
The Response Mechanism and Threshold of Spring Wheat to Rapid Drought

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment Materials and Site
2.2. Experimental Design
2.3. Observation Content and Methods
2.4. Data Analysis
3. Results
3.1. Response Characteristics of Spring Wheat Leaves to Rapid-Drought Stress
3.1.1. Response Characteristics of Photosynthetic Physiological Parameters of Spring Wheat Leaves to Drought Stress
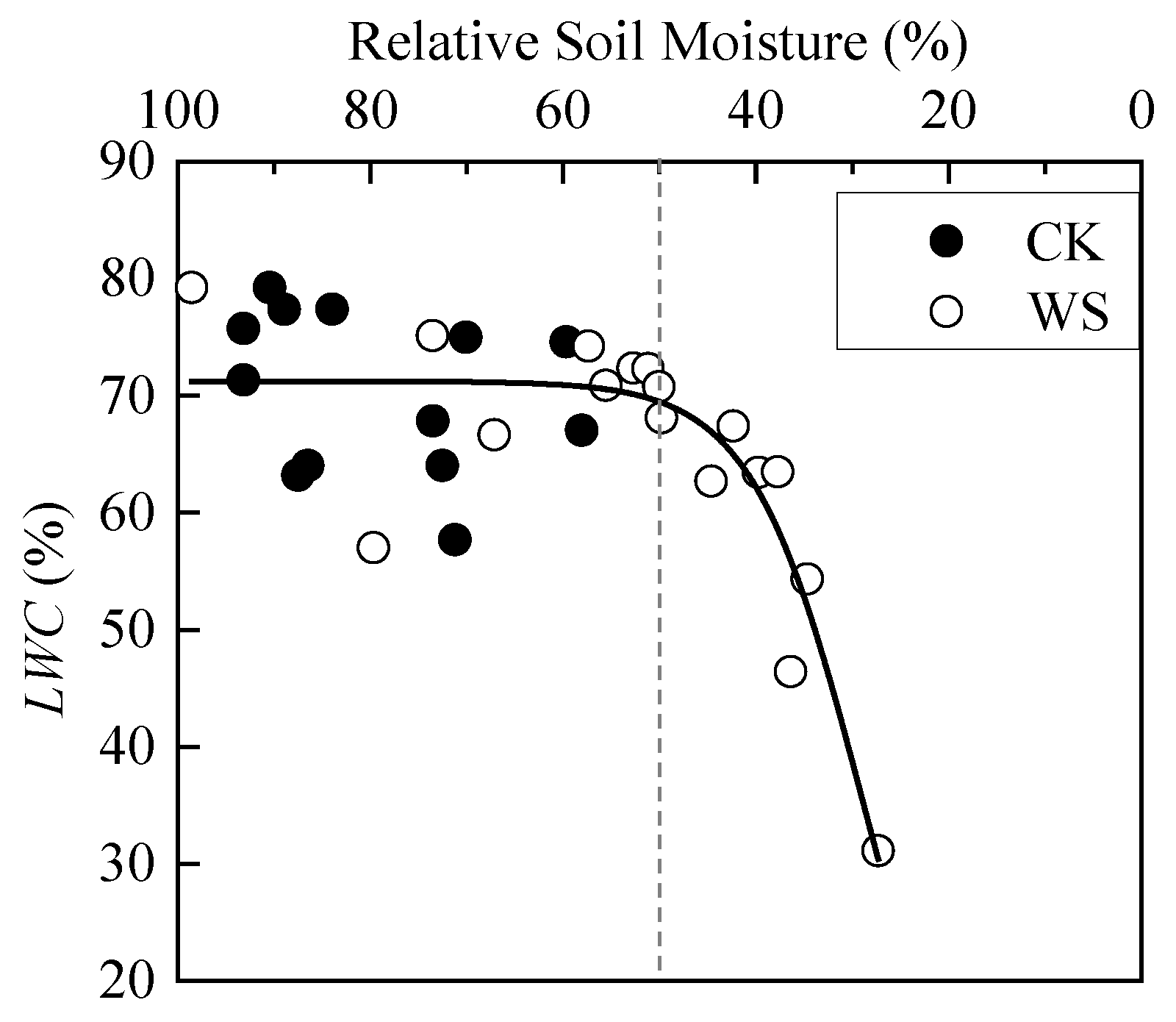
3.1.2. Response Characteristics of Water Physiological Parameters of Spring Wheat Leaves to Drought Stress
3.1.3. Response Characteristics of Photosynthetic Biochemical Parameters of Spring Wheat Leaves to Drought Stress
3.2. Drought Threshold of Spring Wheat under Rapid-Drought Stress
4. Discussion
4.1. Response Characteristics and the Corresponding Thresholds of Spring Wheat under Rapid-Drought Stress
4.2. Physiological and Biochemical Response Mechanism of Spring Wheat to Rapid Drought
4.3. Uncertainties in Research
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Orimoloye, I.R.; Belle, J.A.; Orimoloye, Y.M.; Olusola, A.O.; Ololade, O.O. Drought: A common environmental disaster. Atmosphere 2022, 13, 111. [Google Scholar] [CrossRef]
- Chatterjee, S.; Desai, A.R.; Zhu, J.; Townsend, P.A.; Huang, J. Soil moisture as an essential component for delineating and forecasting agricultural rather than meteorological drought. Remote Sens. Environ. 2022, 269, 112833. [Google Scholar] [CrossRef]
- Zhao, D.; Zhao, X.; Khongnawang, T.; Arshad, M.; Triantafilis, J.A. Vis-NIR Spectral Library to Predict Clay in Australian Cotton Growing Soil. Soil Sci. Soc. Am. J. 2018, 82, 1347–1357. [Google Scholar] [CrossRef]
- Zhao, B.; Yang, D.; Yang, S.; Santisirisomboon, J. Spatiotemporal characteristics of droughts and their propagation during the past 67 years in Northern Thailand. Atmosphere 2022, 13, 277. [Google Scholar] [CrossRef]
- Otkin, J.A.; Anderson, M.C.; Hain, C.; Mladenova, I.E.; Basara, J.B.; Svoboda, M. Examining rapid onset drought development using the thermal infrared-based evaporative stress index. J. Hydrometeorol. 2013, 14, 1057–1074. [Google Scholar] [CrossRef]
- Jin, C.; Luo, X.; Xiao, X.; Dong, J.; Li, X.; Feng, Z.; Yang, J.; Zhao, D. The 2012 flash drought threatened US Midwest agroecosystems. Chin. Geogr. Sci. 2019, 29, 768–783. [Google Scholar] [CrossRef]
- Christian, J.I.; Basara, J.B.; Hunt, E.D.; Otkin, J.A.; Furtado, J.C.; Mishra, V.; Xiao, X.; Randall, R.M. Global distribution, trends, and drivers of flash drought occurrence. Nat. Commun. 2021, 12, 6330. [Google Scholar] [CrossRef]
- Hoerling, M.; Eischeid, J.; Kumar, A.; Leung, R.; Mariotti, A.; Mo, K.; Schubert, S.; Seager, R. Causes and predictability of the 2012 Great Plains drought. Bull. Am. Meteorol. Soc. 2014, 95, 269–282. [Google Scholar] [CrossRef]
- Soltania, A.; Khooie, F.R.; Ghassemi-Golezani, K.; Moghaddam, M. Thresholds for chickpea leaf expansion and transpiration response to soil water deficit. Field Crops Res. 2000, 68, 205–210. [Google Scholar] [CrossRef]
