L Chromosome Behaviour and Chromosomal Imprinting in Sciara Coprophila



Abstract
1. Introduction
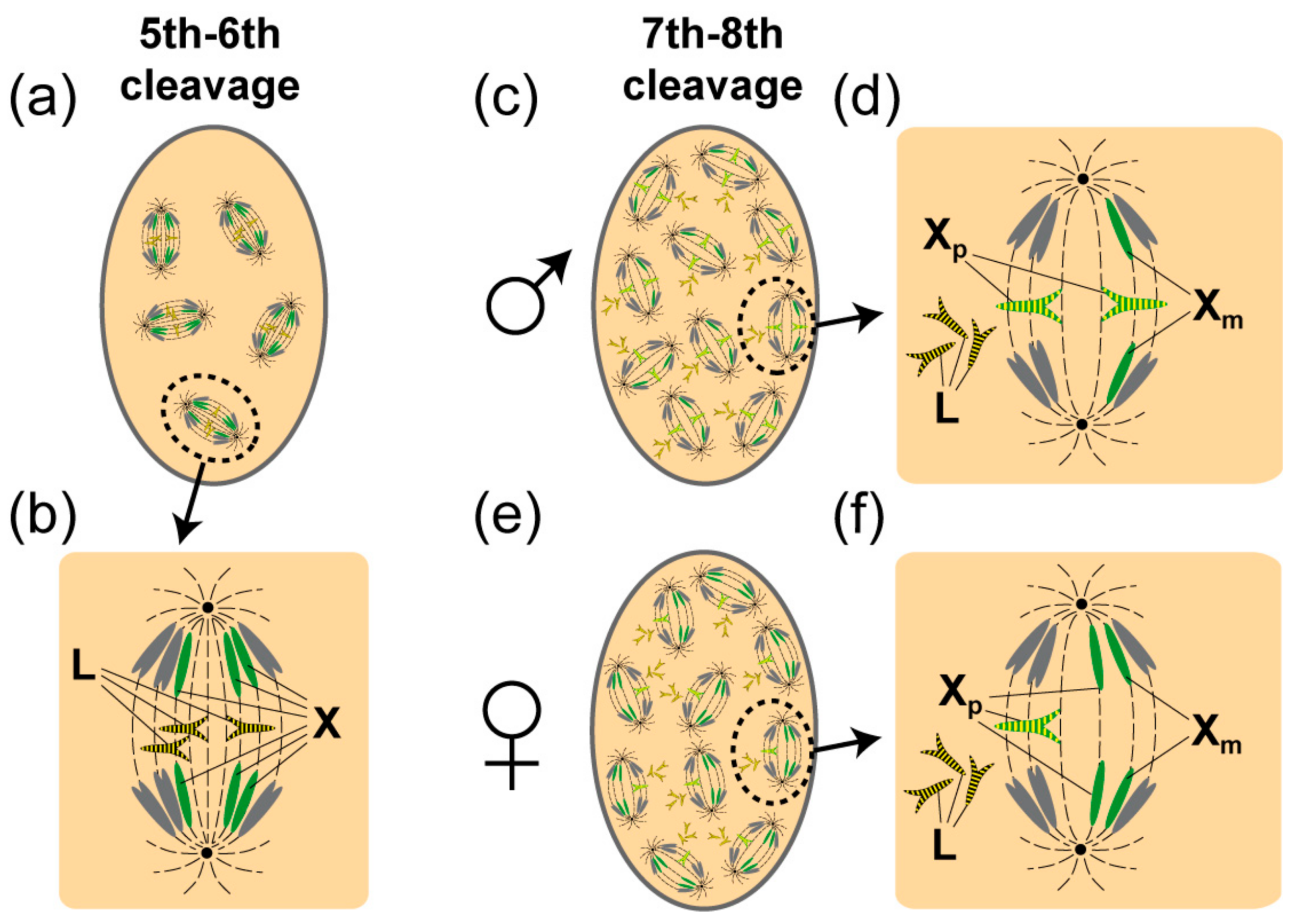
1.1. L Chromosome Elimination in the Soma
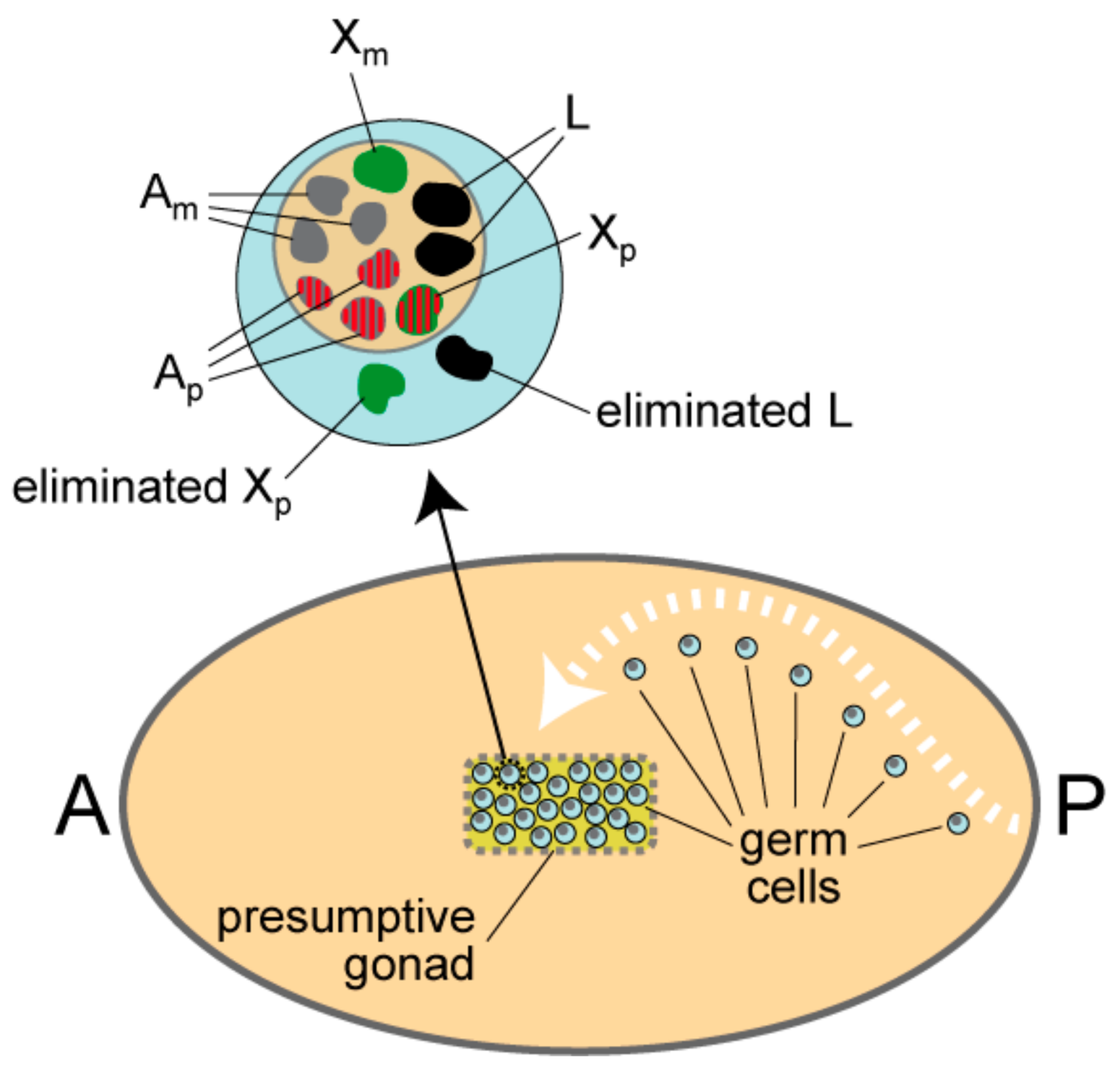
1.2. L Chromosome Elimination in the Germ-Line
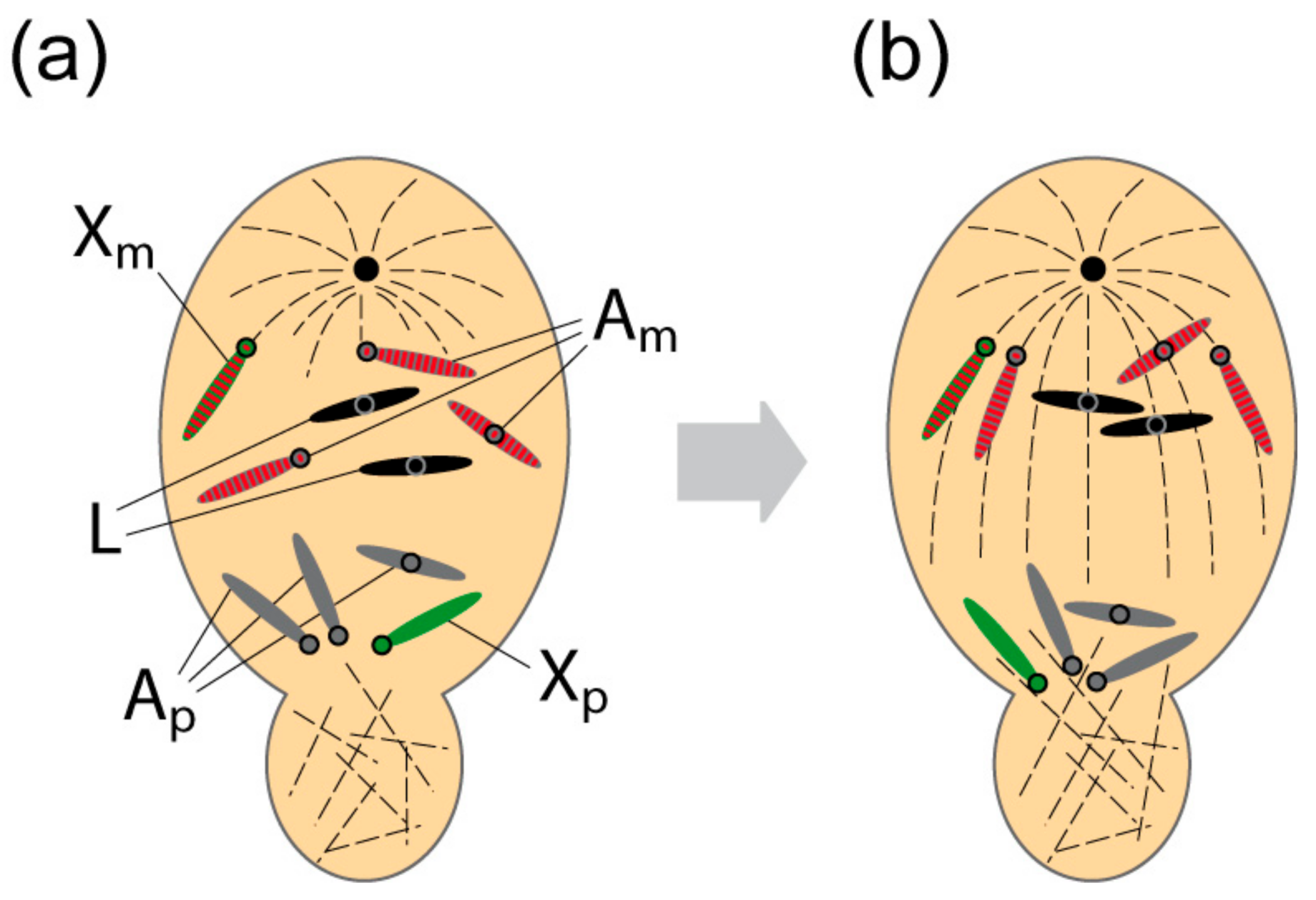
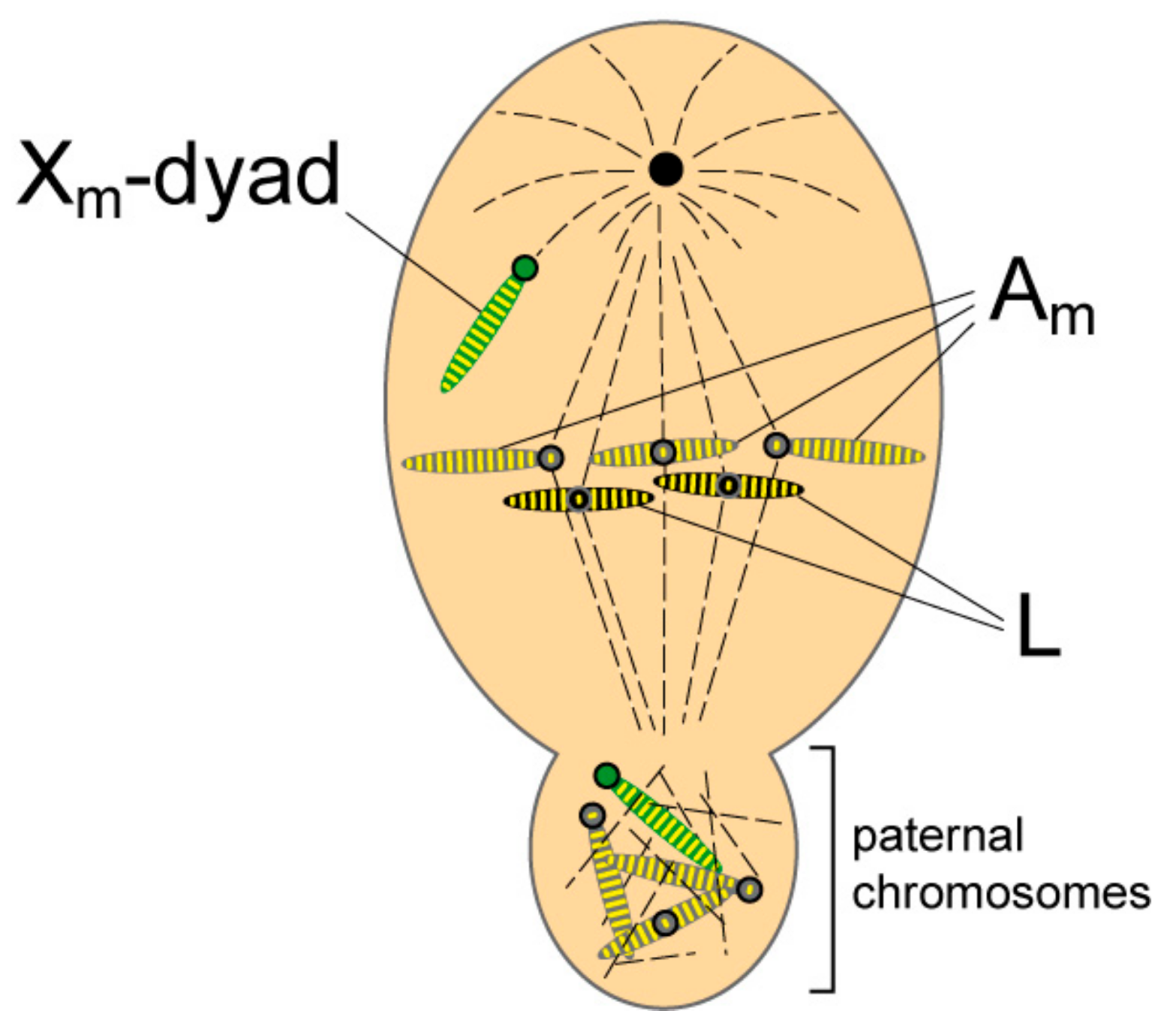
1.3. L Chromosome Behaviour During Male Meiosis
2. The Epigenotype of L Chromosomes
3. L Chromosomes and Chromosomal Imprinting of the Regular Chromosomes
3.1. Erasure of Parental Imprints and Selective Retention of L Chromosomes and Regular Maternal Chromosomes at the First Spermatocyte Division
3.2. L Chromosomes and Imprinting of the Controlling Element
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Metz, C.W.C. Chromosome behaviour, inheritance and sex determination in Sciara. Am. Nat. 1938, 72, 485–520. [Google Scholar] [CrossRef]
- Gerbi, S.A. Unusual chromosome movements in sciarid flies. Results Probl. Cell Differ. 1986, 13, 71–104. [Google Scholar] [PubMed]
- Goday, C.; Esteban, M.R. Chromosome elimination in sciarid flies. Bioessays 2001, 23, 242–250. [Google Scholar] [CrossRef]
