Abstract
Dust is the collection of fine particles of solid matter, and it is a major issue of atmospheric pollution. Dust particles are becoming the major pollutants of the urban environment due to hyperbolic manufacturing and automobile pollution. These atmospheric pollutants are not only hazardous for human beings, but they also affect tree growth, particularly in urban environments. This study was designed to examine the changes in morphological and physiological traits of three tree species seedlings (Eucalyptus camaldulensis, Conocarpus erectus, and Bombax ceiba) in response to different dust types. In a pot experiment under controlled conditions, three-month-old seedlings of selected trees species were subjected to four treatments of dust: T1 = controlled; T2 = wood dust; T3 = soil dust; and T4 = carbon dust. During the whole experiment, 10 g/plant/dose was applied in 8 doses with a one-week interval. The results depicted that the growth was the maximum in T1 (control) and the minimum in T4 (carbon dust). In our findings, B. ceiba performed better under the same levels of dust pollution as compared with the other two tree species. The B. ceiba tree species proved to be the most tolerant to dust pollution by efficiently demolishing oxidative bursts by triggering SOD, POD, and CAT under different dust types compared to controlled conditions. Stomatal conductance, photosynthetic rate, and transpiration rate were negatively influenced in all three tree species in response to different dust applications. Based on the findings, among these three tree species, B. ceiba is recommended for dust polluted areas followed by E. camaldulensis and Conocarpus erectus due to their better performance and efficient dust-foraging potential.
1. Introduction
Environmental factors can affect the eco-physiology of different tree species to ensure their survival in an exceedingly explicit atmosphere. Several environments impart stress that disturbs the equilibrium potential of plants [1]. In this condition, a given species can cope with these extreme environmental conditions through specific physiological mechanisms in the system which act as efficient airborne pollutants interceptors [2]. Dust particles are the main factors of atmospheric pollution taking place because of industrial processes and cause serious threats to living beings. In India, almost 35% of dirt particles are present in the air, which causes air pollution [3].
Dust consists of solid matter in a minute and fine state of subdivision so that the particles are small enough to be raised and carried by the wind. Different activities such as coal-based power plants, road transport, stone crushing, thermal power plants, and cement industries are adding an excessive amount of dust particles to the atmosphere [3]. Dust particles often arise from natural sources such as soil dust lifted up by the wind and volcanic eruptions, but occasionally, dust may also contain small amounts of pollen grains, human and animal hairs, and paper fibers [4]. In the current situation of wide-scale deforestation, the destruction of biota and other ecosystem elements is observed as an impact of air pollution [5].
The enhanced dust particle deposition on various parts of trees causes a serious threat. Leaves of delicate trees are extremely exposed to fine particulate matter in the atmosphere [6]. Moreover, several tree species facilitate getting rid of such impacting dust particles from the atmosphere to reduce their load and clean the air quality in urban places. Nowadays, dust is turning into a big challenge for the atmosphere, human health, plant physiology, and their ecological interactions [7]. In plants, gas exchange and water uptake potential are slowed down when dust forms a layer on the leaves of plants, which influences photosynthesis [8]. The absorbance level of infrared radiations is 2–3 °C higher in dusted leaves than dust-free ones, which has a degradation impact on tree growth [9].
Plants mostly depend on the leaves’ surface area, epidermal strength, height, and crown of the trees [10,11]. Atmospheric dust ends up causing iron deficiency in leaves because of its impact on the biogenesis of chlorophyll [12]. It is expected that the smaller size of dirt particles causes additional stress and more harm; however, these particles’ impact on plants has not gained much attention [3]. There may be visible injury symptoms in plants, resulting in biomass decline. The structure of the plant is altered by dust deposition, such as epiphytic lichen, and sphagnum communities are highly sensitive to dust [13,14]. Plant species, particularly trees and shrubs, are important sinks for trapping and absorbing many gases, particulates, aerosols, and airborne pollutants [15].
Plants, an important part of all ecosystems, play a crucial role in monitoring and maintaining the ecological balance by actively participating in the cycling of nutrients and gases such as carbon dioxide, oxygen, etc. [16]. They are most likely to be affected by airborne pollutants, and the effects are widely observed on the leaves, which are usually the most abundant and most obvious primary receptors of a large number of air pollutants [17]. Plants provide an enormous leaf area for the impingement, absorption, and accumulation of air pollutants to reduce the pollution level in the air environment [18].
E. camaldulensis, C. erectus, and B. ceiba are fast-growing, resilient, and incredibly efficient: these are just a few of the qualities that make bamboo such a good crop for both farmers and investors alike. These species are widely planted in urban environments under semiarid climates. They are multipurpose and have several environmental and economic benefits [12,19,20].
The roadside plants play a significant role in the assimilation and accumulation of pollutants and act as efficient interceptors of airborne pollutants. Dust impairs visibility, and the particulate dust falling on leaves may cause foliar injuries, reduction in yield, change in photosynthesis, and transpiration rate [21,22]. Different studies have been published on the effect of dust on physiological functions and leaf characteristics of forest or ornamental species and vegetables. However, there is still very limited work on the effect of various dust types on trees when planted along dusty roads. Because E. camaldulensis, C. erectus, and B. ceiba are our local species, their use in urban areas is economically beneficial nowadays. Consequently, the present study was meant to evaluate the effect of different types of dust application on morphological, physiological, and biochemical traits of Eucalyptus camaldulensis, Conocarpus erectus, and Bombax ceiba tree species.
