
Impact of El Niño–Southern Oscillation on Global Vegetation

Abstract
1. Introduction
2. Impact of ENSO on Vegetation
2.1. El Niño and La Niña
2.1.1. Asia
2.1.2. The Maritime Continent
2.1.3. Australia
2.1.4. North America
2.1.5. The Amazon Region of South America
2.1.6. Africa
2.1.7. Global Vegetation
2.2. CP ENSO and EP ENSO
2.3. ENSO Interactions with Other Climate Modes
3. Discussion
3.1. Current State of Research
3.1.1. Limitations of Existing Studies
3.1.2. Uncertainty in the Impact of ENSO on Vegetation
3.1.3. Impacts of ENSO Diversity on Vegetation
3.2. Future Research Focus
3.2.1. Numerical Models Used to Investigate ENSO’s Influence on Vegetation Under Global Warming
3.2.2. Impacts of Multiyear La Niña Events on Vegetation
3.2.3. The Influence of ENSO on Other Aspects of Vegetation
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McPhaden, M.J.; Zebiak, S.E.; Glantz, M.H. ENSO as an integrating concept in earth science. Science 2006, 314, 1740–1745. [Google Scholar] [CrossRef] [PubMed]
- NOAA Climate Prediction Center (CPC). Cold & Warm Episodes by Season. 2025. Available online: https://www.cma.gov.cn/2011xwzx/2011xtt/201706/t20170628_429061.html (accessed on 27 March 2025).
- Trenberth, K.E. The definition of EI Niño. Bull. Am. Meteorol. Soc. 1997, 78, 2771–2778. [Google Scholar] [CrossRef]
- Taschetto, A.S.; Ummenhofer, C.C.; Stuecker, M.F.; Dommenget, D.; Ashok, K.; Rodrigues, R.R.; Yeh, S.W. ENSO atmospheric teleconnections. In El Niño Southern Oscillation in a Changing Climate; McPhaden, M.J., Santoso, A.W.C., Eds.; Wiley & Sons, Inc.: Hoboken, NJ, USA, 2020; pp. 309–335. [Google Scholar] [CrossRef]
- Wang, C. Comparison of Vegetation Anomalies to EI Niño-Southern Oscillation in China and Australia. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2022, 43, 1027–1032. [Google Scholar] [CrossRef]
- Lenssen, N.J.L.; Goddard, L.; Mason, S. Seasonal Forecast Skill of ENSO Teleconnection Maps. Weather Forecast. 2020, 35, 2387–2406. [Google Scholar] [CrossRef]
- Jiao, K.; Gao, J.; Wu, S.; Hou, W. Research progress on the response processes of vegetation activity to climate change. Acta Ecol. Sin. 2018, 38, 2229–2238. [Google Scholar] [CrossRef]
- IPPC Secretariat. Scientific Review of the Impact of Climate Change on Plant Pests-A Global Challenge to Prevent and Mitigate Plant Pest Risks in Agriculture, Forestry and Ecosystems; FAO on behalf of the IPPC Secretariat: Rome, Italy, 2021. [Google Scholar] [CrossRef]
- Jia, G.; Shevliakova, E.; Artaxo, P.; De Noblet-Ducoudré, N.; Houghton, R.; House, J.; Kitajima, K.; Lennard, C.; Popp, A.; Sirin, A.; et al. Land-climate interactions. In Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Shukla, P.R., Skea, J., Calvo Buendia, E., Masson-Delmotte, V., Pörtner, H.-O., Roberts, D.C., Zhai, P., Slade, R., Connors, S., van Diemen, R., et al., Eds.; IPCC: Geneva, Switzerland, 2019. [Google Scholar] [CrossRef]
