The Colorful Sex Chromosomes of Teleost Fish


{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Sex Determination, Sex Chromosomes, and Sexually Antagonistic Genes
1.2. Pigmentation in Fish
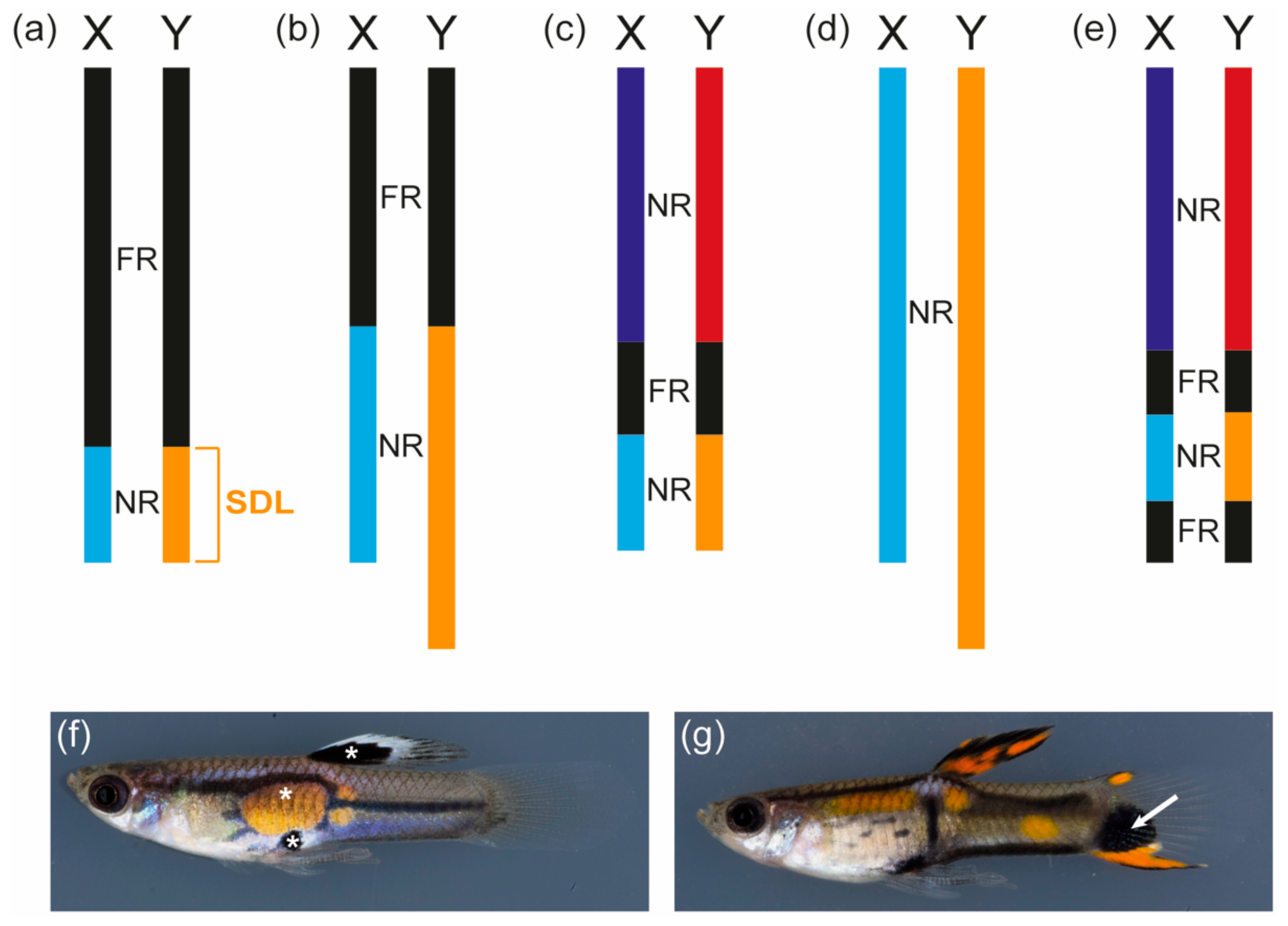
2. Guppy
3. Other Poecilia Species
4. Swordtails and Platyfish
5. Eastern Mosquitofish
6. Cichlids
7. Medaka
8. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Aida, T. On the inheritance of color in a fresh-water fish, Aplocheilus latipes Temmick and Schlegel, with special reference to sex-linked inheritance. Genetics 1921, 6, 554–573. [Google Scholar] [PubMed]
- Cott, H.B. Adaptive Coloration in Animals; Methuen & Co Ltd: London, UK, 1940. [Google Scholar]
- Goodrich, H.B.; Josephson, N.D.; Trinkaus, J.P.; Slate, J.M. The cellular expression and genetics of two new genes in Lebistes reticulatus. Genetics 1944, 29, 584–592. [Google Scholar] [PubMed]
- Owen, S.E. The reaction of fish to sex hormones. Endocrinology 1936, 20, 214–218. [Google Scholar] [CrossRef]
- Winge, Ö. One-sided masculine and sex-linked inheritance in Lebistes reticulatus. J. Genet. 1922, 12, 145–162. [Google Scholar] [CrossRef]
- Winge, Ö. The location of eighteen genes in Lebistes reticulatus. J. Genet. 1927, 18, 1–43. [Google Scholar] [CrossRef]
- Endler, J.A. Natural and sexual selection on color patterns in poeciliid fishes. Environ. Biol. Fishes 1983, 9, 173–190. [Google Scholar] [CrossRef]
- Leclercq, E.; Taylor, J.F.; Migaud, H. Morphological skin colour changes in teleosts. Fish Fish. 2010, 11, 159–193. [Google Scholar] [CrossRef]
- Maan, M.E.; Seehausen, O.; Soderberg, L.; Johnson, L.; Ripmeester, E.A.; Mrosso, H.D.; Taylor, M.I.; van Dooren, T.J.; van Alphen, J.J. Intraspecific sexual selection on a speciation trait, male coloration, in the Lake Victoria cichlid Pundamilia nyererei. Proc. Biol. Sci. 2004, 271, 2445–2452. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.B.; Ser, J.R.; Kocher, T.D. Sexual conflict resolved by invasion of a novel sex determiner in Lake Malawi cichlid fishes. Science 2009, 326, 998–1001. [Google Scholar] [CrossRef] [PubMed]
- Rowland, W.J. The relationships among nuptial coloration, aggression, and courtship of male three-spined sticklebacks, Gasterosteus aculeatus. Can. J. Zool. 1984, 62, 999–1004. [Google Scholar] [CrossRef]
- Endler, J.A. Natural selection on color patterns in Poecilia reticulata. Evolution 1980, 34, 76–91. [Google Scholar] [CrossRef] [PubMed]
- Endler, J.A. Multiple-trait coevolution and environmental gradients in guppies. Trends Ecol. Evol. 1995, 10, 22–29. [Google Scholar] [CrossRef]
- Owens, I.P.; Hartley, I.R. Sexual dimorphism in birds: Why are there so many different forms of dimorphism? Proc. R. Soc. Lond. B 1998, 265, 397–407. [Google Scholar] [CrossRef]
- Allender, C.J.; Seehausen, O.; Knight, M.E.; Turner, G.F.; Maclean, N. Divergent selection during speciation of Lake Malawi cichlid fishes inferred from parallel radiations in nuptial coloration. Proc. Natl. Acad. Sci. USA 2003, 100, 14074–14079. [Google Scholar] [CrossRef] [PubMed]
- Van Doorn, G.S. Intralocus sexual conflict. Ann. N. Y. Acad. Sci. 2009, 1168, 52–71. [Google Scholar] [CrossRef] [PubMed]
- Rice, W.R. Sex chromosomes and the evolution of sexual dimorphism. Evolution 1984, 38, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Chapman, T.; Arnqvist, G.; Bangham, J.; Rowe, L. Sexual conflict. Trends Ecol. Evol. 2003, 18, 41–47. [Google Scholar] [CrossRef]