- Yonemura, S.; Kodama, N.; Taniguchi, Y.; Ikawa, H.; Adachi, S.; Hanba, Y.T. A high-performance system of multiple gas-exchange chambers with a laser spectrometer to estimate leaf photosynthesis, stomatal conductance, and mesophyll conductance. J. Plant Res. 2019, 132, 705–718. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Flexas, J.; Ribas-Carbó, M.; Diaz-Espejo, A.; Galmés, J.; Medrano, H. Mesophyll conductance to CO2: Current knowledge and future prospects. Plant Cell Environ. 2008, 31, 602–621. [Google Scholar] [CrossRef]
- Epron, D.; Dreyer, E. Effects of severe dehydration on leaf photosynthesis in Quercus petraea (Matt.) Liebl.: Photosystem II efficiency, photochemical and nonphotochemical fluorescence quenching and electrolyte leakage. Tree Physiol. 1992, 10, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Parry, M.A.J.; Andralojc, P.J.; Khan, S.; Lea, P.J.; Keys, A.J. Rubisco activity: Effect of drought stress. Ann. Bot. 2002, 89, 833–839. [Google Scholar] [CrossRef]
- Miyazawa, S.I.; Yoshimura, S.; Shinzaki, Y.; Maeshima, M.; Miyake, C. Relationship between mesophyll CO2 gas diffusion conductance and leaf plasma-membrane-type aquaporin contents in tobacco plants grown under drought conditions. Photosynthesis 2008, 91, 805–808. [Google Scholar]
- Han, J.; Meng, H.; Wang, S.; Jiang, C.; Liu, F.; Zhang, W.; Zhang, Y. Variability of mesophyll conductance and its relationship with water use efficiency in cotton leaves under drought pretreatment. J. Plant Physiol. 2016, 194, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Barbour, M.M.; Brendel, O.; Cabrera, H.M.; Carriquí, M.; Díaz-Espejo, A.; Douthe, C.; Dreyer, E.; Ferrio, J.P.; Gago, J.; et al. Mesophyll diffusion conductance to CO2: An unappreciated central player in photosynthesis. Plant Sci. 2012, 193–194, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Centritto, M.; Loreto, F.; Chartzoulakis, K. The use of low [CO2] to estimate diffusional and non-diffusional limitations of photosynthetic capacity of salt-stressed olive saplings. Plant Cell Environ. 2003, 26, 585–594. [Google Scholar] [CrossRef]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Sheffield, J.; Wood, E.F.; Chaney, N.; Guan, K.; Sadri, S.; Yuan, X.; Olang, L.; Amani, A.; Ali, A.; Demuth, S.; et al. A drought monitoring and forecasting system for sub-sahara African water resources and food security. Bull. Am. Meteorol. Soc. 2014, 95, 861–882. [Google Scholar] [CrossRef]
- Bachmair, S.; Kohn, I.; Stahl, K. Exploring the link between drought indicators and impacts. Nat. Hazards Earth Syst. Sci. 2015, 15, 1381–1397. [Google Scholar] [CrossRef]
- Christa, D.P.; David, M.M.; Lu, S.; Dennis, P.L.; Pierre, G.; Michael, B. Advances in land surface models and indicators for drought monitoring and prediction. Bull. Am. Meteorol. Soc. 2021, 102, 1099–1122. [Google Scholar]
