Seasonal and Sexual Differences in the Microbiota of the Hoopoe Uropygial Secretion

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area, Study Species, and Sampling Procedures
2.2. Fluorescence In Situ Hybridization
2.3. DNA Extraction and Purification
2.4. DNA Quantification by Quantitative Polymerase Chain Reaction
2.5. Amplicon Libraries Construction and Pyrosequencing
2.6. Sequence Data Analysis and Taxonomic Identification
2.7. Statistical Analyses
3. Results
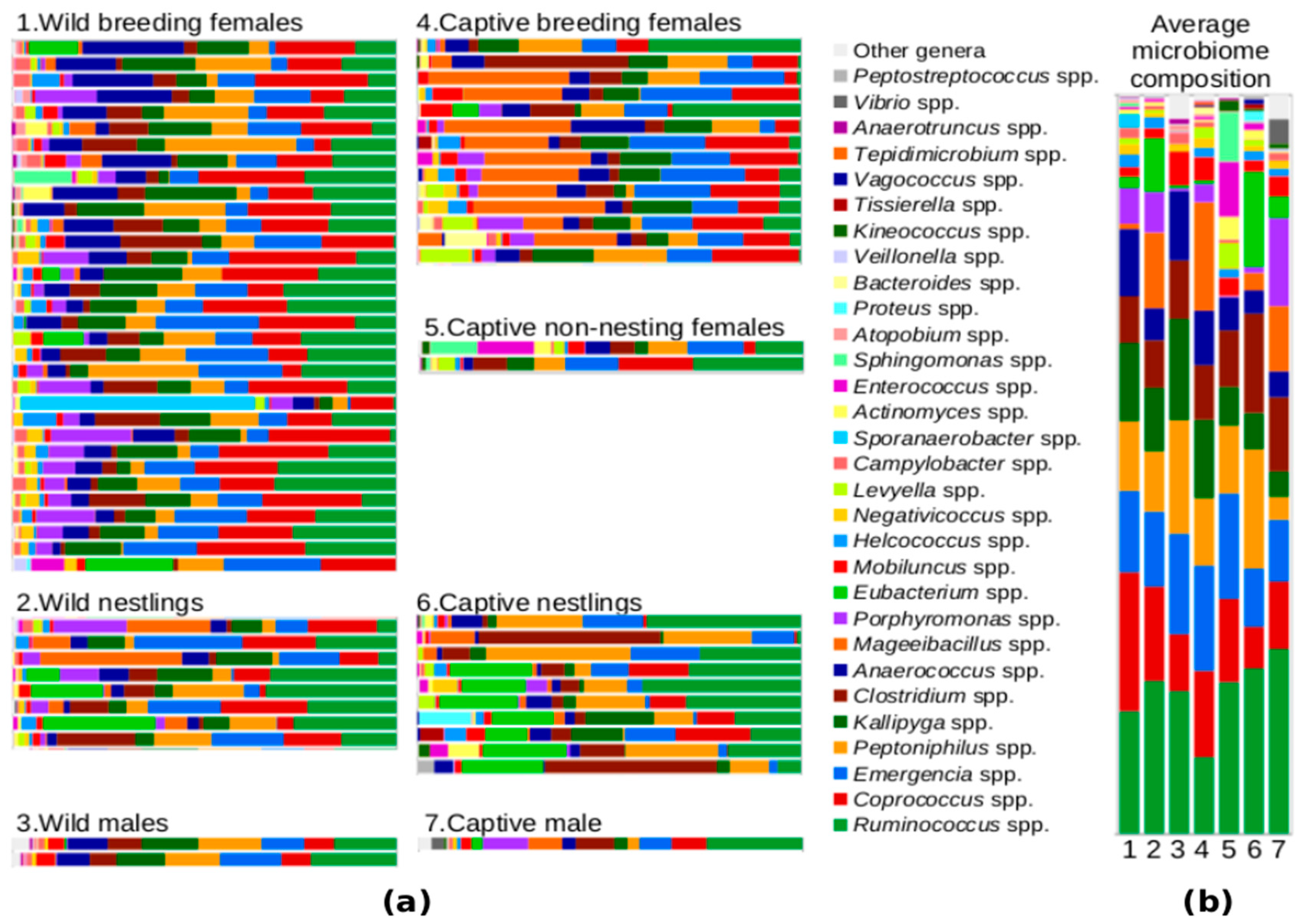
3.1. The Uropygial Microbiota during the Breeding Season
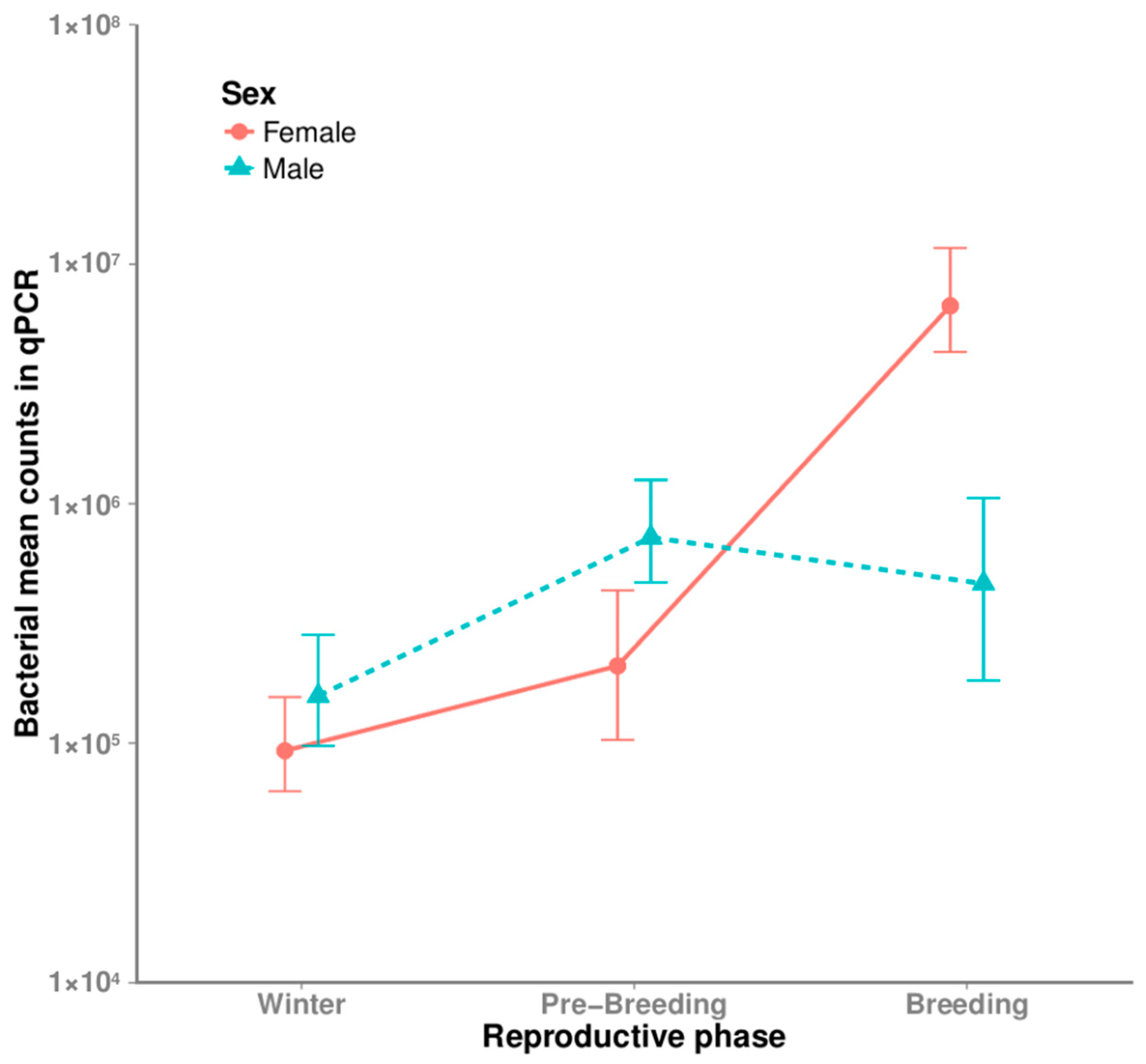
3.2. Seasonal and Sexual Differences in Bacterial Densities
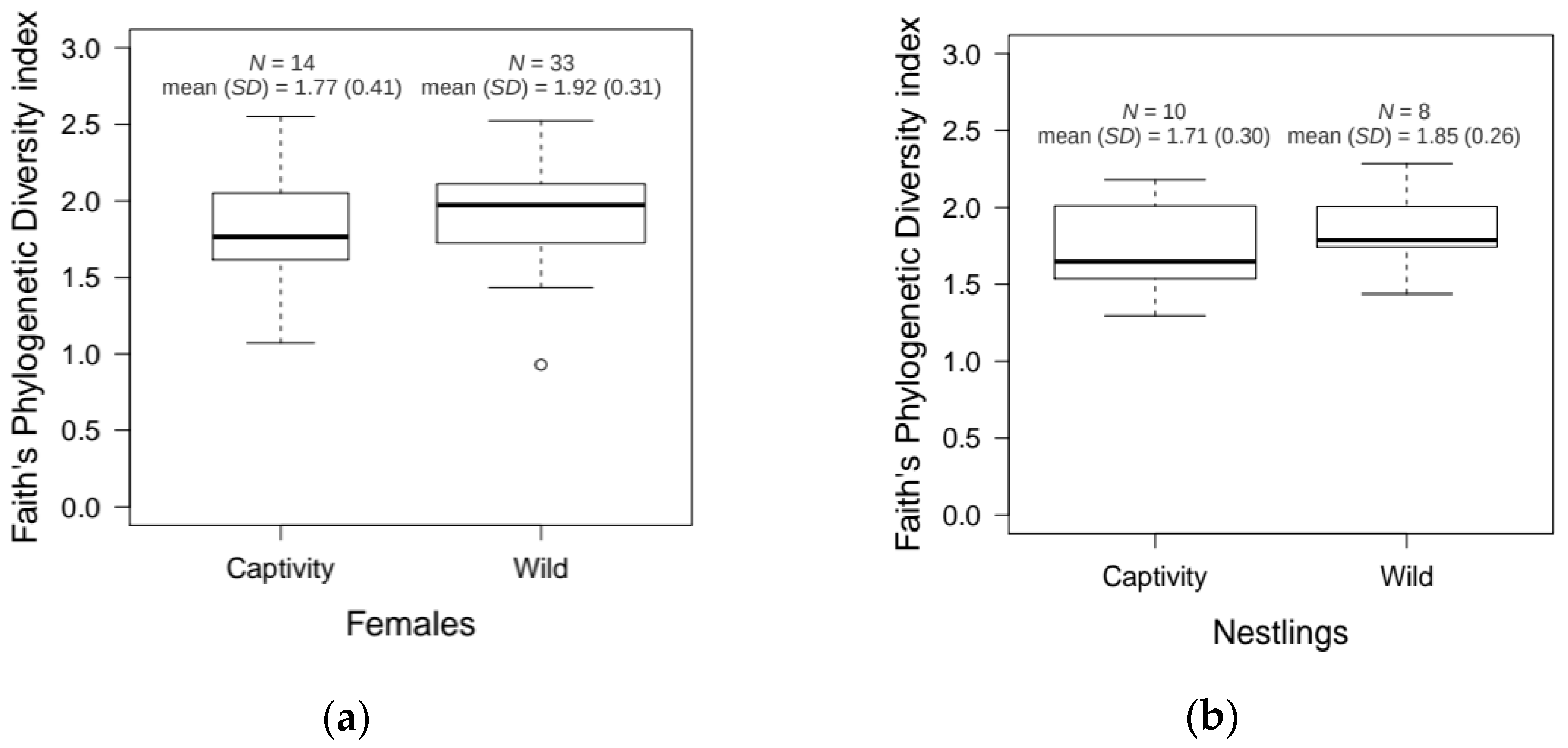
3.3. Age and Environmental Effects in the Hoopoe Uropygial Microbiota of Nesting Individuals
4. Discussion
4.1. Composition of the Uropygial Microbiota: Clostridia as Key Partners
4.2. The Uropygial Microbiota of Nesting Hoopoes: Age and Environmental Contributions
4.3. Hoopoe Reproductive Behavior and the Microbiota Dynamics and Functions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Fluorescence In Situ Hybridization: Hybridization and DNA Labeling with Hoechst
Appendix B
Fluorescence In Situ Hybridization: Quantification
References
- Moran, N.A. Symbiosis. Curr. Biol. 2006, 16, R866–R871. [Google Scholar] [CrossRef] [PubMed]
- Moya, A.; Peretó, J.; Gil, R.; Latorre, A. Learning how to live together: Genomic insights into prokaryote-animal symbioses. Nat. Rev. Genet. 2008, 9, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Bordenstein, S.R.; Theis, K.R. Host biology in light of the microbiome: Ten principles of holobionts and hologenomes. PLoS Biol. 2015, 13, e1002226. [Google Scholar] [CrossRef] [PubMed]
- Flórez, L.V.; Biedermann, P.H.W.; Engl, T.; Kaltenpoth, M. Defensive symbioses of animals with prokaryotic and eukaryotic microorganisms. Nat. Prod. Rep. 2015, 32, 904–936. [Google Scholar] [CrossRef] [PubMed]