- Ding, Y.; Li, Z.; Peng, S. Global analysis of time-lag and-accumulation effects of climate on vegetation growth. Int. J. Appl. Earth Obs. Geoinf. 2020, 92, 102179. [Google Scholar] [CrossRef]
- Jian, D.; Niu, G.-Y.; Ma, Z.; Liu, H.; Guan, D.; Zhou, X.; Zhou, J. Limited driving of elevated CO2 on vegetation greening over global drylands. Environ. Res. Lett. 2023, 18, 104024. [Google Scholar] [CrossRef]
- Stuecker, M.F.; Jin, F.F.; Timmermann, A.; McGregor, S. Combination Mode Dynamics of the Anomalous Northwest Pacific Anticyclone. J. Clim. 2015, 28, 1093–1111. [Google Scholar] [CrossRef]
- Li, J.; Fan, K.; Xu, Z. Asymmetric response in Northeast Asia of summer NDVI to the preceding ENSO cycle. Clim. Dyn. 2016, 47, 2765–2783. [Google Scholar] [CrossRef]
- Huang, S.; Tang, L.; Hupy, J.P.; Wang, Y.; Shao, G. A commentary review on the use of normalized difference vegetation index (NDVI) in the era of popular remote sensing. J. For. Res. 2020, 32, 1–6. [Google Scholar] [CrossRef]
- Zhao, Q.; Ma, X.; Yao, W.; Liu, Y.; Yao, Y. Anomaly Variation of Vegetation and Its Influencing Factors in Mainland China During ENSO Period. IEEE Access 2019, 8, 721–734. [Google Scholar] [CrossRef]
- Wang, C.; Li, J.; Liu, Q.; Huete, A.; Li, L.; Dong, Y.; Zhao, J. Eastern-Pacific and Central-Pacific Types of ENSO Elicit Diverse Responses of Vegetation in the West Pacific Region. Geophys. Res. Lett. 2022, 49, e2021GL096666. [Google Scholar] [CrossRef]
- Wang, H.; Dai, Y.; Yang, S.; Li, T.; Luo, J.; Sun, B.; Duan, M.; Ma, J.; Yin, Z.; Huang, Y. Predicting climate anomalies: A real challenge. Atmos. Oceanic Sci. Lett. 2022, 15, 100115. [Google Scholar] [CrossRef]
- Fang, H.; Baret, F.; Plummer, S.; Schaepman-Strub, G. An Overview of Global Leaf Area Index (LAI): Methods, Products, Validation, and Applications. Rev. Geophys. 2019, 57, 739–799. [Google Scholar] [CrossRef]
- Supari; Tangang, F.; Salimun, E.; Aldrian, E.; Sopaheluwakan, A.; Juneng, L. ENSO modulation of seasonal rainfall and extremes in Indonesia. Clim. Dyn. 2018, 51, 2559–2580. [Google Scholar] [CrossRef]
- Nagai, S.; Ichii, K.; Morimoto, H. Interannual variations in vegetation activities and climate variability caused by ENSO in tropical rainforests. Int. J. Remote Sens. 2007, 28, 1285–1297. [Google Scholar] [CrossRef]
- Arjasakusuma, S.; Yamaguchi, Y.; Hirano, Y.; Zhou, X. ENSO-and Rainfall-Sensitive Vegetation Regions in Indonesia as Identified from Multi-Sensor Remote Sensing Data. ISPRS Int. J. Geo-Inf. 2018, 7, 103. [Google Scholar] [CrossRef]
- Erasmi, S.; Propastin, P.; Kappas, M. Spatial Patterns of NDVI Variation over Indonesia and Their Relationship to ENSO Warm Events during the Period 1982–2006. J. Clim. 2009, 22, 6612–6623. [Google Scholar] [CrossRef]
- Yan, Y.; Mao, K.; Shen, X.; Cao, M.; Xu, T.; Guo, Z.; Bao, Q. Evaluation of the influence of ENSO on tropical vegetation in long time series using a new indicator. Ecol. Indic. 2021, 129, 107872. [Google Scholar] [CrossRef]