2. Materials and Methods
2.1. Site Description
This proposed study was conducted in the Department of Forestry and Range Management, University of Agriculture, Faisalabad (UAF) research area. The location was situated at a latitude of 30.35° to 31.47° N and longitude of 72.08°–73° E, including an elevation of 184 m. Agricultural meteorological cell UAF observed weather conditions throughout the trial. The physical and chemical aspects of soil were also included (Table 1).

Table 1.
Physico-chemical Properties of soil.
2.2. Plant Materials
A pot experiment was conducted to check the effect of dust on the growth of Eucalyptus camaldulensis, Bombax ceiba, and Conocarpus erectus. Three-month-old seedlings of E. camaldulensis and C. erectus while stumps of the same height for B. ceiba were taken from Punjab Forestry Research Institute (PFRI) Gatwala, Faisalabad. Plants were grown in pots with 8 kg of loamy soil for one month to give them sufficient time for establishment in a new environment. Three different types of dust were collected. Wood dust was collected from the sawmill, soil dust from the earth’s surface, and carbon dust from bricks kiln. These different dust types range from 90 to 300 microns; however, carbon dust is found to be more refine. The following treatments, T1 = Controlled; T2 = Wood dust (300 microns); T3 = Soil dust (150 microns); and T4 = Carbon dust (90 microns), were used to check the impact of dust types on the growth of the tree species as mentioned above.
In this experiment, 80 g/plant of dust (10 g/plant/dose) was applied in 8 doses with a one-week interval on 27 plants, each treatment with 3 replications in 2 months (total plants = 108). A unique plastic bottle with a small hole in the lid cover covered with porous cloth was designed to apply the dust to plants.
2.3. Measurement of Plant Growth Parameters
Seedlings of all species were harvested, and initially, plant height was measured with tailor tape, and stem diameter was measured with a digital vernier caliper. Root and shoot fresh weights were measured, and then all the samples were put into the drying oven at 80 °C for 24 h to measure their dry weights.
2.4. Measurement of Physiological Parameters
Physiological parameters (stomatal conductance, photosynthetic rate, transpiration rate) were measured using an Infra-Red Gas Analyzer. IRGA was used in the morning when the sunlight was normal. Readings of second mature leaves were taken from each plant.
2.5. Measurement of Biochemical Parameters
For analyzing antioxidant enzymes (SOD, POD, and CAT), extraction was made as described by [23].
2.6. Measurement of Chlorophyll and Carotenoids Contents
Chlorophyll was extracted from 0.5 g leaf discs with 10 mL 80% acetone at 10 degrees centigrade overnight and then centrifuged at 14,000 rpm for 5 min. The absorbance of the supernatant was measured at 645, 652, 663, and 480 nm on a spectrophotometer (PG, T60U, Leicestershire, UK).
The chlorophyll and carotenoid contents were calculated with the following formula:
Chlorophyll a (mg g−1 FW) = [12.7 (OD663) − 2.69 (OD645)] × v/1000 × w
Chlorophyll b (mg g−1 FW) = [22.9 (OD645) − 4.68 (OD663)] × v/1000 × w
Chlorophyll a + b (mg g−1 FW) = [20.2 (OD645) − 8.02 (OD663)] × v/1000 × w
Carotenoids (mg g−1 FW) = Acar/Emax100
2.7. Statistical Analysis
The collected data were subjected to a two-factor factorial analysis of variance (ANOVA), while the treatment means were compared using the LSD test at 95% probability by using Minitab 19. GraphPad Prism 8.02 (GraphPad Software, La Jolla, San Diego, CA, USA) was used to make graphs.
3. Results
3.1. Morphological Traits
There were significant differences observed in treatment and species on plant height (p < 0.001). Results demonstrated that maximum plant height was obtained in B. ceiba (34.11 ± 2.1 cm), whereas the lowest plant height was observed in C. erectus (21.11 ± 1.1 cm) in response to T1 treatment. The T1 treatment found the largest plant height, while the T4 treatment observed the smallest plant height in all species (Figure 1a).
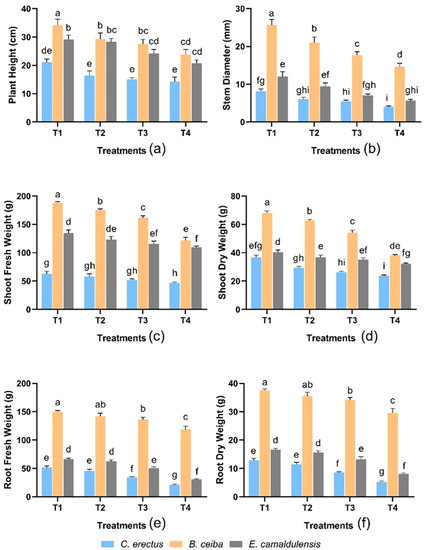
Figure 1.