- Harano, T.; Okada, K.; Nakayama, S.; Miyatake, T.; Hosken, D.J. Intralocus sexual conflict unresolved by sex-limited trait expression. Curr. Biol. 2010, 20, 2036–2039. [Google Scholar] [CrossRef] [PubMed]
- Kocher, T.D. Adaptive evolution and explosive speciation: The cichlid fish model. Nat. Rev. Genet. 2004, 5, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Lande, R.; Seehausen, O.; van Alphen, J.J. Mechanisms of rapid sympatric speciation by sex reversal and sexual selection in cichlid fish. Genetica 2001, 112–113, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.G. Sexual selection and speciation. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 79–102. [Google Scholar] [CrossRef]
- Charlesworth, D. The status of supergenes in the 21st century: Recombination suppression in Batesian mimicry and sex chromosomes and other complex adaptations. Evol. Appl. 2016, 9, 74–90. [Google Scholar] [CrossRef] [PubMed]
- Haskins, C.P.; Young, P.; Hewitt, R.E.; Haskins, E.F. Stabilised heterozygosis of supergenes mediating certain Y-linked colour patterns in populations of Lebistes reticulatus. Heredity 1970, 25, 575. [Google Scholar] [CrossRef]
- Peterson, M.P.; Rosvall, K.A.; Choi, J.H.; Ziegenfus, C.; Tang, H.; Colbourne, J.K.; Ketterson, E.D. Testosterone affects neural gene expression differently in male and female juncos: A role for hormones in mediating sexual dimorphism and conflict. PLoS ONE 2013, 8, e61784. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.P.; Lopez-Sepulcre, A.; Reznick, D.N. Predation-associated differences in sex linkage of wild guppy coloration. Evolution 2012, 66, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.P.; Lopez-Sepulcre, A.; Rumbo, D.; Reznick, D.N. Rapid changes in the sex linkage of male coloration in introduced guppy populations. Am. Nat. 2017, 189, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Haskins, C.; Haskins, E.; McLaughlin, J.; Hewitt, R. Polymorphism and population structure in Lebistes reticulatus, an ecological study. In Vertebrate Speciation; University of Texas Press: Austin, TX, USA, 1961; pp. 320–395. [Google Scholar]
- Rice, W.R. Evolution of the Y sex chromosome in animals. Bioscience 1996, 46, 331–343. [Google Scholar] [CrossRef]
- Rice, W.R. The accumulation of sexually antagonistic genes as a selective agent promoting the evolution of reduced recombination between primitive sex chromosomes. Evolution 1987, 41, 911–914. [Google Scholar] [CrossRef] [PubMed]
- Van Doorn, G.S.; Kirkpatrick, M. Turnover of sex chromosomes induced by sexual conflict. Nature 2007, 449, 909–912. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A. The evolution of dominance. Biol. Rev. 1931, 6, 345–368. [Google Scholar] [CrossRef]
- Handley, L.J.; Ceplitis, H.; Ellegren, H. Evolutionary strata on the chicken Z chromosome: Implications for sex chromosome evolution. Genetics 2004, 167, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Lahn, B.T.; Page, D.C. Four evolutionary strata on the human X chromosome. Science 1999, 286, 964–967. [Google Scholar] [CrossRef] [PubMed]
- Doncaster, L. Inheritance and sex in Abraxas grossulariata. Nature 1907, 76, 248. [Google Scholar] [CrossRef]