- Ma, X.; Huete, A.; Cleverly, J.; Eamus, D.; Chevallier, F.; Joiner, J.; Poulter, B.; Zhang, Y.; Guanter, L.; Meyer, W.; et al. Drought rapidly diminishes the large net CO₂ uptake in 2011 over semi-arid Australia. Sci. Rep. 2016, 6, 37747. [Google Scholar] [CrossRef]
- Byrne, M.; Yeates, D.K.; Joseph, L.; Kearney, M.; Bowler, J.; Williams, M.A.; Cooper, S.; Donnellan, S.C.; Keogh, J.S.; Leys, R.; et al. Birth of a biome: Insights into the assembly and maintenance of the Australian arid zone biota. Mol. Ecol. 2008, 17, 4398–4417. [Google Scholar] [CrossRef] [PubMed]
- Poulter, B.; Frank, D.; Ciais, P.; Myneni, R.B.; Andela, N.; Bi, J.; Broquet, G.; Canadell, J.G.; Chevallier, F.; Liu, Y.Y.; et al. Contribution of semi-arid ecosystems to interannual variability of the global carbon cycle. Nature 2014, 509, 600–603. [Google Scholar] [CrossRef]
- Liu, Y.; de Jeu, R.A.M.; van Dijk, A.I.J.M.; Owe, M. TRMM-TMI satellite observed soil moisture and vegetation density (1998–2005) show strong connection with El Niño in eastern Australia. Geophys. Res. Lett. 2007, 34. [Google Scholar] [CrossRef]
- Potter, C.; Boriah, S.; Steinbach, M.; Kumar, V.; Klooster, S. Terrestrial vegetation dynamics and global climate controls in North America: 2001–05. Earth Interact. 2008, 12, 1–12. [Google Scholar] [CrossRef]
- Li, Z.; Kafatos, M. Interannual Variability of Vegetation in the United States and Its Relation to El Niño/Southern Oscillation. Remote Sens. Environ. 2000, 71, 239–247. [Google Scholar] [CrossRef]
- Peters, A.J.; Ji, L.; Walter-Shea, E.; Southeastern, U.S. Vegetation Response to ENSO Events (1989–1999). Clim. Change 2003, 60, 175–188. [Google Scholar] [CrossRef]
- Mennis, J. Exploring relationships between ENSO and vegetation vigour in the south-east USA using AVHRR data. Int. J. Remote Sens. 2010, 22, 3077–3092. [Google Scholar] [CrossRef]
- Cai, W.; McPhaden, M.J.; Grimm, A.M.; Rodrigues, R.R.; Taschetto, A.S.; Garreaud, R.D.; Dewitte, B.; Poveda, G.; Ham, Y.-G.; Santoso, A.; et al. Climate impacts of the El Niño-Southern Oscillation on South America. Nat. Rev. Earth Environ. 2020, 1, 215–231. [Google Scholar] [CrossRef]
- Liu, W.; Juárez Negrón, R. ENSO drought onset prediction in northeast Brazil using NDVI. Int. J. Remote Sens. 2001, 22, 3483–3501. [Google Scholar] [CrossRef]
- Vilanova, R.S.; Delgado, R.C.; de Andrade, C.F.; dos Santos, G.L.; Magistrali, I.C.; de Oliveira, C.M.M.; Teodoro, P.E.; Silva, G.F.C.; da Silva Junior, C.A.; de Ávila Rodrigues, R. Vegetation degradation in ENSO events: Drought assessment, soil use and vegetation evapotranspiration in the Western Brazilian Amazon. Remote Sens. Appl. Soc. Environ. 2021, 23, 100531. [Google Scholar] [CrossRef]
- Malhi, Y.; Roberts, J.T.; Betts, R.A.; Killeen, T.J.; Li, W.; Nobre, C.A. Climate change, deforestation, and the fate of the Amazon. Science 2008, 319, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Hilker, T.; Lyapustin, A.I.; Tucker, C.J.; Hall, F.G.; Myneni, R.B.; Wang, Y.; Bi, J.; Mendes de Moura, Y.; Sellers, P.J. Vegetation dynamics and rainfall sensitivity of the Amazon. Proc. Natl. Acad. Sci. USA 2014, 111, 16041–16046. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, J.; Solera, A.; Avilés, A.; Tonón, M.D. Influence of ENSO on droughts and vegetation in a high mountain equatorial climate basin. Atmosphere 2022, 13, 2123. [Google Scholar] [CrossRef]