- Crouse, H.V. The nature of the influence of X-translocations on sex of progeny in Sciara coprophila. Chromosoma 1960, 11, 146–166. [Google Scholar] [CrossRef] [PubMed]
- DuBois, A.M. Chromosome behaviour during cleavage in the eggs of Sciara coprophila (Diptera) in relation to the problem of sex determination. Z Zellf mikr Anat 1933, 19, 595–614. [Google Scholar]
- Rieffel, S.M.; Crouse, H.V. The elimination and differentiation of chromosomes in the germ line of Sciara. Chromosoma 1966, 19, 231–276. [Google Scholar] [CrossRef] [PubMed]
- Berry, R.O. Chromosome behaviour in the germ cells and development of the gonads of Sciara ocellaris. J. Morph. 1941, 68, 547–583. [Google Scholar] [CrossRef]
- Singh, P.B.; Georgatos, S.D. HP1: Facts, open questions and speculation. J. Struct. Biol. 2002, 140, 10–16. [Google Scholar] [CrossRef]
- Goday, C.; Ruiz, M.F. Differential acetylation of histones H3 and H4 in paternal and maternal germline chromosomes during development of sciarid flies. J. Cell Sci. 2002, 115, 4765–4775. [Google Scholar] [CrossRef] [PubMed]
- Metz, C.W. An apparent case of monocentric mitosis in Sciara (Diptera). Science 1926, 63, 190–191. [Google Scholar] [CrossRef] [PubMed]