- Devlin, R.H.; Nagahama, Y. Sex determination and sex differentiation in fish: An overview of genetic, physiological, and environmental influences. Aquaculture 2002, 208, 191–364. [Google Scholar] [CrossRef]
- Volff, J.N.; Schartl, M. Variability of genetic sex determination in poeciliid fishes. Genetica 2001, 111, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Terai, Y.; Mizoiri, S.; Aibara, M.; Nishihara, H.; Watanabe, M.; Kuroiwa, A.; Hirai, H.; Hirai, Y.; Matsuda, Y.; et al. B chromosomes have a functional effect on female sex determination in Lake Victoria cichlid fishes. PLoS Genet. 2011, 7, e1002203. [Google Scholar] [CrossRef] [PubMed]
- Bachtrog, D.; Kirkpatrick, M.; Mank, J.E.; McDaniel, S.F.; Pires, J.C.; Rice, W.R.; Valenzuela, N. Are all sex chromosomes created equal? Trends Genet. 2011, 27, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Baroiller, J.F.; D’Cotta, H.; Saillant, E. Environmental effects on fish sex determination and differentiation. Sex. Dev. 2009, 3, 118–135. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, D.; Mank, J.E. The birds and the bees and the flowers and the trees: Lessons from genetic mapping of sex determination in plants and animals. Genetics 2010, 186, 9–31. [Google Scholar] [CrossRef] [PubMed]
- Godwin, J.; Luckenbach, J.A.; Borski, R.J. Ecology meets endocrinology: Environmental sex determination in fishes. Evol. Dev. 2003, 5, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Mank, J.E.; Promislow, D.E.L.; Avise, J.C. Evolution of alternative sex-determining mechanisms in teleost fishes. Biol. J. Linn. Soc. 2006, 87, 83–93. [Google Scholar] [CrossRef]
- Kelsh, R.N.; Harris, M.L.; Colanesi, S.; Erickson, C.A. Stripes and belly-spots—A review of pigment cell morphogenesis in vertebrates. Semin. Cell Dev. Biol. 2009, 20, 90–104. [Google Scholar] [CrossRef] [PubMed]
- Sauka-Spengler, T.; Bronner-Fraser, M. A gene regulatory network orchestrates neural crest formation. Nat. Rev. Mol. Cell. Biol. 2008, 9, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Schartl, M.; Larue, L.; Goda, M.; Bosenberg, M.W.; Hashimoto, H.; Kelsh, R.N. What is a vertebrate pigment cell? Pigment Cell Melanoma Res. 2016, 29, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Fujii, R. Cytophysiology of fish chromatophores. In International Review of Cytology; Elsevier: London, UK, 1993; Volume 143, pp. 191–255. [Google Scholar]
- Goda, M.; Fujiyoshi, Y.; Sugimoto, M.; Fujii, R. Novel dichromatic chromatophores in the integument of the mandarin fish Synchiropus splendidus. Biol. Bull. 2013, 224, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Goda, M.; Ohata, M.; Ikoma, H.; Fujiyoshi, Y.; Sugimoto, M.; Fujii, R. Integumental reddish-violet coloration owing to novel dichromatic chromatophores in the teleost fish, Pseudochromis diadema. Pigment Cell Melanoma Res. 2011, 24, 614–617. [Google Scholar] [CrossRef] [PubMed]
