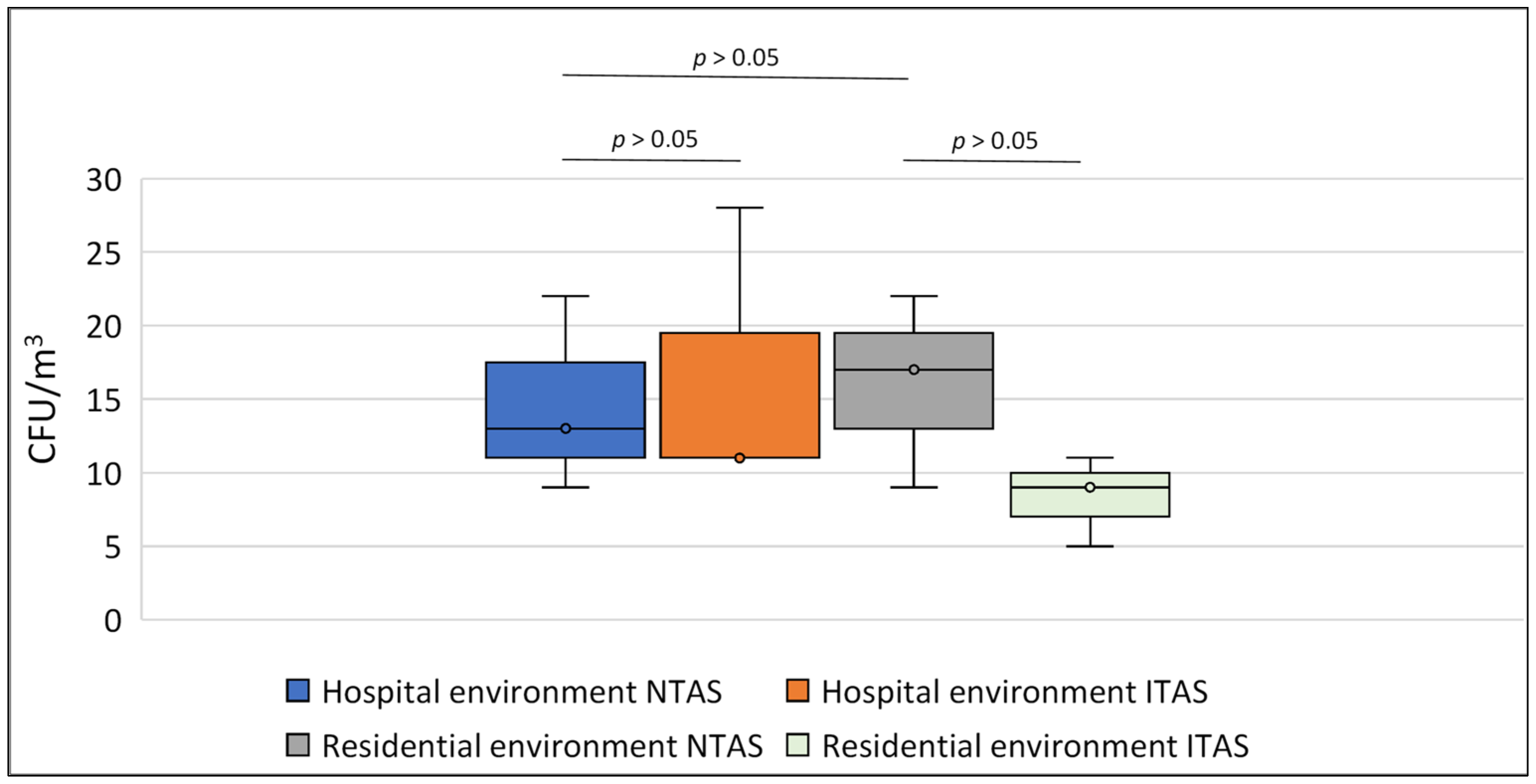
4.1. Effect of NAIs on Airborne Fungal Spores
4.2. Effect of NAIs on Aerosolized EHV-1
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, A.H.; Martin, S.; Phan, B.; Benigno, M.; Stephens, J.; Chambers, R.; Aram, J.A. Patient characteristics and risk factors in invasive mold infections: Comparison from a systematic review and database analysis. Clinicoecon. Outcomes Res. 2021, 13, 593–602. [Google Scholar] [CrossRef]
- Kraft, S.; Buchenauer, L.; Polte, T. Mold, mycotoxins and a dysregulated immune system: A combination of concern? Int. J. Mol. Sci. 2021, 22, 12269. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.; Oliveira, D.; Lisboa, C.; Boechat, J.L.; Delgado, L. Clinical manifestations of human exposure to fungi. J. Fungi 2023, 9, 381. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.O. Diagnosis and treatment of invasive mold diseases. Infect. Chemother. 2023, 55, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Paiva Macedo, J.; Dias, V.C. Antifungal resistance: Why are we losing this battle? Future Microbiol. 2024, 19, 1027–1040. [Google Scholar] [CrossRef]
- Yun, H.; Seo, J.H.; Kim, Y.G.; Yang, J. Impact of scented candle use on indoor air quality and airborne microbiome. Sci. Rep. 2025, 15, 10181. [Google Scholar] [CrossRef]
- Azuma, K.; Yanagi, U.; Kagi, N.; Kim, H.; Ogata, M.; Hayashi, M. Environmental factors involved in SARS-CoV-2 transmission: Effect and role of indoor environmental quality in the strategy for COVID-19 infection control. Environ. Health Prev. Med. 2020, 25, 66. [Google Scholar] [CrossRef]
- Rawat, M.S.; Roberts, A.D.; Brown, D.M.; Ferro, A.R. Resuspension of Seeded Particles Containing Live Influenza A Virus in a Full-Scale Laboratory. Buildings 2023, 13, 1734. [Google Scholar] [CrossRef]
- van Doremalen, N.; Bushmaker, T.; Morris, D.H.; Holbrook, M.G.; Gamble, A.; Williamson, B.N.; Tamin, A.; Harcourt, J.L.; Thornburg, N.J.; Gerber, S.I.; et al. Aerosol and Surface Stability of SARS-CoV-2 as Compared with SARS-CoV-1. N. Engl. J. Med. 2020, 382, 1564–1567. [Google Scholar] [CrossRef]
- Morawska, L. Droplet fate in indoor environments, or can we prevent the spread of infection? Indoor Air 2006, 16, 335–347. [Google Scholar] [CrossRef]
- Feng, Z.; Cao, S.-J.; Haghighat, F. Removal of SARS-CoV-2 Using UV+Filter in Built Environment. Sustain. Cities Soc. 2021, 74, 103226. [Google Scholar] [CrossRef]
- Park, D.H.; An, S.-H.; Lee, Y.; Kim, Y.-J.; Han, B.; Kim, H.-J. Development of On-Demand Antiviral Electrostatic Precipitators with Electrothermal-Based Antiviral Surfaces against Airborne Virus Particles. Toxics 2022, 10, 601. [Google Scholar] [CrossRef]