- Oliveira, L.; França, G.; Nicácio, R.; Antunes, M.; Costa, T.; Torres, A., Jr.; França, J. A study of the El Niño-Southern Oscillation influence on vegetation indices in Brazil using time series analysis from 1995 to 1999. Int. J. Remote Sens. 2010, 31, 423–437. [Google Scholar] [CrossRef]
- Pereira, M.P.S.; Mendes, K.R.; Justino, F.; Couto, F.; da Silva, A.S.; da Silva, D.F.; Malhado, A.C.M. Brazilian dry forest (Caatinga) response to multiple ENSO: The role of Atlantic and Pacific Ocean. Sci. Total Environ. 2020, 705, 135717. [Google Scholar] [CrossRef]
- Estupinan-Suarez, L.M.; Mahecha, M.D.; Brenning, A.; Kraemer, G.; Poveda, G.; Reichstein, M.; Sierra, C.A. Spatial patterns of vegetation activity related to ENSO in northern South America. J. Geophys. Res. Biogeosci. 2024, 129, e2022JG007344. [Google Scholar] [CrossRef]
- Vining, B.R.; Hillman, A.; Contreras, D. El Niño Southern Oscillation and enhanced arid land vegetation productivity in NW South America. J. Arid Environ. 2022, 198, 104695. [Google Scholar] [CrossRef]
- Hao, Z.; Hao, F.; Singh, V.P.; Zhang, X. Quantifying the relationship between compound dry and hot events and El Niño–southern Oscillation (ENSO) at the global scale. J. Hydrol. 2018, 567, 332–338. [Google Scholar] [CrossRef]
- Anyamba, A.; Tucker, C.J.; Eastman, J.R. NDVI anomaly patterns over Africa during the 1997/98 ENSO warm event. Int. J. Remote Sens. 2010, 22, 1847–1859. [Google Scholar] [CrossRef]
- Hao, Y.; Hao, Z.; Feng, S.; Zhang, X.; Hao, F. Response of vegetation to El Niño-Southern Oscillation (ENSO) via compound dry and hot events in southern Africa. Global Planet. Change 2020, 195, 103358. [Google Scholar] [CrossRef]
- Sazib, N.; Mladenova, L.E.; Bolten, J.D. Assessing the impact of ENSO on agriculture over Africa using earth observation data. Front. Sustain. Food Syst. 2020, 4, 509914. [Google Scholar] [CrossRef]
- Shikwambana, L.; Xongo, K.; Mashalane, M.; Mhangara, P. Climatic and Vegetation Response Patterns over South Africa during the 2010/2011 and 2015/2016 Strong ENSO Phases. Atmosphere 2023, 14, 416. [Google Scholar] [CrossRef]
- Gong, D.Y.; Ho, C.H. Detection of large-scale climate signals in spring vegetation index (normalized difference vegetation index) over the Northern Hemisphere. J. Geophys. Res. Atmos. 2003, 108. [Google Scholar] [CrossRef]
- Zhang, A.; Jia, G.; Epstein, H.E.; Xia, J. ENSO elicits opposing responses of semi-arid vegetation between Hemispheres. Sci. Rep. 2017, 7, 42281. [Google Scholar] [CrossRef] [PubMed]