- Hird, S.M. Evolutionary biology needs wild microbiomes. Front. Microbiol. 2017, 8, 725. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.T.; Sharma, V.; Elmén, L.; Peterson, S.N. Immune homeostasis, dysbiosis and therapeutic modulation of the gut microbiota. Clin. Exp. Immunol. 2015, 179, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Rajilić-Stojanović, M. Function of the microbiota. Best Pract. Res. Clin. Gastroenterol. 2013, 27, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Gordon, J.I. Commensal host-bacterial relationships in the gut. Science 2001, 292, 1115–1118. [Google Scholar] [CrossRef] [PubMed]
- Leser, T.D.; Mølbak, L. Better living through microbial action: The benefits of the mammalian gastrointestinal microbiota on the host. Environ. Microbiol. 2009, 11, 2194–2206. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.; Ziswiler, V. The uropygial gland. In Avian Biology; Academic Press: London, UK, 1982. [Google Scholar]
- Moreno-Rueda, G. Preen oil and bird fitness: A critical review of the evidence. Biol. Rev. 2017, 92, 2131–2143. [Google Scholar] [CrossRef] [PubMed]
- Hagelin, J.C.; Jones, I.L. Bird odors and other chemical substances: A defense mechanism or overlooked mode of intraspecific communication? Auk Waco 2007, 124, 741–761. [Google Scholar] [CrossRef]
- Law-Brown, J.; Meyers, P.R. Enterococcus phoeniculicola sp. nov., a novel member of the enterococci isolated from the uropygial gland of the red-billed woodhoopoe, Phoeniculus purpureus. Int. J. Syst. Evol. Microbiol. 2003, 53, 683–685. [Google Scholar] [CrossRef] [PubMed]
- Martín-Vivaldi, M.; Peña, A.; Peralta-Sánchez, J.M.; Sánchez, L.; Ananou, S.; Ruiz-Rodríguez, M.; Soler, J.J. Antimicrobial chemicals in hoopoe preen secretions are produced by symbiotic bacteria. Proc. R. Soc. B Biol. Sci. 2010, 277, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Soler, J.J.; Martín-Vivaldi, M.; Ruiz-Rodríguez, M.; Valdivia, E.; Martín-Platero, A.M.; Martínez-Bueno, M.; Peralta-Sánchez, J.M.; Méndez, M. Symbiotic association between hoopoes and antibiotic-producing bacteria that live in their uropygial gland. Funct. Ecol. 2008, 22, 864–871. [Google Scholar] [CrossRef]