- Braasch, I.; Brunet, F.; Volff, J.N.; Schartl, M. Pigmentation pathway evolution after whole-genome duplication in fish. Genome Biol. Evol. 2009, 1, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Braasch, I.; Schartl, M.; Volff, J.N. Evolution of pigment synthesis pathways by gene and genome duplication in fish. BMC Evol. Biol. 2007, 7, 74. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.P.; Nüsslein-Volhard, C. Zebrafish stripes as a model for vertebrate colour pattern formation. Curr. Biol. 2015, 25, R81–R92. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, H.E. Genetics, development and evolution of adaptive pigmentation in vertebrates. Heredity 2006, 97, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Odenthal, J.; Rossnagel, K.; Haffter, P.; Kelsh, R.N.; Vogelsang, E.; Brand, M.; van Eeden, F.J.; Furutani-Seiki, M.; Granato, M.; Hammerschmidt, M.; et al. Mutations affecting xanthophore pigmentation in the zebrafish, Danio rerio. Development 1996, 123, 391–398. [Google Scholar] [PubMed]
- Kelsh, R.N.; Brand, M.; Jiang, Y.J.; Heisenberg, C.P.; Lin, S.; Haffter, P.; Odenthal, J.; Mullins, M.C.; van Eeden, F.J.; Furutani-Seiki, M.; et al. Zebrafish pigmentation mutations and the processes of neural crest development. Development 1996, 123, 369–389. [Google Scholar] [PubMed]
- Haffter, P.; Odenthal, J.; Mullins, M.C.; Lin, S.; Farrell, M.J.; Vogelsang, E.; Haas, F.; Brand, M.; van Eeden, F.J.; Furutani-Seiki, M.; et al. Mutations affecting pigmentation and shape of the adult zebrafish. Dev. Genes Evol. 1996, 206, 260–276. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Ferrer, Á.; Rodríguez-López, J.N.; García-Cánovas, F.; García-Carmona, F. Tyrosinase: A comprehensive review of its mechanism. Biochim. Biophys. Acta 1995, 1247, 1–11. [Google Scholar] [CrossRef]
- Del Marmol, V.; Beermann, F. Tyrosinase and related proteins in mammalian pigmentation. FEBS Lett. 1996, 381, 165–168. [Google Scholar] [CrossRef]
- Ben, J.; Lim, T.M.; Phang, V.P.; Chan, W.K. Cloning and tissue expression of 6-Pyruvoyl Tetrahydropterin synthase and Xanthine Dehydrogenase from Poecilia reticulata. Mar. Biotechnol. 2003, 5, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Parichy, D.M.; Rawls, J.F.; Pratt, S.J.; Whitfield, T.T.; Johnson, S.L. Zebrafish sparse corresponds to an orthologue of c-kit and is required for the morphogenesis of a subpopulation of melanocytes, but is not essential for hematopoiesis or primordial germ cell development. Development 1999, 126, 3425–3436. [Google Scholar] [PubMed]
- Kottler, V.A.; Fadeev, A.; Weigel, D.; Dreyer, C. Pigment pattern formation in the guppy, Poecilia reticulata, involves the Kita and Csf1ra receptor tyrosine kinases. Genetics 2013, 194, 631–646. [Google Scholar] [CrossRef] [PubMed]
- Rawls, J.F.; Johnson, S.L. Temporal and molecular separation of the kit receptor tyrosine kinase’s roles in zebrafish melanocyte migration and survival. Dev. Biol. 2003, 262, 152–161. [Google Scholar] [CrossRef]
- Parichy, D.M.; Ransom, D.G.; Paw, B.; Zon, L.I.; Johnson, S.L. An orthologue of the kit-related gene fms is required for development of neural crest-derived xanthophores and a subpopulation of adult melanocytes in the zebrafish, Danio rerio. Development 2000, 127, 3031–3044. [Google Scholar] [PubMed]
- Parichy, D.M.; Turner, J.M. Temporal and cellular requirements for Fms signaling during zebrafish adult pigment pattern development. Development 2003, 130, 817–833. [Google Scholar] [CrossRef] [PubMed]
- Frohnhöfer, H.G.; Krauss, J.; Maischein, H.-M.; Nüsslein-Volhard, C. Iridophores and their interactions with other chromatophores are required for stripe formation in zebrafish. Development 2013, 140, 2997–3007. [Google Scholar] [CrossRef] [PubMed]