- An, S.I.; Tziperman, E.; Okumura, Y.M.; Li, T. ENSO irregularity and asymmetry. In El Niño Southern Oscillation in a Changing Climate; McPhaden, M.J., Santoso, A., Cai, W., Eds.; Wiley & Sons, Inc.: Hoboken, NJ, USA, 2020; pp. 153–172. [Google Scholar] [CrossRef]
- Hu, Z.Z.; Kumar, A.; Huang, B.; Zhu, J.; L’Heureux, M.; McPhaden, M.J.; Yu, J.Y. The Interdecadal Shift of ENSO Properties in 1999/2000: A Review. J. Clim. 2020, 33, 4441–4462. [Google Scholar] [CrossRef]
- McPhaden, M.J. A 21st century shift in the relationship between ENSO SST and warm water volume anomalies. Geophys. Res. Lett. 2012, 39. [Google Scholar] [CrossRef]
- Xia, Y.; Sun, X.; Yan, Y.; Feng, W.; Huang, F.; Yang, X. Change of ENSO characteristics in response to global warming. Chin. Sci. Bull. 2017, 62, 1738–1751. [Google Scholar] [CrossRef]
- Kao, H.-Y.; Yu, J.-Y. Contrasting Eastern-Pacific and Central-Pacific Types of ENSO. J. Clim. 2009, 22, 615–632. [Google Scholar] [CrossRef]
- Lee, T.; McPhaden, M.J. Increasing intensity of El Niño in the central-equatorial Pacific. Geophys. Res. Lett. 2010, 37. [Google Scholar] [CrossRef]
- Yeh, S.W.; Cai, W.; Min, S.K.; McPhaden, M.J.; Dommenget, D.; Dewitte, B.; Collins, M.; Ashok, K.; An, S.I.; Yim, B.Y.; et al. ENSO Atmospheric Teleconnections and Their Response to Greenhouse Gas Forcing. Rev. Geophys. 2018, 56, 185–206. [Google Scholar] [CrossRef]
- Zhang, W.; Jin, F.-F.; Turner, A. Increasing autumn drought over southern China associated with ENSO regime shift. J. Geophys. Res. 2014, 41, 4020–4026. [Google Scholar] [CrossRef]
- Li, J.; Fan, K.; Zhou, L. Satellite Observations of El Niño Impacts on Eurasian Spring Vegetation Greenness during the Period 1982–2015. Remote Sens. 2017, 9, 628. [Google Scholar] [CrossRef]
- Chen, H.; Qian, M.; Hua, W. Vegetation cover change in the Indo-China peninsula in spring and its relation to ENSO. Trans. Atmos. Sci. 2020, 43, 1065–1075. [Google Scholar] [CrossRef]
- Philippon, N.; Martiny, N.; Camberlin, P.; Hoffman, M.; Gond, V. Timing and patterns of the ENSO signal in Africa over the last 30 years: Insights from normalized difference vegetation index data. J. Clim. 2014, 27, 2509–2532. [Google Scholar] [CrossRef]
- Li, J.; Zheng, F.; Sun, C.; Feng, J.; Wang, J. Pathways of Influence of the Northern Hemisphere Mid-high Latitudes on East Asian Climate: A Review. Adv. Atmos. Sci. 2019, 36, 902–921. [Google Scholar] [CrossRef]
- Cai, W.; Wu, L.; Lengaigne, M.; Li, T.; McGregor, S.; Kug, J.-S.; Yu, J.-Y.; Stuecker, M.F.; Santoso, A.; Li, X.; et al. Pantropical climate interactions. Science 2019, 363, eaav4236. [Google Scholar] [CrossRef]
- Saji, N.H.; Goswami, B.N.; Vinayachandran, P.N.; Yamagata, T. A dipole mode in the tropical Indian Ocean. Nature 1999, 401, 360–363. [Google Scholar] [CrossRef]
- Londhe, D.S.; Katpatal, Y.B.; Singh, C.K.; Bokde, N.D. Assessment of vegetation variation and its response to ENSO and IOD in the semi-arid ecosystem of Western India. J. Water Clim. Change 2023, 14, 2642–2662. [Google Scholar] [CrossRef]