- Imakumbili, M.L.E.; Semu, E.; Semoka, J.M.R.; Abass, A.; Mkamilo, G. Managing cassava growth on nutrient poor soils under different water stress conditions. Heliyon 2021, 7, e07331. [Google Scholar] [CrossRef]
- AL-Quraan, N.A.; AL-Ajlouni, Z.I.; Qawasma, N.F. Physiological and biochemical characterization of the GABA shunt pathway in pea (Pisum sativum L.) seedlings under drought stress. Horticulturae 2021, 7, 125. [Google Scholar] [CrossRef]
- Maggio, A.; Raimondi, G.; Martino, A.; Pascale, S.D. Salt stress response in tomato beyond the salinity tolerance threshold. Environ. Exp. Bot. 2007, 59, 276–282. [Google Scholar] [CrossRef]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Pallardy, S.G.; Tu, K.; Law, B.E.; Wullschleger, S.D. Reliable estimation of biochemical parameters from C3 leaf photosynthesis-intercellular carbon dioxide response curves. Plant Cell Environ. 2010, 33, 1852–1874. [Google Scholar] [CrossRef]
- Harley, P.C.; Thomas, R.B.; Reynolds, J.F.; Strain, B.R. Modelling photosynthesis of cotton grown in elevated CO2. Plant Cell Environ. 1992, 15, 271–282. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Bernacchi, C.J.; Farquhar, G.D.; Singsaas, E.L. Fitting photosynthetic carbon dioxide response curves for C3 leaves. Plant Cell Environ. 2007, 30, 1035–1040. [Google Scholar] [CrossRef]
- Su, Y.; Zhu, G.; Miao, Z.; Feng, Q.; Chang, Z. Estimation of parameters of a biochemically based model of photosynthesis using a genetic algorithm. Plant Cell Environ. 2009, 32, 1710–1723. [Google Scholar] [CrossRef]
- Feng, X.; Dietze, M. Scale dependence in the effects of leaf ecophysiological traits on photosynthesis: Bayesian parameterization of photosynthesis models. New Phytol. 2013, 200, 1132–1144. [Google Scholar] [CrossRef] [PubMed]
- Vincent, C.; Schaffer, B.; Rowland, D. Water-deficit priming of papaya reduces high-light stress through oxidation avoidance rather than anti-oxidant activity. Environ. Exp. Bot. 2018, 156, 106–119. [Google Scholar] [CrossRef]
- Tcherkez, G.; Bligny, R.; Gout, E.; Mahe, A.; Hodges, M.; Cornic, G. Respiratory metabolism of illuminated leaves depends on CO2 and O2 conditions. Proc. Natl. Acad. Sci. USA 2008, 105, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Lacape, M.J.; Wery, J.; Annerose, D.J.M. Relationships between plant and soil water status in five field-grown cotton (Gossypium hirsutum L.) cultivars. Field Crops Res. 1998, 57, 29–43. [Google Scholar] [CrossRef]
- Flexasa, J.; Bota, J.; Escalona, J.M.; Sampol, B.; Medrano, H. Effects of drought on photosynthesis in grapevines under field conditions: An evaluation of stomatal and mesophyll limitations. Funct. Plant Biol. 2002, 29, 461–471. [Google Scholar] [CrossRef]
- Zhao, D.; Li, N.; Zare, E.; Wang, J.; Triantafilis, J. Mapping cation exchange capacity using a quasi-3d joint inversion of EM38 and EM31 data. Soil Tillage Res. 2020, 200, 104618. [Google Scholar] [CrossRef]