Effect of different types of dust on (a) plant height (cm), (b) stem diameter (mm), (c) shoot fresh weight (g), (d) shoot dry weight (g), (e) root fresh weight (g), and (f) root dry weight (g). Notes: Different letters shows that values are significantly varied.
In terms of stem diameter, the interaction between treatment and species was significant (p < 0.01). B. ceiba showed a greater stem diameter (25.64 ± 1.4 mm), while C. erectus was observed to be lower (8.08 ± 0.6 mm) at the T1 treatment. The impact of treatments on the stem diameter in the given species was observed in the order: T1 > T2 > T3 > T4 (Figure 1b).
3.2. Biomass Distribution
Shoot (fresh and dry) weight was observed to be significantly different with the interaction of treatments and species (p < 0.001). The shoot biomass gradually decreased towards the T4 treatment, followed by all species. According to the findings, B. ceiba was higher shoot biomass, and C. erectus was found to have the lowest (Figure 1c,d).
In root (fresh and dry) weight, there was non-significant interaction between treatment and species were studied. The trend of root biomass in given species was found as B. ceiba > E. camaldulensis > C. erectus in response to all treatments (Figure 1e,f).
3.3. Physiological Traits
With regard to plant physiological traits, the interaction between treatments and species was found significantly varied (p < 0.001) for stomatal conductance. E. camaldulensis exhibited greater stomatal conductance when T1 treatment was added to the growing media, whereas a lower response was observed in C. erectus. Figure 2 showed that the photosynthetic rate observed similarly higher in all species in response to T1 treatment while gradually decreasing towards the T4 treatment. Consequently, the transpiration rate was maximum in E. camaldulensis, and the decline was found in C. erectus at the T1 treatment.
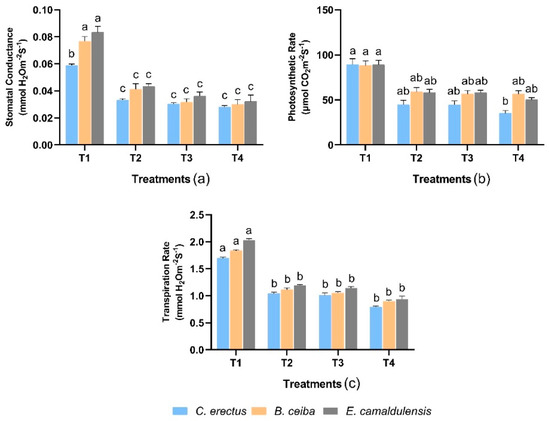
Figure 2.
Effect of different types of dust on (a) stomatal conductance (mmol H2O m−2 S−1), (b) photosynthetic rate (μmol CO2 m−2 S−1), and (c) transpiration rate (mmol H2O m−2 S−1). Notes: Different letters shows that values are significantly varied.
3.4. Enzyme Activities of Antioxidant Systems
Among the examined enzymes, the interaction between treatment and species was significant (p = 0.02) for SOD and non-significant for POD and CAT. The activities of antioxidant enzymes in both species are shown in (Figure 3). Results revealed that SOD and POD activity was higher in B. ceiba while the CAT activity was found higher in B. ceiba and E. camaldulensis. In all species, the enzymes’ response to the treatments was in the order: T4 > T3 >T2 > T1.
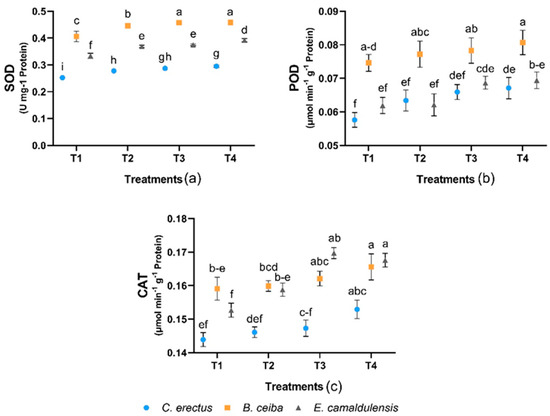
Figure 3.
Effect of different types of dust on (a) superoxide dismutase SOD (U mg−1 Protein), (b) peroxidase POD (μmol min−1 g−1 Protein), and (c) catalase CAT (μmol min−1 g−1 Protein). Notes: Different letters shows that values are significantly varied.
3.5. Chlorophyll Content
Chlorophyll content was observed to be non-significant with respect to within-subject factor interaction (treatment and species). According to the findings, T1 treatment had a greater influence on all species for chlorophyll content. Chlorophyll a, b, and a + b was observed in all species with the trend of B. ceiba > E. camaldulensis > C. erectus while carotenoids B. ceiba > E. camaldulensis = C. erectus (Figure 4). Chlorophyll content gradually declined towards the T4 treatment.