- Cowan, T.; van Rensch, P.; Cai, W.; Hendon, H.H. An Asymmetry in the IOD and ENSO Teleconnection Pathway and Its Impact on Australian Climate. J. Clim. 2012, 25, 6318–6329. [Google Scholar] [CrossRef]
- Teng, Y.; Zhang, W.; Liu, C.; Geng, X.; Qin, C. Possible role of ENSO spatial pattern variation in interdecadal weakening of ENSO-IOD relationship. Acta Meteorol. Sin. 2020, 78, 210–220. [Google Scholar] [CrossRef]
- Williams, C.A.; Hanan, N.P. ENSO and IOD teleconnections for African ecosystems: Evidence of destructive interference between climate oscillations. Biogeosciences 2011, 8, 27–40. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, Z.; Stuecker, M.F.; Turner, A.G.; Jin, F.-F.; Geng, X. Impact of ENSO longitudinal position on teleconnections to the NAO. Clim. Dyn. 2019, 52, 257–274. [Google Scholar] [CrossRef]
- Liu, C.; Li, J.; Liu, Q.; Gao, J.; Mumtaz, F.; Dong, Y.; Wang, C.; Gu, C.; Zhao, J. Combined influence of ENSO and North Atlantic Oscillation (NAO) on Eurasian Steppe during 1982–2018. Sci. Total Environ. 2023, 892, 164735. [Google Scholar] [CrossRef]
- Wang, L.; Gong, H.; Lan, X. Interdecadal variation of the Arctic Oscillation and its influence on climate. Trans. Atmos. Sci. 2021, 44, 50–60. [Google Scholar] [CrossRef]
- Le, T. Increased impact of the El Niño–Southern Oscillation on global vegetation under future warming environment. Sci. Rep. 2023, 13, 14459. [Google Scholar] [CrossRef]
- Fer, I.; Tietjen, B.; Jeltsch, F.; Wolff, C. The influence of El Niño–Southern Oscillation regimes on eastern African vegetation and its future implications under the RCP8.5 warming scenario. Biogeosciences 2017, 14, 4355–4374. [Google Scholar] [CrossRef]
- Cai, W.; Borlace, S.; Lengaigne, M.; Rensch, P.v.; Collins, M.; Vecchi, G.; Timmermann, A.; Santoso, A.; McPhaden, M.J.; Wu, L.; et al. Increasing frequency of extreme El Niño events due to greenhouse warming. Nat. Clim. Change 2014, 4, 111–116. [Google Scholar] [CrossRef]
- Cai, W.; Wang, G.; Santoso, A.; McPhaden, M.J.; Wu, L.; Jin, F.-F.; Timmermann, A.; Collins, M.; Vecchi, G.; Lengaigne, M.; et al. Increased frequency of extreme La Niña events under greenhouse warming. Nat. Clim. Change 2015, 5, 132–137. [Google Scholar] [CrossRef]
- Cai, W.; Ng, B.; Geng, T.; Jia, F.; Wu, L.; Wang, G.; Liu, Y.; Gan, B.; Yang, K.; Santoso, A. Anthropogenic impacts on twentieth-century ENSO variability changes. Nat. Rev. Earth Environ. 2023, 4, 407–418. [Google Scholar] [CrossRef]
- Cai, W.; Santoso, A.; Collins, M.; Dewitte, B.; Karamperidou, C.; Kug, J.-S.; Lengaigne, M.; McPhaden, M.J.; Stuecker, M.F.; Taschetto, A.S. Changing El Niño–Southern oscillation in a warming climate. Nat. Rev. Earth Environ. 2021, 2, 628–644. [Google Scholar] [CrossRef]
- Cai, W.; Wang, G.; Dewitte, B.; Wu, L.; Santoso, A.; Takahashi, K.; Yang, Y.; Carréric, A.; McPhaden, M.J. Increased variability of eastern Pacific El Niño under greenhouse warming. Nature 2018, 564, 201–206. [Google Scholar] [CrossRef]