- Siddique, M.R.B.; Hamid, A.; Islam, M.S. Drought stress effects on water relations of wheat. Bot. Bull. Acad. Sin. 2000, 41, 35–39. [Google Scholar]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Perez-Martin, A.; Michelazzo, C.; Torres-Ruiz, J.M.; Flexas, J.; Fernández, J.E.; Sebastiani, L.; Diaz-Espejo, A. Regulation of photosynthesis and stomatal and mesophyll conductance under water stress and recovery in olive trees: Correlation with gene expression of carbonic anhydrase and aquaporins. J. Exp. Bot. 2014, 65, 3143–3156. [Google Scholar] [CrossRef]
- Cano, F.J.; Sánchez-Gómez, D.; Rodríguez-Calcerrada, J.; Warren, C.R.; Gil, L.; Aranda, I. Effects of drought on mesophyll conductance and photosynthetic limitations at different tree canopy layers. Plant Cell Environ. 2013, 36, 1961–1980. [Google Scholar] [CrossRef]
- Cutler, J.M.; Rains, D.W.; Loomis, R.S. The importance of cell size in the water relations of plants. Physiol. Plant. 1977, 40, 255–260. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Vassey, T.L.; Vanderveer, P.J.; Vierstra, R.D. Carbon metabolism enzymes and photosynthesis in transgenic tobacco (Nicotiana tabacum L.) having excess phytochrome. Planta 1991, 185, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Pakatas, A.; Stavrakas, D.; Fisarakis, I. Relationship between CO2 assimilation and leaf anatomical characteristics of two grapevine cultivars. Agronomie 2003, 23, 293–296. [Google Scholar] [CrossRef][Green Version]
- Terashima, I.; Hanba, Y.T.; Tholen, D.; Niinemets, Ü. Leaf functional anatomy in relation to photosynthesis. Plant Physiol. 2011, 155, 108–116. [Google Scholar] [CrossRef]
- Li, Y.; Ren, B.; Yang, X.; Xu, G.; Shen, Q.; Guo, S. Chloroplast downsizing under nitrate nutrition restrained mesophyll conductance and photosynthesis in rice (Oryza sativa L.) under drought conditions. Plant Cell Physiol. 2012, 53, 892–900. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Magnani, F. Stomatal, mesophyll conductance and biochemical limitations to photosynthesis as affected by drought and leaf ontogeny in ash and oak trees. Plant Cell Environ. 2005, 28, 834–849. [Google Scholar] [CrossRef]
- Anderegg, J.; Aasen, H.; Perich, G.; Roth, L.; Walter, A.; Hund, A. Temporal trends in canopy temperature and greenness are potential indicators of late-season drought avoidance and functional stay-green in wheat. Field Crops Res. 2021, 274, 108311. [Google Scholar] [CrossRef]
- Crawford, A.J.; McLachlan, D.H.; Hetherington, A.M.; Franklin, K.A. High temperature exposure increases plant cooling capacity. Curr. Biol. 2012, 22, R396–R397. [Google Scholar] [CrossRef]
- Medrano, H.; Parry, M.A.J.; Socias, X.; Lawlor, D.W. Long term water stress inactivates Rubisco in subterranean clover. Ann. Appl. Biol. 1997, 131, 491–501. [Google Scholar] [CrossRef]
- Escalona, J.M.; Flexas, J.; Medrano, H. Stomatal and non-stomatal limitations of photosynthesis under water stress in field-grown grapevines. Aust. J. Plant Physiol. 1999, 26, 421–433. [Google Scholar] [CrossRef]