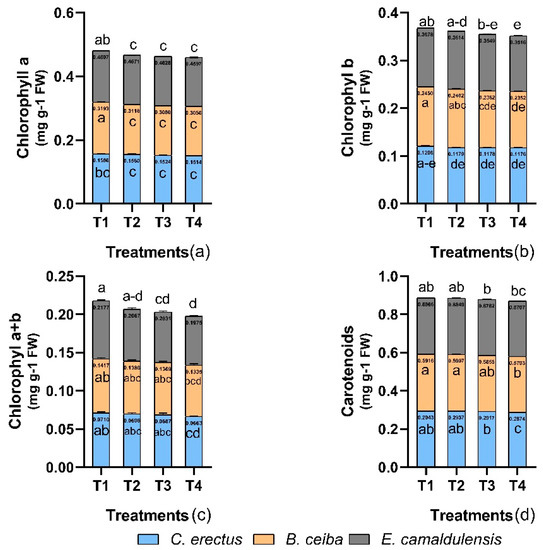
Figure 4.
Effect of different types of dust on (a) chlorophyl a (mg g−1 FW), (b) chlorophyl b (mg g−1 FW), (c) chlorophyl a + b (mg g−1 FW), and (d) carotenoids (mg g−1 FW). Notes: Different letters shows that values are significantly varied.
3.6. Principal Component Analysis of Plant Growth Parameters, Plant Physiology and Plant Biochemical Contents
To further evaluate the effect of different treatments of dust on seedling’s morphology, biomass distribution, physiology, and biochemical content, a multivariate analysis was executed. Figure 5 demonstrated that the effect of all dust treatments varied significantly among all seedling’s growth parameters. According to the analysis, all studied parameters positively contributed to the PC1 component. In PC2 components, plant height, stem diameter, shoot fresh weight (SFW), shoot dry weight (SDW), root fresh weight (RFW), root dry weight (RDW), SOD, POD, and CAT showed negative contributions. In contrast, stomatal conductance, photosynthetic rate, transpiration rate, chlorophyll a, b, a + b, and carotenoids showed a positive influence.
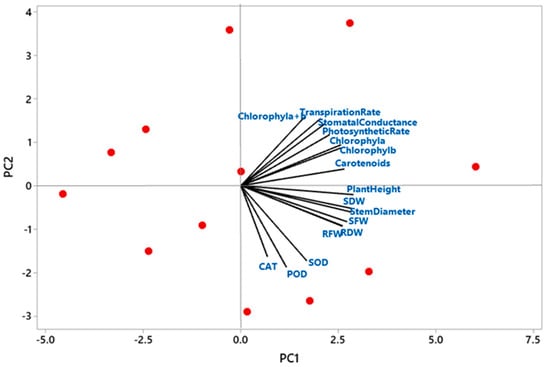
Figure 5.
Principal component analysis (PCA) of different studied parameters such as plant growth, plant physiology, and plant biochemical contents.
4. Discussion
Dust particles are absorbed through various parts of trees affecting tree growth by reducing the chemical process rate and causing cell death and chlorophyll degradation. Several studies have shown that dust pollution has a negative impact on plants as it reduces photosynthesis and causes leaf fall with tissue death [24]. The response of the plant to dust accumulation may vary according to different species, as dust deposition fluctuates with plant species due to leaf orientation, leaf surface geometry, leaf pubescence, height, and canopy of roadside plants [25].
Our findings demonstrated that the maximum reduction in plant height, stem diameter, shoot and root biomass, and chlorophyll content were observed due to the severe effect of carbon dust intensity [26,27,28,29]. Dust affected leaves of the vegetation due to roadside automobiles and coal power plant, etc., as discussed by [30]. Many countries of the world have observed elevated amounts of metals in urban street dust, which has been associated with excessive vehicle traffic flow. Street dust have been discovered to have significant quantities of metals, which have a deleterious impact on plant growth and development. Metals from road traffic can alter the soil composition and have an effect on plants; in particular, they help in the reduction of chlorophyll content [31,32].
Accumulation of dust particles on leaf surfaces during a dusty circumstance has been shown to alter leaf attributes, particularly surface reflectance in the visible and short-wave infrared radiation spectrums, which confines the quality and quantity of light that reaches chloroplasts for plant photosynthesis activities [32]. Additionally, the stomata are obstructed, and the gas exchange is disrupted by the dusty carbon coating covering the plant tissues [29,33]. Consequently, net photosynthesis and transpiration decreases, resulting in a reduction of growth and biomass [29,34,35]. Leghari et al. [32] also found a link between Vitis vinifera’s shorter height and lower (green mass), net primary output, and chlorophyll content when the plants were exposed to road dust. In the current study, B. ceiba revealed higher values in most of the growth and biomass traits, whereas C. erectus showed lower productivity. As stomata become blocked by carbon and other pollution, it indicates that these variables are critical to the plant’s ability to develop and produce biomass [27,29].
Smaller dust particles enter the leaf through stomatal openings, and the larger particles usually pile up on the stomatal pores, affecting gaseous exchange processes, which in turn affect photosynthesis, respiration, and transpiration that allow the penetration of phytotoxic gaseous pollutants and overall plant growth [36,37,38]. In our investigation, carbon dust concentration reduced photosynthetic rate, transpiration rate, and stomatal conductance despite increasing leaf temperature in all three species, which is in line with the results of [29,39,40,41]. However, due to the dust-loaded leaf substrate, increasing near-infrared light spectrum irradiance absorption might raise leaf temperature and photorespiration while decreasing net photosynthesis [42].