- Law-Brown, J. Chemical Defence in the Red-Billed Wood Hoopoe: Phoeniculus purpureus. Master’s Thesis, University of Cape Town, Rondebosch, South Africa, 2001. [Google Scholar]
- Soler, J.J.; Martín-Vivaldi, M.; Peralta-Sánchez, J.M.; Ruiz-Rodríguez, M. Antibiotic-producing bacteria as a possible defence of birds against pathogenic microorganisms. Open Ornithol. J. 2010, 3, 93–100. [Google Scholar] [CrossRef]
- Kristin, A. Family Upupidae (Hoopoes). In Handbook of the Birds of the World; Lynx Edicions: Barcelona, Spain, 2001; Volume 6, pp. 396–411. [Google Scholar]
- Ligon, J.D. Family Phoeniculidae (Wood-hoopoes). In Handbook of the Birds of the World; Lynx Edicions: Barcelona, Spain, 2001; Volume 6, pp. 412–434. [Google Scholar]
- Martín-Platero, A.M.; Valdivia, E.; Soler, J.J.; Martín-Vivaldi, M.; Maqueda, M.; Martínez-Bueno, M. Characterization of antimicrobial substances produced by Enterococcus faecalis MRR 10-3, isolated from the uropygial gland of the hoopoe (Upupa epops). Appl. Environ. Microbiol. 2006, 72, 4245–4249. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Rodríguez, M.; Valdivia, E.; Martín-Vivaldi, M.; Martín-Platero, A.M.; Martínez-Bueno, M.; Méndez, M.; Peralta-Sánchez, J.M.; Soler, J.J. Antimicrobial activity and genetic profile of enteroccoci isolated from hoopoes uropygial gland. PLoS ONE 2012, 7, e41843. [Google Scholar] [CrossRef] [PubMed]
- Martín-Vivaldi, M.; Ruiz-Rodríguez, M.; José Soler, J.; Manuel Peralta-Sánchez, J.; Méndez, M.; Valdivia, E.; Manuel Martín-Platero, A.; Martínez-Bueno, M. Seasonal, sexual and developmental differences in hoopoe Upupa epops preen gland morphology and secretions: Evidence for a role of bacteria. J. Avian Biol. 2009, 40, 191–205. [Google Scholar] [CrossRef]
- Martín-Vivaldi, M.; Soler, J.J.; Peralta-Sánchez, J.M.; Arco, L.; Martín-Platero, A.M.; Martínez-Bueno, M.; Ruiz-Rodríguez, M.; Valdivia, E. Special structures of hoopoe eggshells enhance the adhesion of symbiont-carrying uropygial secretion that increase hatching success. J. Anim. Ecol. 2014, 83, 1289–1301. [Google Scholar] [CrossRef] [PubMed]