- Inaba, M.; Yamanaka, H.; Kondo, S. Pigment pattern formation by contact-dependent depolarization. Science 2012, 335, 677. [Google Scholar] [CrossRef] [PubMed]
- Maderspacher, F.; Nüsslein-Volhard, C. Formation of the adult pigment pattern in zebrafish requires leopard and obelix dependent cell interactions. Development 2003, 130, 3447–3457. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Iwashita, M.; Ishii, M.; Kurachi, Y.; Kawakami, A.; Kondo, S.; Okada, N. Spot pattern of leopard Danio is caused by mutation in the zebrafish connexin41.8 gene. EMBO Rep. 2006, 7, 893–897. [Google Scholar] [CrossRef] [PubMed]
- Künstner, A.; Hoffmann, M.; Fraser, B.A.; Kottler, V.A.; Sharma, E.; Weigel, D.; Dreyer, C. The genome of the Trinidadian guppy, Poecilia reticulata, and variation in the Guanapo population. PLoS ONE 2016, 11, e0169087. [Google Scholar] [CrossRef] [PubMed]
- Dooley, C.M.; Schwarz, H.; Mueller, K.P.; Mongera, A.; Konantz, M.; Neuhauss, S.C.; Nüsslein-Volhard, C.; Geisler, R. Slc45a2 and V-ATPase are regulators of melanosomal pH homeostasis in zebrafish, providing a mechanism for human pigment evolution and disease. Pigment Cell Melanoma Res. 2013, 26, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Winge, Ö.; Ditlevsen, E. Colour inheritance and sex determination in Lebistes. Heredity 1947, 1, 65. [Google Scholar] [CrossRef]
- Charlesworth, B. The evolution of sex chromosomes. Science 1991, 251, 1030–1033. [Google Scholar] [CrossRef] [PubMed]
- Bergero, R.; Charlesworth, D. The evolution of restricted recombination in sex chromosomes. Trends Ecol. Evol. 2009, 24, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J. Racial investigations: IV. The genetic behaviour of a secondary sexual character. Cpts. Rds. Trav. Laborat. Carlsberg 1920, 14, 1–12. [Google Scholar] [CrossRef]
- Nanda, I.; Schories, S.; Tripathi, N.; Dreyer, C.; Haaf, T.; Schmid, M.; Schartl, M. Sex chromosome polymorphism in guppies. Chromosoma 2014, 123, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Magurran, A.E. Evolutionary Ecology: The Trinidadian Guppy; Oxford University Press on Demand: New York, NY, USA, 2005. [Google Scholar]
- Reznick, D.; Endler, J.A. The impact of predation on life history evolution in Trinidadian guppies (Poecilia reticulata). Evolution 1982, 36, 160–177. [Google Scholar] [PubMed]
- Reznick, D.N.; Bryga, H.A. Life-history evolution in guppies (Poecilia reticulata: Poeciliidae). V. Genetic basis of parallelism in life histories. Am. Nat. 1996, 147, 339–359. [Google Scholar] [CrossRef]
- Houde, A.E. Mate choice based upon naturally-occurring color-pattern variation in a guppy population. Evolution 1987, 41, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Olendorf, R.; Rodd, F.H.; Punzalan, D.; Houde, A.E.; Hurt, C.; Reznick, D.N.; Hughes, K.A. Frequency-dependent survival in natural guppy populations. Nature 2006, 441, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Rodd, F.H.; Hughes, K.A.; Grether, G.F.; Baril, C.T. A possible non-sexual origin of mate preference: Are male guppies mimicking fruit? Proc. Biol. Sci. 2002, 269, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.; Hoffmann, M.; Weigel, D.; Dreyer, C. Linkage analysis reveals the independent origin of poeciliid sex chromosomes and a case of atypical sex inheritance in the guppy (Poecilia reticulata). Genetics 2009, 182, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.; Hoffmann, M.; Willing, E.M.; Lanz, C.; Weigel, D.; Dreyer, C. Genetic linkage map of the guppy, Poecilia reticulata, and quantitative trait loci analysis of male size and colour variation. Proc. R. Soc. Lond. B 2009, 276, 2195–2208. [Google Scholar] [CrossRef] [PubMed]