The significant production of reactive oxygen species such as hydrogen peroxide (H2O2), hydroxyl radicals (OH−1), and superoxide radicals is affected by different toxicities in tree species. Lipid peroxidation is also induced by the overproduction of MDA in the case of excessive toxicity [43]. In significant amounts, ROS and lipid peroxidation cause oxidative stress in cellular components, resulting in ion loss, DNA denaturation and strand damage, protein hydrolysis and conformational changes, lipid peroxidation, and membrane concussion, ultimately leading to apoptosis [44,45,46]. Trees have inherent defensive mechanisms against this oxidative burst in the form of antioxidative enzymes such as superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT).
SOD’s primary role is to limit the amount of Haber–Weiss reaction substrates such as O2 and hydrogen peroxide (H2O2) and to minimize the detrimental threat of OH radical generation, which is known to be capable of reacting and potentially cause irreparable damage to cellular membranes, DNA, and proteins [45,46]. In our investigation (Figure 3), the toxic level of dust (carbon dust) increased the amount of SOD. This exposed that the carbon dust has a serious impact on plants which may exert their harmful effects via the production of free radicals. It was shown that B. ceiba had the highest levels of SOD activity when exposed to severe (carbon dust) toxicity than either E. camaldulensis or C. erectus, indicating that B. ceiba tree species have a better antioxidant mechanism for detoxifying ROS-damaging effects (Figure 3). B. ceiba performed well under different toxicities, i.e., heavy metals, etc., by activating antioxidant enzymes and limiting the oxidative stress more effectively [47].
Oxidative stress can be relieved by CAT enzymes, which decompose H2O2 into water and oxygen, but CAT is less efficient than POD in detoxifying H2O2. The mitochondrial membrane’s antiapoptotic potential is enhanced by CAT enzymes, which also reduces oxidative stress and restores the mitochondrial arrangement, restoring both replication and healing capacity [48]. Due to this, the tree species only engaged the antioxidative mechanism by raising SOD and POD concentrations to oppose ROS, which was less oxidative stress from heavy metals [49]. Our findings indicated that, E. camaldulensis, B. ceiba, and C. erectus produced CAT antioxidant enzymes under the toxicities of different dust types in the soil. It was shown that the CAT activity was less under control, and therefore, it increased at the intense effect of carbon dust (Figure 3). Under the different types of dust toxicities, the maximum CAT contents were recorded in B. ceiba, and E. camaldulensis compared to C. erectus, emphasizing the higher handling capacity of B. ceiba and E. camaldulensis tree species.
Furthermore, POD enzymes performed an important part in regulating ROS in plants and trees. In our results, B. ceiba produced a higher amount of POD enzymes in response to the severe toxicity of carbon dust (Figure 3). It highlighted B. ceiba’s effective ROS-scavenging capacity when compared to other subject tree species under similar conditions of dust toxicity.
However, POD and SOD are antioxidant enzymes attached to the cell wall of higher tree plants. They are actively engaging in the production of ROS, which is needed for the development of multiple plant and tree species and has already been confirmed in the cell walls of young plants, root systems, germinated seeds, and leaves but has not been expressly identified in mentioned species peapods [43]. This highlighted the significance of ROS when synthesized in sufficient amounts; nevertheless, excessive ROS production produces oxidative stress in different plant/tree species, which is destroyed by POD, SOD, and CAT enzymes. Earlier investigations revealed that POD enzymes are found to be the primary H2O2-degrading enzyme after ascorbate peroxidases are exhausted. POD improved the reduction of H2O2 and hydroperoxides to alcohols or water molecules in order to remove harmful organic hydroperoxides from the atmosphere [50].
This study observed greater quantities of chlorophyll a, b, a + b, and carotenoids in B. ceiba. Carbon dust intensity resulted in a decrease in the amount of photosynthetic pigments present in all species studied [16,27,38]. Chloroplast degradation from coal-dust pollution may result in infected leaves having lower concentrations of chlorophyll [16]. In order for photosynthetic pigments to be synthesized, light and oxygen must be involved. Dust-deposited particles adhere to the surface of the leaf and make a coating layer. This covering appears to obstruct the stomata, reducing the amount of oxygen and light that may be absorbed. This has the effect of decreasing the energy dissipation of those antenna proteins that are associated with light foraging [51,52]. Furthermore, declines in chlorophyll pigments may be caused by pH variations within the leaf cells. It appears that the solubility of chemical compounds derived from dust particles in cell sap may result in chlorophyll degradation or inhibition of enzymes required for chlorophyll production [53].
According to the findings of this study, carbon dust has an adverse effect on plant growth. This could be because carbon dust contains a variety of harmful particles. E. camaldulensis and Conocarpus erectus were found to have the most altered phenological behavior, whereas B. ceiba was found to have the least. Our environment is clearly being degraded by carbon dust pollution, which is an active ecological element. As a result of resistance to carbon dust toxicity, B. ceiba should be planted around the urban territories. The study also recommends that a comprehensive examination of the harmful contaminants included in carbon dust be conducted. Ultimately, it is expected that such studies would allow for the recommendation of plants for use as green belts in industrial regions and bad urban sites to reduce dust and enhance air quality.