- Soler, J.J.; Martín-Vivaldi, M.; Peralta-Sánchez, J.M.; Arco, L.; Juárez-García-Pelayo, N. Hoopoes color their eggs with antimicrobial uropygial secretions. Naturwissenschaften 2014, 101, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Ministerio de la Presidencia. REAL DECRETO 1201/2005, de 10 de octubre, sobre protección de los animales utilizados para experimentación y otros fines científicos. Boletín Oficial del Estado (BOE) 2005, 252, 34367–34391. [Google Scholar]
- Amann, R.I.; Binder, B.J.; Olson, R.J.; Chisholm, S.W.; Devereux, R.; Stahl, D.A. Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl. Environ. Microbiol. 1990, 56, 1919–1925. [Google Scholar] [PubMed]
- Wellinghausen, N.; Bartel, M.; Essig, A.; Poppert, S. Rapid identification of clinically relevant Enterococcus species by Fluorescence In Situ Hybridization. J. Clin. Microbiol. 2007, 45, 3424–3426. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schüffler, P.J.; Fuchs, T.J.; Ong, C.S.; Wild, P.J.; Rupp, N.J.; Buhmann, J.M. TMARKER: A free software toolkit for histopathological cell counting and staining estimation. J. Pathol. Inform. 2013, 4, S2. [Google Scholar] [CrossRef] [PubMed]
- Klammer, S.; Knapp, B.; Insam, H.; Dell’Abate, M.T.; Ros, M. Bacterial community patterns and thermal analyses of composts of various origins. Waste Manag. Res. 2008, 26, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [PubMed]
- Větrovský, T.; Baldrian, P. The variability of the 16S rRNA gene in bacterial genomes and its consequences for bacterial community analyses. PLoS ONE 2013, 8, e57923. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Hamady, M.; Lauber, C.L.; Knight, R. The influence of sex, handedness, and washing on the diversity of hand surface bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 17994–17999. [Google Scholar] [CrossRef] [PubMed]