- Lisachov, A.P.; Zadesenets, K.S.; Rubtsov, N.B.; Borodin, P.M. Sex chromosome synapsis and recombination in male guppies. Zebrafish 2015, 12, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Traut, W.; Winking, H. Meiotic chromosomes and stages of sex chromosome evolution in fish: Zebrafish, platyfish and guppy. Chromosome Res. 2001, 9, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Nanda, I.; Feichtinger, W.; Schmid, M.; Schröder, J.H.; Zischler, H.; Epplen, J.T. Simple repetitive sequences are associated with differentiation of the sex chromosomes in the guppy fish. J. Mol. Evol. 1990, 30, 456–462. [Google Scholar] [CrossRef]
- Winge, Ö.; Ditlevsen, E. A lethal gene in the Y-chromosome of Lebistes. CR Trav. Lab. Carlsberg. Ser. Physiol. 1938, 22, 203–210. [Google Scholar]
- Winge, Ö. The experimental alteration of sex chromosomes into autosomes and vice versa, as illustrated by Lebistes. CR Trav. Lab. Carlsberg. Ser. Physiol. 1934, 21, 1–49. [Google Scholar]
- Lindholm, A.; Breden, F. Sex chromosomes and sexual selection in poeciliid fishes. Am. Nat. 2002, 160 (Suppl. 6), S214–S224. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.; Hoffmann, M.; Dreyer, C. Natural variation of male ornamental traits of the guppy, Poecilia reticulata. Zebrafish 2008, 5, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.E.; Darolti, I.; Bloch, N.I.; Oostra, V.; Sandkam, B.; Buechel, S.D.; Kolm, N.; Breden, F.; Vicoso, B.; Mank, J.E. Convergent recombination suppression suggests role of sexual selection in guppy sex chromosome formation. Nat. Commun. 2017, 8, 14251. [Google Scholar] [CrossRef] [PubMed]
- Kottler, V.A.; Künstner, A.; Koch, I.; Flötenmeyer, M.; Langenecker, T.; Hoffmann, M.; Sharma, E.; Weigel, D.; Dreyer, C. Adenylate cyclase 5 is required for melanophore and male pattern development in the guppy (Poecilia reticulata). Pigment Cell Melanoma Res. 2015, 28, 545–558. [Google Scholar] [CrossRef] [PubMed]
- Sharma, E.; Künstner, A.; Fraser, B.A.; Zipprich, G.; Kottler, V.A.; Henz, S.R.; Weigel, D.; Dreyer, C. Transcriptome assemblies for studying sex-biased gene expression in the guppy, Poecilia reticulata. BMC Genomics 2014, 15, 400. [Google Scholar] [CrossRef] [PubMed]
- Hurtado-Gonzales, J.L.; Uy, J.A.C. Alternative mating strategies may favour the persistence of a genetically based colour polymorphism in a pentamorphic fish. Anim. Behav. 2009, 77, 1187–1194. [Google Scholar] [CrossRef]
- Lindholm, A.K.; Brooks, R.; Breden, F. Extreme polymorphism in a Y-linked sexually selected trait. Heredity 2004, 92, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Bourne, G.R.; Breden, F.; Allen, T.C. Females prefer carotenoid colored males as mates in the pentamorphic livebearing fish, Poecilia parae. Naturwissenschaften 2003, 90, 402–405. [Google Scholar] [CrossRef] [PubMed]