5. Conclusions
The findings of the study indicated that the morphology and physiology of B. ceiba, E. camaldulensis, and C. erectus could be dramatically altered by the presence of different dust types. A combination of oxidative burst caused by ROS overproduction and the resulting root cell osmotic stress caused by dust pollution dramatically increased the activity of antioxidant enzymes (e.g., CAT, SOD, and POD) while decreasing total biomass and photosynthetic pigments in all tree species examined. All three tree species were negatively affected by dust pollution, although B. ceiba was shown to be more resistant than E. camaldulensis and C. erectus to this toxicity. This was due to B. ceiba’s ability to efficiently eliminate oxidative stress by activating antioxidant enzymes, reduce osmotic stress by controlling osmotic, water, and pressure potential, and improve photosynthesis by increasing transpiration rate, evapotranspiration rate, and stomatal conductance. B. ceiba, accompanied by E. camaldulensis, could be suggested to grow in places contaminated with different dust types as an efficient dust toxicities regulator.
Author Contributions
M.F.N.; methodology: M.S.-U.-R. and M.A.S.; validation: M.H.U.R. and E.S.D.; formal analysis: M.H.U.R. and N.R.A.; investigation: S.G. and T.H.F.; data curation: M.A.S. and J.I.; writing—original draft preparation: M.H.U.R.; writing—review and editing: M.F.N., M.H.U.R., S.S.A. and N.R.A.; visualization: J.I., S.G., S.S.A. and M.S.-U.-R.; supervision: M.F.N., M.H.U.R., T.H.F. and E.S.D.; funding acquisition: S.S.A. All authors have read and agreed to the published version of the manuscript.
Funding
The current research supported by Taif University Researchers Supporting Project number (TURSP-2020/38), Taif University, Taif, Saudi Arabia.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data obtained in this study is owned by the donor and University. However, data can be made available on request.
Acknowledgments
The authors extend their appreciation to Taif University for funding current work by Taif University Researchers Supporting Project number (TURSP-2020/38), Taif University, Taif, Saudi Arabia. We are thankful to the field and laboratory staff of University of Agriculture Faisalabad for helping in data acquisition.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Craven, D.; Gulamhussein, S.; Berlyn, G. Physiological and anatomical responses of Acacia koa (Gray) seedlings to varying light and drought conditions. Environ. Exp. Bot. 2010, 69, 205–213. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Kramer, P.J.; Pallardy, S.G. The Physiological Ecology of Woody Plant; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Shukla, S.; Sharma, R.; Sahu, M. Research article dust pollution affect morphophysiological traits of plant Mangifera indica Linn. Int. J. Bot. 2019, 15, 1–4. [Google Scholar] [CrossRef]
- Hess-Kosa, K. Indoor Air Quality: Sampling Methodologies; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Sett, R. Responses in plants exposed to dust pollution. Hortic. Int. J. 2017, 1, 53–56. [Google Scholar] [CrossRef]
- Areington, C.A.; Varghese, B.; Ramdhani, S. An assessment of morphological, physiological and biochemical biomarkers of industrial air pollution in the leaves of Brachylaena discolor. Water Air Soil Pollut. 2015, 226, 291. [Google Scholar] [CrossRef]
- Singh, H.; Yadav, M.; Kumar, N.; Kumar, A.; Kumar, M. Assessing adaptation and mitigation potential of roadside trees under the influence of vehicular emissions: A case study of Grevillea robusta and Mangifera indica planted in an urban city of India. PLoS ONE 2020, 15, e0227380. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.; Amin, N.; Ahmad, I.; Shah, S.; Hussain, K. Dust particles induce stress, reduce various photosynthetic pigments and their derivatives in Ficus benjamina: A landscape plant. Int. J. Agric. Biol. 2017, 19, 1469–1474. [Google Scholar]
- Sharifi, M.R.; Gibson, A.C.; Rundel, P.W. Surface dust impacts on gas exchange in Mojave Desert shrubs. J. Appl. Ecol. 1997, 34, 837–846. [Google Scholar] [CrossRef]
- Farooq, T.H.; Yan, W.; Chen, X.; Shakoor, A.; Rashid, M.H.U.; Gilani, M.M.; He, Z.; Wu, P. Dynamics of canopy development of Cunninghamia lanceolata mid-age plantation in relation to foliar nitrogen and soil quality influenced by stand density. Glob. Ecol. Conserv. 2020, 24, e01209. [Google Scholar] [CrossRef]