- Huse, S.M.; Dethlefsen, L.; Huber, J.A.; Welch, D.M.; Relman, D.A.; Sogin, M.L. Exploring microbial diversity and taxonomy using SSU rRNA hypervariable tag sequencing. PLoS Genet. 2008, 4, e1000255. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Preheim, S.P.; Perrotta, A.R.; Martín-Platero, A.M.; Gupta, A.; Alm, E.J. Distribution-Based Clustering: Using ecology to refine the Operational Taxonomic Unit. Appl. Environ. Microbiol. 2013, 79, 6593–6603. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- McDonald, J.H. Handbook of Biological Statistics, 3rd ed.; Sparky House Publishing: Baltimore, MD, USA, 2014. [Google Scholar]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Faith, D.P.; Baker, A.M. Phylogenetic diversity (PD) and biodiversity conservation: Some bioinformatics challenges. Evol. Bioinform. Online 2007, 2, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package; The Comprehensive R Archive Network CRAN. 2013. Available online: https://www.cran.r-project.org/ (accessed on 1 August 2018).
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009; ISBN 978-0-387-98140-6. [Google Scholar]
- Vázquez-Baeza, Y.; Pirrung, M.; Gonzalez, A.; Knight, R. EMPeror: A tool for visualizing high-throughput microbial community data. GigaScience 2013, 2, 16. [Google Scholar] [CrossRef] [PubMed]
- Pedrós-Alió, C. The rare bacterial biosphere. Annu. Rev. Mar. Sci. 2012, 4, 449–466. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ruano, S.M.; Martín-Vivaldi, M.; Martín-Platero, A.M.; López-López, J.P.; Peralta-Sánchez, J.M.; Ruiz-Rodríguez, M.; Soler, J.J.; Valdivia, E.; Martínez-Bueno, M. The hoopoe’s uropygial gland hosts a bacterial community influenced by the living conditions of the bird. PLoS ONE 2015, 10, e0139734. [Google Scholar] [CrossRef] [PubMed]
- Lopetuso, L.R.; Scaldaferri, F.; Petito, V.; Gasbarrini, A. Commensal Clostridia: Leading players in the maintenance of gut homeostasis. Gut Pathog. 