- Wang, G.; Cai, W.; Gan, B.; Wu, L.; Santoso, A.; Lin, X.; Chen, Z.; McPhaden, M.J. Continued increase of extreme El Niño frequency long after 1.5 C warming stabilization. Nat. Clim. Change 2017, 7, 568–572. [Google Scholar] [CrossRef]
- Zhu, Z.; Piao, S.; Myneni, R.B.; Huang, M.; Zeng, Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A.; et al. Greening of the Earth and its drivers. Nat. Clim. Change 2016, 6, 791–795. [Google Scholar] [CrossRef]
- Wang, B. ; Sun, W. ; Jin, C. ; Luo, X. ; Yang, Y.-M.; Li, T.; Xiang, B.; McPhaden, M.J.; Cane, M.A.; Jin, F.; et al. Understanding the recent increase in multiyear La Niñas. Nat. Clim. Change 2023, 13, 1075–1081. [Google Scholar] [CrossRef]
- Redmond, K.T.; Koch, R.W. Surface climate and streamflow variability in the western United States and their relationship to large-scale circulation indices. Water Resour. Res. 1991, 27, 2381–2399. [Google Scholar] [CrossRef]
- Dannenberg, M.P.; Song, C.; Hwang, T.; Wise, E.K. Empirical evidence of El Niño–Southern Oscillation influence on land surface phenology and productivity in the western United States. Remote Sens. Environ. 2015, 159, 167–180. [Google Scholar] [CrossRef]
- Dech, J.; Nosko, P. Rapid growth and early flowering in an invasive plant, purple loosestrife (Lythrum salicaria L.) during an El Nino spring. Int. J. Biometeorol. 2004, 49, 26–31. [Google Scholar] [CrossRef]
- Bretfeld, M.; Ewers, B.E.; Hall, J.S. Plant water use responses along secondary forest succession during the 2015–2016 El Niño drought in Panama. New Phytol. 2018, 219, 885–899. [Google Scholar] [CrossRef]
- Erasmi, S.; Schucknecht, A.; Barbosa, M.; Matschullat, J. Vegetation Greenness in Northeastern Brazil and Its Relation to ENSO Warm Events. Remote Sens. 2014, 6, 3041–3058. [Google Scholar] [CrossRef]
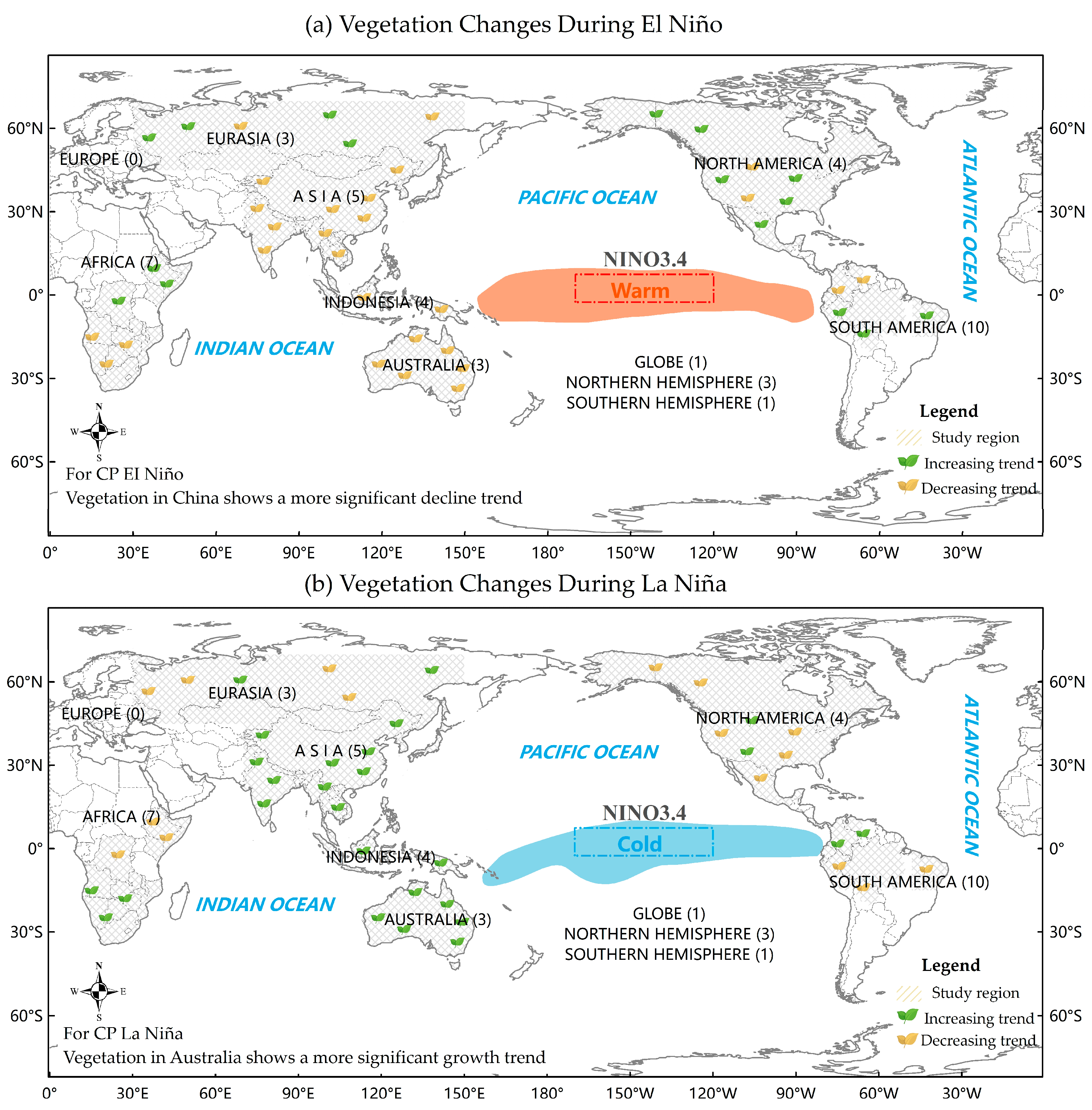


{kind=link}
{kind=link}
| Region | ENSO | ENSO | |
|---|---|---|---|
| El Niño | La Niña | ||
| Eurasia | The asymmetric response of atmospheric circulation to ENSO leads to asymmetric inverse phase changes in vegetation cover [13,57] | NDVI anomalies exhibit a negative–positive–negative pattern, reflecting the spatial variation in El Niño’s impacts Spring vegetation changes in the East Russian region are closely related to ENSO, with EP types inhibiting vegetation growth Significant negative NDVI anomalies occur in summer vegetation across Northeast Asia | Summer vegetation NDVI anomalies in Northeast Asia increased only slightly |
| China | ENSO disrupts the Asian monsoon, altering temperature and precipitation patterns, which in turn affect vegetation The distribution of ENSO sensitive vegetation in China exhibits notable regional variations [5,15] | Vegetation in the southern region (e.g., mixed forests) responds negatively to reduced precipitation Compared with EP El Niño, CP El Niño exerts a more pronounced inhibitory effect on vegetation | Positive response in central cultivated land, grassland in northern Inner Mongolia, and the northern basin of the Tibetan plateau |