- Singh, S.; Singh, P.; Mishra, R.; Singh, M. Leaf dust accumulation and its impact on chlorophyll content of Azadirachta indica and Bauhinia variegata developing in the proximity of jaypee cement plant, Rewa (Mp), India. Int. J. Biol. Innov. 2021, 3, 173–178. [Google Scholar] [CrossRef]
- Nawaz, M.F.; Rashid, M.H.U.; Arif, M.Z.; Sabir, M.A.; Farooq, T.H.; Gul, S.; Gautam, N.P. Ecophysiological response of Eucalyptus camaldulensis to dust and lead pollution. N. Z. J. For. Sci. 2021, 51, 13. [Google Scholar] [CrossRef]
- Gilbert, O. An alkaline dust effect on epiphytic lichens. Lichenologist 1976, 8, 173–178. [Google Scholar] [CrossRef]
- Walker, D.; Everett, K. Road dust and its environmental impact on Alaskan taiga and tundra. Arct. Alp. Res. 1987, 19, 479–489. [Google Scholar] [CrossRef]
- Gaghate, D.; Hasan, M. Ambient lead levels in urban areas. Bull. Environ. Contam. Toxicol. 1999, 62, 403–408. [Google Scholar] [CrossRef]
- Rai, P.K. Impacts of particulate matter pollution on plants: Implications for environmental biomonitoring. Ecotoxicol. Environ. Saf. 2016, 129, 120–136. [Google Scholar] [CrossRef]
- Sharma, B.; Sharma, S.; Bhardwaj, S. Plant-pollutant interactions with a special mention of dust accumulation by plants—A review. Nat. Environ. Pollut. Technol. 2017, 16, 375–384. [Google Scholar]
- Escobedo, F.J.; Wagner, J.E.; Nowak, D.J.; De la Maza, C.L.; Rodriguez, M.; Crane, D.E. Analyzing the cost effectiveness of Santiago, Chile’s policy of using urban forests to improve air quality. J. Environ. Manag. 2008, 86, 148–157. [Google Scholar] [CrossRef]
- Ahmad, I.; Jabeen, B.; Nawaz, M.F.; Asif, M.; Rashid, M.H.U.; Hussain, M.; Farooq, T.H.; Ahmed, S.; Rafiq, M. Influence of salinity on the morphological behavior and ionic response of different commercially important bamboo species. Fresenius Environ. Bull. 2022, 31, 668–676. [Google Scholar]
- Bashir, M.; Uzair, M.; Chaudhry, B.A. A review of phytochemical and biological studies on Conocarpus erectus (Combretaceae). Pak. J. Pharm. Res. 2015, 1, 1–8. [Google Scholar] [CrossRef]
- Singare, P.U.; Talpade, M.S. Physiological responses of some plant species as a bio-indicator of roadside automobile pollution stress using the air pollution tolerance index approach. Int. J. Plant Res. 2013, 3, 9–16. [Google Scholar]
- Raina, A.K.; Rathore, V.; Sharma, A. Effect of stone crusher dust on leaves of Melia azedarach Linn. and Dalbergia sissoo Roxb. in Jammu (J&K). Nat. Environ. Pollut. Technol. 2008, 7, 279. [Google Scholar]
- Alici, E.H.; Arabaci, G. Determination of SOD, POD, PPO and CAT enzyme activities in Rumex obtusifolius L. Annu. Res. Rev. Biol. 2016, 11, 1–7. [Google Scholar] [CrossRef]
- Bao, L.; Ma, K.; Zhang, S.; Lin, L.; Qu, L. Urban dust load impact on gas-exchange parameters and growth of Sophora japonica L. seedlings. Plant Soil Environ. 2015, 61, 309–315. [Google Scholar] [CrossRef]
- Chaturvedi, R.; Prasad, S.; Rana, S.; Obaidullah, S.; Pandey, V.; Singh, H. Effect of dust load on the leaf attributes of the tree species growing along the roadside. Environ. Monit. Assess. 2013, 185, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Tomar, D.; Khan, A.A.; Ahmad, G. Response of potato plants to foliar application of cement dust. Trop. Plant Res. 2018, 5, 41–45. [Google Scholar] [CrossRef]
- Javanmard, Z.; Tabari Kouchaksaraei, M.; Bahrami, H.; Hosseini, S.; Modarres Sanavi, S.; Struve, D.; Ammere, C. Soil dust effects on morphological, physiological and biochemical responses of four tree species of semiarid regions. Eur. J. For. Res. 2020, 139, 333–348. [Google Scholar] [CrossRef]
- Gupta, M.; Ghouse, A. Effects of coal-smoke pollutants from different sources on the growth, chlorophyll content, stem anatomy and cuticular traits of Euphorbia hirta L. Environ. Pollut. 1987, 47, 221–229. [Google Scholar] [CrossRef]
- Iqbal, M.; Mahmooduzzafar; Nighat, F.; Khan, P.R. Photosynthetic, metabolic and growth responses of Triumfetta rhomboidea to coal-smoke pollution at different stages of plant ontogeny. J. Plant Interact. 2010, 5, 11–19. [Google Scholar] [CrossRef]
- Swami, A. Impact of automobile induced air pollution on roadside vegetation: A review. Essence-Int. J. Environ. Rehabil. Conserv. 2018, 9, 101–116. [Google Scholar] [CrossRef]
- Al-Taani, A.A.; Nazzal, Y.; Howari, F.M. Assessment of heavy metals in roadside dust along the Abu Dhabi–Al Ain National Highway, UAE. Environ. Earth Sci. 2019, 78, 411. [Google Scholar] [CrossRef]