2013, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, Á.; Martín-Vivaldi, M.; Ruiz-Rodríguez, M.; Martínez-Bueno, M.; Arco, L.; Rodríguez-Ruano, S.M.; Peralta-Sánchez, J.M.; Soler, J.J. The microbiome of the uropygial secretion in hoopoes is shaped along the nesting phase. Microb. Ecol. 2016, 72, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Martín-Vivaldi, M.; Soler, J.J.; Martínez-García, Á.; Arco, L.; Juárez-García-Pelayo, N.; Ruiz-Rodríguez, M.; Martínez-Bueno, M. Acquisition of uropygial gland microbiome by hoopoe nestlings. Microb. Ecol. 2018, 76, 285–297. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Wienemann, T.; Schmitt-Wagner, D.; Meuser, K.; Segelbacher, G.; Schink, B.; Brune, A.; Berthold, P. The bacterial microbiota in the ceca of Capercaillie (Tetrao urogallus) differs between wild and captive birds. Syst. Appl. Microbiol. 2011, 34, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Young, V.B.; Schmidt, T.M. Overview of the gastrointestinal microbiota. In GI Microbiota and Regulation of the Immune System; Huffnagle, G.B., Noverr, M.C., Eds.; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2008; pp. 29–40. ISBN 978-0-387-79989-6. [Google Scholar]
- Kreisinger, J.; Kropáčková, L.; Petrželková, A.; Adámková, M.; Tomášek, O.; Martin, J.-F.; Michálková, R.; Albrecht, T. Temporal stability and the effect of transgenerational transfer on fecal microbiota structure in a long distance migratory bird. Front. Microbiol. 2017, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Soler, J.J.; Martínez-García, Á.; Rodríguez-Ruano, S.M.; Martínez-Bueno, M.; Martín-Platero, A.M.; Peralta-Sánchez, J.M.; Martín-Vivaldi, M. Nestedness of hoopoes’ bacterial communities: Symbionts from the uropygial gland to the eggshell. Biol. J. Linn. Soc. 2016, 118, 763–773. [Google Scholar] [CrossRef]
- Kulkarni, S.; Heeb, P. Social and sexual behaviours aid transmission of bacteria in birds. Behav. Process. 2007, 74, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, Á.; Soler, J.J.; Rodríguez-Ruano, S.M.; Martínez-Bueno, M.; Martín-Platero, A.M.; Juárez-García, N.; Martín-Vivaldi, M. Preening as a vehicle for key bacteria in hoopoes. Microb. Ecol. 2015, 70, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Reichlin, T.S.; Hobson, K.A.; Wilgenburg, S.L.V.; Schaub, M.; Wassenaar, L.I.; Martín-Vivaldi, M.; Arlettaz, R.; Jenni, L. Conservation