- Grinshpun, S.A.; Mainelis, G.; Trunov, M.; Adhikari, A.; Reponen, T.; Willeke, K. Evaluation of Ionic Air Purifiers for Reducing Aerosol Exposure in Confined Indoor Spaces. Indoor Air 2005, 15, 235–245. [Google Scholar] [CrossRef]
- Claus, H. Ozone generation by ultraviolet lamps. Photochem. Photobiol. 2021, 97, 471–476. [Google Scholar] [CrossRef]
- Hashimoto, K.; Kawakami, Y. Effectiveness of airborne fungi removal by using a HEPA air purifier fan in houses. Biocontrol Sci. 2018, 23, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Curtis, L. HEPA filters and airborne viruses, bacteria, and fungi. Otolaryngol. Head Neck Surg. 2022, 166, 1005. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, L.A.; Gaunt, L.F.; Beggs, C.B.; Shepherd, S.J.; Sleigh, P.A.; Noakes, C.J.; Kerr, K.G. Bactericidal action of positive and negative ions in air. BMC Microbiol. 2007, 7, 32. [Google Scholar] [CrossRef]
- Li, J.; Ma, C.; Zhu, S.; Yu, F.; Dai, B.; Yang, D. A review of recent advances of dielectric barrier discharge plasma in catalysis. Nanomaterials 2019, 9, 1428. [Google Scholar] [CrossRef]
- Altamimi, G.; Illias, H.A.; Mokhtar, N.; Mokhlis, H.; Bakar, A.H.A. Corona discharges under various types of electrodes. In Proceedings of the IEEE International Conference on Power and Energy (PECon), Kuching, Malaysia, 1–3 December 2014. [Google Scholar]
- Radalj, A.; Nikšić, A.; Trajković, J.; Knezević, T.; Janković, M.; De Luka, S.; Djoković, S.; Mijatović, S.; Ilić, A.; Arandjelović, I.; et al. Combating Pathogens Using Carbon-Fiber Ionizers (CFIs) for Air Purification: A Narrative Review. Appl. Sci. 2024, 14, 7311. [Google Scholar] [CrossRef]
- Panich, I.; Tippayawong, N. Effect of needle cone angle and air flow rate on electrostatic discharge characteristics of a corona-needle ionizer. J. Electrostat. 2010, 68, 254–260. [Google Scholar]
- Han, B.; Kim, H.J.; Kim, Y.J.; Sioutas, C. Unipolar charging of fine and ultra-fine particles using carbon fiber ionizers. Aerosol Sci. Technol. 2008, 42, 793–800. [Google Scholar] [CrossRef]
- Kolarž, P.; Ilić, A.Ž.; Janković, M.; Janićijević, A.; Trbovich, A.M. Estimating aerosol particle removal in indoor air by ion-enhanced deposition. J. Aerosol Sci. 2023, 173, 106199. [Google Scholar] [CrossRef]
- Yang, G.; Niu, B.; Zong, Z.; Wu, W.; Fang, X.; Chen, H.; Zhang, Y.; Mu, H.; Gao, H. Microbicidal effect of negative air ion against Penicillium citrinum and quality control of Chinese bayberry. Food Control. 2024, 162, 110476. [Google Scholar] [CrossRef]
- Pratt, R.; Barnard, R.W. Some effects of ionized air on Penicillium notatum. J. Am. Pharm. Assoc. Sci. Ed. 1960, 49, 643–646. [Google Scholar] [CrossRef]
- Pan, M.; Lednicky, J.A.; Wu, C.Y. Collection, Particle Sizing and Detection of Airborne Viruses. J. Appl. Microbiol. 2019, 127, 1596–1611. [Google Scholar] [CrossRef] [PubMed]