- Breden, F.; Ptacek, M.B.; Rashed, M.; Taphorn, D.; Figueiredo, C.A. Molecular phylogeny of the live-bearing fish genus Poecilia (Cyprinodontiformes: Poeciliidae). Mol. Phylogenet. Evol. 1999, 12, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, A.K.; Sandkam, B.; Pohl, K.; Breden, F. Poecilia picta, a close relative to the guppy, exhibits red male coloration polymorphism: A system for phylogenetic comparisons. PLoS ONE 2015, 10, e0142089. [Google Scholar] [CrossRef] [PubMed]
- Kallman, K. Genetic control of size at maturity in Xiphophorus. In Ecology and Evolution of Livebearing Fishes (Poeciliidae); Prentice-Hall: Englewood Cliffs, NJ, USA, 1989; pp. 163–184. [Google Scholar]
- Baer, C.F.; Dantzker, M.; Ryan, M.J. A test for preference of association in a color polymorphic poeciliid fish: Laboratory study. Environ. Biol. Fishes 1995, 43, 207–212. [Google Scholar] [CrossRef]
- Kingston, J.J.; Rosenthal, G.G.; Ryan, M.J. The role of sexual selection in maintaining a colour polymorphism in the pygmy swordtail, Xiphophorus pygmaeus. Anim. Behav. 2003, 65, 735–743. [Google Scholar] [CrossRef]
- Zimmerer, E.J.; Kallman, K.D. Genetic basis for alternative reproductive tactics in the pygmy swordtail, Xiphophorus nigrensis. Evolution 1989, 43, 1298–1307. [Google Scholar] [CrossRef] [PubMed]
- Lampert, K.P.; Schmidt, C.; Fischer, P.; Volff, J.N.; Hoffmann, C.; Muck, J.; Lohse, M.J.; Ryan, M.J.; Schartl, M. Determination of onset of sexual maturation and mating behavior by melanocortin receptor 4 polymorphisms. Curr. Biol. 2010, 20, 1729–1734. [Google Scholar] [CrossRef] [PubMed]
- Zimmerer, E.J.; Kallman, K.D. The inheritance of vertical barring (aggression and appeasement signals) in the pygmy swordtail, Xiphophorus nigrensis (Poeciliidae, Teleostei). Copeia 1988, 299–307. [Google Scholar] [CrossRef]
- Morris, M.R.; Mussel, M.; Ryan, M.J. Vertical bars on male Xiphophorus multilineatus: A signal that deters rival males and attracts females. Behav. Ecol. 1995, 6, 274–279. [Google Scholar] [CrossRef]
- Morris, M.R.; Nicoletto, P.F.; Hesselman, E. A polymorphism in female preference for a polymorphic male trait in the swordtail fish Xiphophorus cortezi. Anim. Behav. 2003, 65, 45–52. [Google Scholar] [CrossRef]
- Weis, S.; Schartl, M. The macromelanophore locus and the melanoma oncogene xmrk are separate genetic entities in the genome of Xiphophorus. Genetics 1998, 149, 1909–1920. [Google Scholar] [PubMed]
- Gordon, M. The hereditary basis for melanosis in hybrids of Mexican killifishes. Proc. Natl. Acad. Sci. USA 1931, 17, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M. A genetic concept for the origin of melanomas. Ann. N. Y. Acad. Sci. 1958, 71, 1213–1222. [Google Scholar] [CrossRef] [PubMed]
- Kallman, K.D. The platyfish, Xiphophorus maculatus. In Handbook of Genetics; Springer: Boston, MA, USA, 1975; pp. 81–132. [Google Scholar]
- Kallman, K.D. A new look at sex determination in poeciliid fishes. In Evolutionary Genetics of Fishes; Springer: Boston, MA, USA, 1984; pp. 95–171. [Google Scholar]
- Nanda, I.; Volff, J.N.; Weis, S.; Korting, C.; Froschauer, A.; Schmid, M.; Schartl, M. Amplification of a long terminal repeat-like element on the Y chromosome of the platyfish, Xiphophorus maculatus. Chromosoma 2000, 109, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Wittbrodt, J.; Adam, D.; Malitschek, B.; Maueler, W.; Raulf, F.; Telling, A.; Robertson, S.M.; Schartl, M. Novel putative receptor tyrosine kinase encoded by the melanoma-inducing Tu locus in Xiphophorus. Nature 1989, 341, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Kosswig, C. Über Bastarde der Teleostier Platyopoecilus und Xiphophorus. Z. Indukt. Abstamm Vererbungsl 1928, 47, 150–158. [Google Scholar]