through connectivity: Can isotopic gradients in Africa reveal winter quarters of a migratory bird? Oecologia 2012, 171, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, Á.; Martín-Vivaldi, M.; Rodríguez-Ruano, S.M.; Peralta-Sánchez, J.M.; Valdivia, E.; Soler, J.J. Nest bacterial environment affects microbiome of hoopoe eggshells, but not that of the uropygial secretion. PLoS ONE 2016, 11, e0158158. [Google Scholar] [CrossRef] [PubMed]
- Dominianni, C.; Sinha, R.; Goedert, J.J.; Pei, Z.; Yang, L.; Hayes, R.B.; Ahn, J. Sex, body mass index, and dietary fiber intake influence the human gut microbiome. PLoS ONE 2015, 10, e0124599. [Google Scholar] [CrossRef] [PubMed]
- Org, E.; Mehrabian, M.; Parks, B.W.; Shipkova, P.; Liu, X.; Drake, T.A.; Lusis, A.J. Sex differences and hormonal effects on gut microbiota composition in mice. Gut Microbes 2016, 7, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Dohrmann, A.B.; Küting, M.; Jünemann, S.; Jaenicke, S.; Schlüter, A.; Tebbe, C.C. Importance of rare taxa for bacterial diversity in the rhizosphere of Bt− and conventional maize varieties. ISME J. 2013, 7, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Jian, W.; Dong, X. Transfer of Bifidobacterium inopinatum and Bifidobacterium denticolens to Scardovia inopinata gen. nov., comb. nov., and Parascardovia denticolens gen. nov., comb. nov., respectively. Int. J. Syst. Evol. Microbiol. 2002, 52, 809–812. [Google Scholar] [CrossRef] [PubMed]
- In, Y.-W.; Kim, J.-J.; Kim, H.-J.; Oh, S.-W. Antimicrobial activities of acetic acid, citric acid and lactic acid against Shigella species. J. Food Saf. 2013, 33, 79–85. [Google Scholar] [CrossRef]
- Batt, C.A. Encyclopedia of Food Microbiology; Batt, C.A., Ed.; Academic Press: Cambridge, MA, USA, 1999; ISBN 978-0-12-384733-1. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Chao1 Index | Faith’s Phylogenetic Diversity Index | |
|---|---|---|---|
| Breeding females | 47 | 47.9 ± 9.6 | 1.88 ± 0.34 |
| Nestlings | 18 | 43.0 ± 8.2 | 1.77 ± 0.28 |