- Honsig, C.; Selitsch, B.; Hollenstein, M.; Vossen, M.G.; Spettel, K.; Willinger, B. Identification of Filamentous Fungi by MALDI-TOF Mass Spectrometry: Evaluation of Three Different Sample Preparation Methods and Validation of an In-House Species Cutoff. J. Fungi 2022, 8, 383. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.H.; Mawla, H.A. Sedimentation with the Omeliansky formula as an accepted technique for quantifying airborne fungi. Pol. J. Environ. Stud. 2012, 21, 1539–1541. [Google Scholar]
- Amaga, I.; Takahashi, T.; Ishii, K.; Kato, M.; Kobayashi, Y. Antifungal effect of blue LED irradiation on the blue mold, Penicillium italicum, in Satsuma mandarin fruits. Hortic. Res. Jpn. 2015, 14, 83–87. [Google Scholar]
- Hussey, S.B.; Clark, R.; Lunn, K.F.; Breathnach, C.; Soboll, G.; Whalley, J.M.; Lunn, D.P. Detection and quantification of equine herpesvirus-1 viremia and nasal shedding by real-time polymerase chain reaction. J. Vet. Diagn. Investig. 2006, 18, 335–342. [Google Scholar] [CrossRef]
- Anees-Hill, S.; Douglas, P.; Pashley, C.; Hansell, A.; Marczylo, E. A systematic review of outdoor airborne fungal spore seasonality across Europe and the implications for health. Sci. Total Environ. 2021, 818, 151716. [Google Scholar] [CrossRef]
- Sautour, M.; Sixt, N.; Dalle, F.; L’Ollivier, C.; Fourquenet, V.; Calinon, C.; Paul, K.; Valvin, S.; Maurel, A.; Aho, S.; et al. Profiles and seasonal distribution of airborne fungi in indoor and outdoor environments at a French hospital. Sci. Total Environ. 2009, 407, 3766–3771. [Google Scholar] [CrossRef]
- Hao, Z.F.; Ao, J.H.; Hao, F.; Yang, R.Y.; Zhu, H.; Zhang, J. Environment surveillance of filamentous fungi in two tertiary care hospitals in China. Chin. Med. J. 2011, 124, 1970–1975. [Google Scholar] [PubMed]
- Priyamvada, H.; Singh, R.K.; Akila, M.; Ravikrishna, R.; Verma, R.S.; Gunthe, S.S. Seasonal variation of the dominant allergenic fungal aerosols—One year study from southern Indian region. Sci. Rep. 2017, 7, 11171. [Google Scholar] [CrossRef] [PubMed]
- Bensch, K.; Groenewald, J.Z.; Meijer, M.; Dijksterhuis, J.; Jurjević, Ž.; Andersen, B.; Houbraken, J.; Crous, P.W.; Samson, R.A. Cladosporium species in indoor environments. Stud. Mycol. 2018, 89, 177–301. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, B.; Hedayati, M.T.; Hedayati, N.; Ilkit, M.; Syedmousavi, S. Aspergillus species in indoor environments and their possible occupational and public health hazards. Curr. Med. Mycol. 2016, 2, 36–42. [Google Scholar] [CrossRef]
- Shamim, M.; Kumar, M.; Kumar, R.R.; Pandey, P.; Srivastava, D.; Kumar, D.; Khan, N.A.; Kumar, R.R.; Singh, K.N. Understanding the diversity of Aspergillus by next-generation sequencing. In New and Future Developments in Microbial Biotechnology and Bioengineering; Gupta, V.K., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; Chapter 2; pp. 29–40. [Google Scholar]