| Central South Peninsula | ENSO influences the South Asian monsoon, and a strong correlation exits between vegetation and ENSO [58] | Inhibition of vegetation growth due to reduced precipitation | Favorable vegetation growth with increased cloudiness and precipitation |
| Maritime Continent | Situated at the core of the Walker Circulation, the region’s climate is strongly influenced by ENSO. Extreme rainfall events, closely related to ENSO, have a significant impact on local vegetation [21,22] | Significant decline in vegetation due to large-scale drought events, such as in 1982/1983 and 1997/1998 | Increased vegetation cover |
| Australia | The climate is influenced by various alterations in convective activity associated with ENSO La Niña events exert a more pronounced impact on vegetation, particularly in the northern and eastern regions Under comparable conditions, CP ENSO has a greater effect than EP ENSO | EP El Niño can either increase or decrease vegetation, while CP El Niño generally leads to vegetation decline | Compared with EP La Niña events, CP La Niña events significantly enhance vegetation growth |
| North America | ENSO negatively impacts on vegetation [28,29,30,31] | Promotes vegetation growth across most of North America, except in the eastern United States | Suppress vegetation growth |
| Amazon region | The combined influence of ENSO-driven modifications to the Walker Circulation and variations in the tropical Atlantic climate pattern plays a critical role in shaping regional climate dynamics ENSO alters precipitation patterns, with El Niño impacts greater than La Niña impacts [33,36,40,41] | Vegetation decreases in tropical regions but increases in extratropical zones Forest degradation in the Amazon region | Increased vegetation north of the equator and decreased vegetation south of the equator |
| Africa | Eastern and Southern vegetation is significantly influenced by precipitation pattern changes induced by ENSO, with a lagged response [43,45,46,59] | Enhanced vegetation cover in Eastern Africa (e.g., Ethiopian region) Decline in vegetation cover in Southern Africa | Crop yields increase in Southern Africa and decrease in Eastern Africa |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, J.; Jian, D.; Zhou, X.; Chen, Q.; Li, Y. Impact of El Niño–Southern Oscillation on Global Vegetation. Atmosphere 2025, 16, 701. https://doi.org/10.3390/atmos16060701
Jin J, Jian D, Zhou X, Chen Q, Li Y. Impact of El Niño–Southern Oscillation on Global Vegetation. Atmosphere. 2025; 16(6):701. https://doi.org/10.3390/atmos16060701
Chicago/Turabian StyleJin, Jie, Dongnan Jian, Xin Zhou, Quanliang Chen, and Yang Li. 2025. "Impact of El Niño–Southern Oscillation on Global Vegetation" Atmosphere 16, no. 6: 701. https://doi.org/10.3390/atmos16060701
APA StyleJin, J., Jian, D., Zhou, X., Chen, Q., & Li, Y. (2025). Impact of El Niño–Southern Oscillation on Global Vegetation. Atmosphere, 16(6), 701. https://doi.org/10.3390/atmos16060701