| Breeding males | 3 | 59.3 ± 9.0 | 3.11 ± 0.15 |
| Non-nesting females | 2 | 50.0 ± 7.1 | 1.97 ± 0.39 |
| Phylum; Class | Breeding Females | Nestlings | Males | Non-Nesting Females | Total |
|---|---|---|---|---|---|
| Actinobacteria; Actinobacteria | 0.002 | 0.002 | 0.003 | 0.015 | 0.006 |
| Bacteroidetes; Bacteroidia | 0.046 | 0.029 | 0.042 | 0.002 | 0.030 |
| Firmicutes; Bacilli | 0.004 | 0.011 | 0.011 | 0.074 | 0.926 |
| Firmicutes; Clostridia | 0.931 | 0.945 | 0.896 | 0.832 | |
| Proteobacteria; Alphaproteobacteria | 0.004 | 0.000 | 0.003 | 0.068 | 0.038 |
| Proteobacteria; Betaproteobacteria | 0.000 | 0.000 | 0.007 | 0.000 | |
| Proteobacteria; Epsilonproteobacteria | 0.011 | 0.003 | 0.013 | 0.005 | |
| Proteobacteria; Gammaproteobacteria | 0.001 | 0.009 | 0.025 | 0.002 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Ruano, S.M.; Martín-Vivaldi, M.; Peralta-Sánchez, J.M.; García-Martín, A.B.; Martínez-García, Á.; Soler, J.J.; Valdivia, E.; Martínez-Bueno, M. Seasonal and Sexual Differences in the Microbiota of the Hoopoe Uropygial Secretion. Genes 2018, 9, 407. https://doi.org/10.3390/genes9080407
Rodríguez-Ruano SM, Martín-Vivaldi M, Peralta-Sánchez JM, García-Martín AB, Martínez-García Á, Soler JJ, Valdivia E, Martínez-Bueno M. Seasonal and Sexual Differences in the Microbiota of the Hoopoe Uropygial Secretion. Genes. 2018; 9(8):407. https://doi.org/10.3390/genes9080407
Chicago/Turabian StyleRodríguez-Ruano, Sonia M., Manuel Martín-Vivaldi, Juan M. Peralta-Sánchez, Ana B. García-Martín, Ángela Martínez-García, Juan J. Soler, Eva Valdivia, and Manuel Martínez-Bueno. 2018. "Seasonal and Sexual Differences in the Microbiota of the Hoopoe Uropygial Secretion" Genes 9, no. 8: 407. https://doi.org/10.3390/genes9080407
APA StyleRodríguez-Ruano, S. M., Martín-Vivaldi, M., Peralta-Sánchez, J. M., García-Martín, A. B., Martínez-García, Á., Soler, J. J., Valdivia, E., & Martínez-Bueno, M. (2018). Seasonal and Sexual Differences in the Microbiota of the Hoopoe Uropygial Secretion. Genes, 9(8), 407. https://doi.org/10.3390/genes9080407