- Crouse, H.V.; Brown, A.; Mumford, B.C. L-chromosome inheritance and the problem of chromosome “imprinting” in Sciara (Sciaridae, Diptera). Chromosoma 1971, 34, 324–339. [Google Scholar] [CrossRef] [PubMed]
- Amabis, J.M.; Reinach, F.C.; Andrews, N. Spermatogenesis in Trichosia pubescens (Diptera:Sciaridae). J. Cell Sci. 1979, 36, 199–213. [Google Scholar] [PubMed]
- Greciano, P.G.; Goday, C. Methylation of histone H3 at Lys4 differs between paternal and maternal chromosomes in Sciara ocellaris germline development. J. Cell Sci. 2006, 119, 4667–4677. [Google Scholar] [CrossRef] [PubMed]
- Greciano, P.G.; Ruiz, M.F.; Kremer, L.; Goday, C. Two new chromodomain-containing proteins that associate with heterochromatin in Sciara coprophila chromosomes. Chromosoma 2009, 118, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Escriba, M.C.; Giardini, M.C.; Goday, C. Histone H3 phosphorylation and non-disjunction of the maternal X chromosome during male meiosis in sciarid flies. J. Cell Sci. 2011, 124, 1715–1725. [Google Scholar] [CrossRef] [PubMed]
- Escriba, M.C.; Goday, C. Histone X3 phosphorylation and elimination of paternal X chromosomes at early cleavages in sciarid flies. J. Cell Sci. 2013, 126, 3214–3222. [Google Scholar] [CrossRef] [PubMed]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Jaenisch, R.; Bird, A. Epigenetic regulation of gene expression: How the genome integrates intrinsic and environmental signals. Nat. Genet. 