- Pilon, C.; Snider, J.L.; Sobolev, V.; Chastain, D.R.; Sorensen, R.B.; Meeks, C.D.; Massa, A.N.; Walk, T.; Singh, B.; Earl, H.J. Assessing stomatal and non-stomatal limitations to carbon assimilation under progressive drought in peanut (Arachis hypogaea L.). J. Plant Physiol. 2018, 231, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Baldocchi, D.D. Seasonal trends in photosynthetic parameters and stomatal conductance of blue oak (Quercus douglasii) under prolonged summer drought and high temperature. Tree Physiol. 2003, 23, 865–877. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Gu, L.; Dickinson, R.E.; Pallardy, S.G.; Baker, J.; Cao, Y.; Da Matta, F.M.; Dong, X.; Ellsworth, D.; Goethem, D.V.; et al. Asymmetrical effects of mesophyll conductance on fundamental photosynthetic parameters and their relationships estimated from leaf gas exchange measurements. Plant Cell Environ. 2014, 37, 978–994. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P.; Bernacchi, C.J. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? procedures and sources of error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef] [PubMed]
- von Caemmerer, S. Steady-state models of photosynthesis. Plant Cell Environ. 2013, 36, 1617–1630. [Google Scholar] [CrossRef]
- Bernacchi, C.J.; Singsaas, E.L.; Pimentel, C.; Portis, J.A.R.; Long, S.P. Improved temperature response functions for models of Rubisco-limited photosynthesis. Plant Cell Environ. 2001, 24, 253–259. [Google Scholar] [CrossRef]
- Flexas, J.; Niinemets, U.; Gallé, A.; Barbour, M.M.; Centritto, M.; Diaz-Espejo, A.; Douthe, C.; Galmés, J.; Ribas-Carbo, M.; Rodriguez, P.L.; et al. Diffusional conductances to CO2 as a target for increasing photosynthesis and photosynthetic water-use efficiency. Photosynth. Res. 2013, 117, 45–59. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, J.; Tischer, S.V.; Christmann, A.; Windisch, W.; Schnyder, H.; Grill, E. Leveraging abscisic acid receptors for efficient water use in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 113, 6791–6796. [Google Scholar] [CrossRef]
- Evans, J.R.; Sharkey, T.D.; Berry, J.A.; Farquhar, G.D. Carbon isotope discrimination measured concurrently with gas exchange to investigate CO2 diffusion in leaves of higher plants. Aust. J. Plant Physiol. 1986, 13, 281–292. [Google Scholar] [CrossRef]
- Marco, G.D.; Manes, F.; Tricoli, D.; Vitale, E. Fluorescence parameters measured concurrently with net photosynthesis to investigate chloroplastic CO2 concentration in leaves of Quercus ilex L. J. Plant Physiol. 1990, 136, 538–543. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Treatments | N | Mean | Standard Deviation | F | Sig. |
|---|---|---|---|---|---|---|
| Pn (μmol·m−2·s−1) | CK | 15 | 22.6 | 3.2 | 14.5 | 0.000 ** |
| WS | 35 | 14.6 | 7.8 | |||
| Tr (mmol·m−2·s−1) | CK | 15 | 5.5 | 1.0 | 10.1 | 0.002 ** |
| WS | 35 | 3.7 | 2.1 | |||
| gs (mol·m−2·s−1) | CK | 15 | 0.4 | 0.1 | 17.1 | 0.000 ** |
| WS | 35 | 0.2 | 0.2 | |||
| Ci (μmol·mol−1) | CK | 15 | 270.9 | 12.9 | 6.8 | 0.012 * |
| WS | 35 | 236.7 | 49.8 |