- Leghari, S.K.; Zaid, M.A.; Sarangzai, A.M.; Faheem, M.; Shawani, G.R. Effect of road side dust pollution on the growth and total chlorophyll contents in Vitis vinifera L.(grape). Afr. J. Biotechnol. 2014, 13, 1237–1242. [Google Scholar]
- Rai, P.K.; Singh, M.M. Lantana camara invasion in urban forests of an Indo–Burma hotspot region and its ecosustainable management implication through biomonitoring of particulate matter. J. Asia-Pac. Biodivers. 2015, 8, 375–381. [Google Scholar] [CrossRef]
- Shah, K.; Ahmad, I.; Ara, G. Impact assessment of leaf pigments in selected landscape plants exposed to roadside dust. Environ. Sci. Pollut. Res. 2018, 25, 23055–23073. [Google Scholar] [CrossRef]
- Pavlík, M.; Pavlíková, D.; Zemanová, V.; Hnilička, F.; Urbanová, V.; Száková, J. Trace elements present in airborne particulate matter—Stressors of plant metabolism. Ecotoxicol. Environ. Saf. 2012, 79, 101–107. [Google Scholar] [CrossRef]
- Rai, A.; Kulshreshtha, K.; Srivastava, P.; Mohanty, C. Leaf surface structure alterations due to particulate pollution in some common plants. Environmentalist 2010, 30, 18–23. [Google Scholar] [CrossRef]
- Zhu, J.; Zhang, X.; He, W.; Yan, X.; Yu, Q.; Xu, C.; Jiang, Q.O.; Huang, H.; Wang, R. Response of plant reflectance spectrum to simulated dust deposition and its estimation model. Sci. Rep. 2020, 10, 15803. [Google Scholar] [CrossRef]
- Siqueira-Silva, A.I.; Pereira, E.G.; Modolo, L.V.; Lemos-Filho, J.P.; Paiva, E.A.S. Impact of cement dust pollution on Cedrela fissilis Vell. (Meliaceae): A potential bioindicator species. Chemosphere 2016, 158, 56–65. [Google Scholar] [CrossRef]
- Zhan-Yi, W.; Jia, H.; Jian-Ying, G.; Cheng-Jie, W.; Ming-Jiu, W. Coal dust reduce the rate of root growth and photosynthesis of five plant species in inner Mongolian grassland. J. Residuals Sci. Technol. 2016, 13, S63–S73. [Google Scholar] [CrossRef][Green Version]
- Moradi, A.; Taheri Abkenar, K.; Afshar Mohammadian, M.; Shabanian, N. Effects of dust on forest tree health in Zagros oak forests. Environ. Monit. Assess. 2017, 189, 549. [Google Scholar] [CrossRef]
- Singh, K.; Chaturvedi, S.N.; Chauhan, S. Plant Biodiversity, Microbial Interaction and Environmental Biology; Pointer Publishers: Jaipur, India, 2005. [Google Scholar]
- Hirano, T.; Kiyota, M.; Aiga, I. Physical effects of dust on leaf physiology of cucumber and kidney bean plants. Environ. Pollut. 1995, 89, 255–261. [Google Scholar] [CrossRef]
- Su, L.-J.; Zhang, J.-H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.Y. Reactive oxygen species-induced lipid peroxidation in apoptosis, autophagy, and ferroptosis. Oxidative Med. Cell. Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef] [PubMed]
- Hendijani, A.K. Regulation of Pro-Longevity ROS by ROS-Handling Enzymes in C. elegans; McGill University: Montreal, QC, Canada, 2020. [Google Scholar]
- Sakthi, V.; Andal, N.M.; Rengaraj, S.; Sillanpää, M. Removal of Pb (II) ions from aqueous solutions using Bombax ceiba saw dust activated carbon. Desalin. Water Treat. 2010, 16, 262–270. [Google Scholar] [CrossRef]
- Guo, Y.; Yu, H.; Yang, M.; Kong, D.; Zhang, Y. Effect of drought stress on lipid peroxidation, osmotic adjustment and antioxidant enzyme activity of leaves and roots of Lycium ruthenicum Murr. seedling. J. Plant Physiol. 2018, 65, 244–250. [Google Scholar] [CrossRef]
- Siedlecka, A.; Krupa, Z. Functions of enzymes in heavy metal treated plants. In Physiology and Biochemistry of Metal Toxicity and Tolerance in Plants; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2002; pp. 303–324. [Google Scholar]
- Bela, K.; Horváth, E.; Gallé, Á.; Szabados, L.; Tari, I.; Csiszár, J. Plant glutathione peroxidases: Emerging role of the antioxidant enzymes in plant development and stress responses. J. Plant Physiol. 2015, 176, 192–201. [Google Scholar] [CrossRef]
- Horton, P.; Ruban, A. Molecular design of the photosystem II light-harvesting antenna: Photosynthesis and photoprotection. J. Exp. Bot. 2005, 56, 365–373. [Google Scholar] [CrossRef]
- Yuan, M.; Zhao, Y.-Q.; Zhang, Z.-W.; Chen, Y.-E.; Ding, C.-B.; Yuan, S. Light regulates transcription of chlorophyll biosynthetic genes during chloroplast biogenesis. Crit. Rev. Plant Sci. 2017, 36, 35–54. [Google Scholar] [CrossRef]
- Prusty, B.; Mishra, P.; Azeez, P. Dust accumulation and leaf pigment content in vegetation near the national highway at Sambalpur, Orissa, India. Ecotoxicol. Environ. Saf. 2005, 60, 228–235. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).