2003, 33, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Saksouk, N.; Simboeck, E.; Dejardin, J. Constitutive heterochromatin formation and transcription in mammals. Epigenetics Chromatin 2015, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Crouse, H.V. The controlling element in sex chromosome behaviour in Sciara. Genetics 1960, 45, 1429–1443. [Google Scholar] [PubMed]
- Metz, C.W. Factors influencing chromosome movements in mitosis. Cytologia 1936, 7, 219–231. [Google Scholar] [CrossRef]
- Kubai, D.F. Meiosis in Sciara coprophila: Structure of the spindle and chromosome behaviour during the first meiotic division. J. Cell Biol. 1982, 93, 655–669. [Google Scholar] [CrossRef] [PubMed]
- Kubai, D.F. Nonrandom chromosome arrangements in germ line nuclei of Sciara coprophila males: The basis for nonrandom chromosome segregation on the meiosis I spindle. J. Cell Biol. 1987, 105, 2433–2446. [Google Scholar] [CrossRef] [PubMed]
- Chandra, H.S.; Brown, S.W. Chromosome imprinting and the mammalian X chromosome. Nature 1975, 253, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.B. Heterochromatin and the molecular mechanisms of “parent-of-origin” effects in animals. J. Biosci. 2016, 41, 759–786. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Epigenetic Modifications |
|---|---|
| First somatic division | L chromosomes are positive for H4K8Ac [9] |
| Syncytial embryo before elimination | L chromosomes are positive for H3S10P at pro-metaphase [16]. Proceeding to metaphase they become positive for H3S28P [16]. The centromeres are specifically stained with H3T3P antibodies at metaphase [16]. |
| Syncytial embryo during elimination | L chromosome arms are stained with H3S10P at metaphase/anaphase [16]. By contrast, the regulars have lost their H3S10P staining at anaphase. |
| Resting stage germ cells before elimination of Xp | L chromosomes are negative for H4K8Ac, H4K12Ac and PanH3Ac [9]. |
| Resting stage germ cells after elimination of Xp | L chromosomes are negative for H4K8Ac, H4K12Ac and PanH3Ac [9]. |
| Germ cells undergoing larval mitosis I (9th day after oviposition) | L chromosomes are negative for H4K8Ac [9]. They are positive for H3K9me2, H3K9me3, H4K20me3, 5MeC and the two HP1-like proteins ScoHET1 and ScoHET2 [13]. |