| Parameter | Fitted Equation | Adj. R2 | F | Sig. |
|---|---|---|---|---|
| Pn | 0.7 | 144.3 | 0.000 ** | |
| Tr | 0.5 | 88.8 | 0.000 ** | |
| gs | 0.6 | 69.0 | 0.000 ** | |
| Ci | 0.1 | 217.8 | 0.000 ** |
| Parameter | Treatments | N | Mean | Standard Deviation | F | Sig. |
|---|---|---|---|---|---|---|
| LWC (%) | CK | 13 | 70.4 | 6.8 | 2.6 | 0.122 |
| WS | 17 | 64.5 | 11.9 | |||
| M50% | 22 | 70.6 | 6.5 | 14.8 | 0.001 ** | |
| L50% | 8 | 57.1 | 12.8 |
| Parameter | Fitted Equation | Adj. R2 | F | Sig. |
|---|---|---|---|---|
| LWC | 0.8 | 709.2 | 0.000 ** |
| Parameter | Treatments | N | Mean | Standard Deviation | F | Sig. |
|---|---|---|---|---|---|---|
| Vcmax (μmol·m−2·s−1) | CK | 15 | 75.0 | 14.1 | 4.2 | 0.048 * |
| WS | 21 | 60.9 | 23.9 | |||
| Jmax (μmol·m−2·s−1) | CK | 18 | 191.8 | 18.1 | 9.8 | 0.003 ** |
| WS | 30 | 146.4 | 59.7 | |||
| TPU (μmol·m−2·s−1) | CK | 19 | 13.9 | 2.3 | 11.0 | 0.002 ** |
| WS | 44 | 9.2 | 6.0 | |||
| gm (μmol·m-2·s−1·Pa−1) | CK | 10 | 2.1 | 0.9 | 9.8 | 0.004 ** |
| WS | 27 | 1.0 | 0.9 |
| Parameter | Fitted Equation | Adj. R2 | F | Sig. |
|---|---|---|---|---|
| Vcmax | 0.4 | 79.1 | 0.000 ** | |
| Jmax | 0.4 | 117.1 | 0.000 ** | |
| TPU | 0.7 | 142.7 | 0.000 ** | |
| gm | 0.5 | 29.8 | 0.000 ** |
| Parameter | Mild Drought | Severe Drought | ||||
|---|---|---|---|---|---|---|
| Drought Thresholds | Disaster Degree | Drought Thresholds | Disaster Degree | |||
| (%) | RR (%) | (%) | RR (%) | |||
| Pn (μmol·m−2·s−1) | 48.9 | 18.2 | 19.6 | 35.7 | 10.9 | 51.6 |
| Tr (mmol·m−2·s−1) | 42.7 | 4.1 | 25.4 | 34.2 | 2.6 | 53.0 |
| gs (mol·m−2·s−1) | 56.6 | 0.3 | 18.7 | 43.6 | 0.2 | 48.0 |
| Ci (μmol·mol−1) | 58.2 | 249.2 | 8.0 | 36.8 | 218.6 | 19.3 |
| LWC (%) | 45.3 | 67.4 | 4.2 | 29.0 | 35.6 | 49.4 |
| Vcmax (μmol·m−2·s−1) | 42.3 | 65.7 | 12.4 | 28.6 | 34.2 | 54.4 |
| Jmax (μmol·m−2·s−1) | 53.3 | 171.3 | 10.7 | 28.2 | 88.1 | 54.1 |
| TPU (μmol·m−2·s−1) | 43.7 | 11.2 | 19.7 | 34.2 | 6.8 | 51.0 |
| gm (μmol·m−2·s−1·Pa−1) | 50.9 | 1.4 | 30.4 | 38.3 | 0.9 | 56.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, F.; Wang, H.; Zhao, F.; Wang, R.; Qi, Y.; Zhang, K.; Zhao, H.; Tang, G.; Yang, Y. The Response Mechanism and Threshold of Spring Wheat to Rapid Drought. Atmosphere 2022, 13, 596. https://doi.org/10.3390/atmos13040596
Chen F, Wang H, Zhao F, Wang R, Qi Y, Zhang K, Zhao H, Tang G, Yang Y. The Response Mechanism and Threshold of Spring Wheat to Rapid Drought. Atmosphere. 2022; 13(4):596. https://doi.org/10.3390/atmos13040596
Chicago/Turabian StyleChen, Fei, Heling Wang, Funian Zhao, Runyuan Wang, Yue Qi, Kai Zhang, Hong Zhao, Guoying Tang, and Yang Yang. 2022. "The Response Mechanism and Threshold of Spring Wheat to Rapid Drought" Atmosphere 13, no. 4: 596. https://doi.org/10.3390/atmos13040596
APA StyleChen, F., Wang, H., Zhao, F., Wang, R., Qi, Y., Zhang, K., Zhao, H., Tang, G., & Yang, Y. (2022). The Response Mechanism and Threshold of Spring Wheat to Rapid Drought. Atmosphere, 13(4), 596. https://doi.org/10.3390/atmos13040596