- Jiang, S.-Y.; Ma, A.; Ramachandran, S. Negative Air Ions and Their Effects on Human Health and Air Quality Improvement. Int. J. Mol. Sci. 2018, 19, 2966. [Google Scholar] [CrossRef]
- Secretariat, M.A. Air cleaning technologies: An evidence-based analysis. Ont. Health Technol. Assess. Ser. 2005, 5, 1–52. [Google Scholar]
- Weng, H.; Zhang, Y.; Huang, X.; Liu, X.; Tang, Y.; Yuan, H.; Xu, Y.; Li, K.; Zhang, Y. Pilot study on the production of negative oxygen ions based on lower voltage ionization method and application in air purification. Atmosphere 2024, 15, 860. [Google Scholar] [CrossRef]
- Čereška, A.; Tetsmann, I.; Bareikis, R.; Jasevičius, R. Research of Air Purification Using Ion Energy Effect on Particulate Matter Agglomeration. Atmosphere 2024, 15, 915. [Google Scholar] [CrossRef]
- Campler, M.R.; Shen, Y.-F.; Klüppel, L.M.; Arruda, A.G. Assessing the Impact of a Negative Air Ionization System on Particulate Matter and Gaseous Pollutants in the Swine Farrowing Environment. PLoS ONE 2025, 20, e0316914. [Google Scholar] [CrossRef]
- Slater, J. Equine Herpesviruses. In Equine Infectious Diseases; Sellon, D.C., Long, M.T., Eds.; WB Saunders: St. Louis, MO, USA, 2014; pp. 151–168. [Google Scholar]
- Pilawa, B.; Buszman, E.; Gondzik, A.; Wilczynski, S.; Zdybel, M.; Witoszynska, T.; Wilczok, T. Effect of pH on paramagnetic centers in Cladosporium cladosporioides melanin. Acta Phys. Pol. A 2005, 108, 147–150. [Google Scholar] [CrossRef]
- Llorente, C.; Bárcena, A.; Vera Bahima, J.; Saparrat, M.C.; Arambarri, A.M.; Rozas, M.F.; Mirífico, M.V.; Balatti, P.A. Cladosporium cladosporioides LPSC 1088 produces the 1,8-dihydroxynaphthalene-melanin-like compound and carries a putative pks gene. Mycopathologia 2012, 174, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.K.; Gajjar, D.U.; Vasavada, A.R. DOPA and DHN pathway orchestrate melanin synthesis in Aspergillus species. Med. Mycol. 2014, 52, 10–18. [Google Scholar]
- Hachinohe, M.; Matsumoto, H. Involvement of melanin synthesis and reactive oxygen species in phytotoxic action of L-DOPA in carrot cells. Crop Prot. 2007, 26, 294–298. [Google Scholar] [CrossRef]
- Shao, H.; Tu, Y.; Wang, Y.; Jiang, C.; Ma, L.; Hu, Z.; Wang, J.; Zeng, B.; He, B. Oxidative stress response of Aspergillus oryzae induced by hydrogen peroxide and menadione sodium bisulfite. Microorganisms 2019, 7, 225. [Google Scholar] [CrossRef]
- Cho, H.-J.; Son, S.-H.; Chen, W.; Son, Y.-E.; Lee, I.; Yu, J.-H.; Park, H.-S. Regulation of conidiogenesis in Aspergillus flavus. Cells 2022, 11, 2796. [Google Scholar] [CrossRef]