| Germ cells at end of mitosis III in third instar larvae (12th day after oviposition) | L chromosomes are negative for H4K8Ac [9]. |
| Prophase of Meiosis I | L chromosomes are negative for H3S10P, H3S28P and H3T3P but positive for H3T11P [15] |
| Meiosis I | L chromosomes are negative for H3S10P, H3S28P and H3T3P but positive for H3T11P [15]. They are also negative for H4K8Ac [9]. |
| Meiosis II | L chromosomes are positive for H3S10P, H3S28P and H3T3P and H3T11P [15]. They are also positive for ScoHET1 [13]. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, P.B.; Belyakin, S.N. L Chromosome Behaviour and Chromosomal Imprinting in Sciara Coprophila. Genes 2018, 9, 440. https://doi.org/10.3390/genes9090440
Singh PB, Belyakin SN. L Chromosome Behaviour and Chromosomal Imprinting in Sciara Coprophila. Genes. 2018; 9(9):440. https://doi.org/10.3390/genes9090440
Chicago/Turabian StyleSingh, Prim B., and Stepan N. Belyakin. 2018. "L Chromosome Behaviour and Chromosomal Imprinting in Sciara Coprophila" Genes 9, no. 9: 440. https://doi.org/10.3390/genes9090440
APA StyleSingh, P. B., & Belyakin, S. N. (2018). L Chromosome Behaviour and Chromosomal Imprinting in Sciara Coprophila. Genes, 9(9), 440. https://doi.org/10.3390/genes9090440