- Yu, J.-H. Regulation of development in Aspergillus nidulans and Aspergillus fumigatus. Mycobiology 2010, 38, 229–237. [Google Scholar] [CrossRef]
- Smith, T.D.; Calvo, A.M. The mtfA transcription factor gene controls morphogenesis, gliotoxin production, and virulence in the opportunistic human pathogen Aspergillus fumigatus. Eukaryot. Cell 2014, 13, 766–775. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Thermocycling Parameters | Primers and Probe (5′–3′) |
|---|---|
| 95 °C (60 s); 40 cycles at 95 °C (15 s) and 60 °C (30 s) | For: CATACGTCCCTGTCCGACAGAT Rev: GGTACTCGGCCTTTGACGAA Probe: 6FAM-TGAGACCGAAGATCTCCTCCACCGA-BHQ1 |
| Fungal Species | Hospital NTAS | Hospital ITAS | Residental NTAS | Residental ITAS |
|---|---|---|---|---|
| Aspergillus niger | 1 | 0 | 3 | 3 |
| Cladosporium herbarum | 9 | 9 | 2 | 1 |
| Cladosporium cladosporioides | 3 | 5 | 3 | 1 |
| Aspergillus versicolor | 1 | 3 | 0 | 0 |
| Epicoccum nigrum | 2 | 0 | 0 | 0 |
| Aspergillus calidoustus | 3 | 1 | 0 | 0 |
| Penicillium digitatum | 0 | 3 | 0 | 0 |
| Penicillium chrysogenum | 0 | 1 | 3 | 2 |
| Alternaria alternata | 0 | 1 | 2 | 0 |
| Rhizopus arrhizus (Rhizopus delemar) | 1 | 0 | 0 | 0 |
| Aspergillus flavus | 0 | 0 | 7 | 3 |
| Aspergillus pseudoglaucus | 0 | 0 | 1 | 0 |
| Trichothecium roseum | 0 | 0 | 1 | 0 |
| Didymella glomerata (Phoma glomerata) | 0 | 0 | 0 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mijatović, S.; Radalj, A.; Ilić, A.; Janković, M.; Trajković, J.; Djoković, S.; Gobeljić, B.; Sovtić, A.; Petrović, G.; Kuzmanović, M.; et al. Assessment of Suppressive Effects of Negative Air Ions on Fungal Growth, Sporulation and Airborne Viral Load. Atmosphere 2025, 16, 896. https://doi.org/10.3390/atmos16080896
Mijatović S, Radalj A, Ilić A, Janković M, Trajković J, Djoković S, Gobeljić B, Sovtić A, Petrović G, Kuzmanović M, et al. Assessment of Suppressive Effects of Negative Air Ions on Fungal Growth, Sporulation and Airborne Viral Load. Atmosphere. 2025; 16(8):896. https://doi.org/10.3390/atmos16080896
Chicago/Turabian StyleMijatović, Stefan, Andrea Radalj, Andjelija Ilić, Marko Janković, Jelena Trajković, Stefan Djoković, Borko Gobeljić, Aleksandar Sovtić, Gordana Petrović, Miloš Kuzmanović, and et al. 2025. "Assessment of Suppressive Effects of Negative Air Ions on Fungal Growth, Sporulation and Airborne Viral Load" Atmosphere 16, no. 8: 896. https://doi.org/10.3390/atmos16080896
APA StyleMijatović, S., Radalj, A., Ilić, A., Janković, M., Trajković, J., Djoković, S., Gobeljić, B., Sovtić, A., Petrović, G., Kuzmanović, M., Stanković, J. A., Kolarž, P., & Arandjelović, I. (2025). Assessment of Suppressive Effects of Negative Air Ions on Fungal Growth, Sporulation and Airborne Viral Load. Atmosphere, 16(8), 896. https://doi.org/10.3390/atmos16080896