- Schartl, M.; Hornung, U.; Gutbrod, H.; Volff, J.N.; Wittbrodt, J. Melanoma loss-of-function mutants in Xiphophorus caused by Xmrk-oncogene deletion and gene disruption by a transposable element. Genetics 1999, 153, 1385–1394. [Google Scholar] [PubMed]
- Gutbrod, H.; Schartl, M. Intragenic sex-chromosomal crossovers of Xmrk oncogene alleles affect pigment pattern formation and the severity of melanoma in Xiphophorus. Genetics 1999, 151, 773–783. [Google Scholar] [PubMed]
- Volff, J.N.; Korting, C.; Froschauer, A.; Zhou, Q.C.; Wilde, B.; Schultheis, C.; Selz, Y.; Sweeney, K.; Duschl, J.; Wichert, K.; et al. The Xmrk oncogene can escape nonfunctionalization in a highly unstable subtelomeric region of the genome of the fish Xiphophorus. Genomics 2003, 82, 470–479. [Google Scholar] [CrossRef]
- Basolo, A.L. Genetic linkage and color polymorphism in the southern platyfish (Xiphophorus maculatus): A model system for studies of color pattern evolution. Zebrafish 2006, 3, 65–83. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, A.A.; Morris, M.R. Mate choice for more melanin as a mechanism to maintain a functional oncogene. Proc. Natl. Acad. Sci. USA 2008, 105, 13503–13507. [Google Scholar] [CrossRef] [PubMed]
- Regan, J.D. Melanism in the poeciliid fish, Gambusia affinis (Baird and Girard). Am. Midl. Nat. 1961, 65, 139–143. [Google Scholar] [CrossRef]
- Angus, R. Inheritance of melanistic pigmentation in the Eastern mosquitofish. J. Heredity 1989, 80, 387–392. [Google Scholar] [CrossRef]
- Horth, L. Melanic body colour and aggressive mating behaviour are correlated traits in male mosquitofish (Gambusia holbrooki). Proc. Biol. Sci. 2003, 270, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Horth, L. Predation and the persistence of melanic male mosquitofish (Gambusia holbrooki). J. Evol. Biol. 2004, 17, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Horth, L. A sex-linked allele, autosomal modifiers and temperature-dependence appear to regulate melanism in male mosquitofish (Gambusia holbrooki). J. Exp. Biol. 2006, 209, 4938–4945. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Horth, L.; Travis, J. Frequency-dependent numerical dynamics in mosquitofish. Proc. Biol. Sci. 2002, 269, 2239–2247. [Google Scholar] [CrossRef] [PubMed]
- Horth, L.; Binckley, C.; Wilk, R.; Reddy, P.; Reddy, A. Color, body size, and genitalia size are correlated traits in eastern mosquitofish (Gambusia holbrooki). Copeia 2010, 196–202. [Google Scholar] [CrossRef]
- Martin, R.G. Density dependent aggressive advantage in melanistic male mosquitofish Gambusia affinis holbrooki (Girard). FL Sci. 1977, 40, 393–400. [Google Scholar]
- Bisazza, A.; Pilastro, A. Variation of female preference for male coloration in the eastern mosquitofish Gambusia holbrooki. Behav. Genet. 2000, 30, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Black, D.A.; Howell, W.M. The North American mosquitofish, Gambusia affinis: A unique case in sex chromosome evolution. Copeia 1979, 1979, 509–513. [Google Scholar] [CrossRef]
- Snelson, F.F., Jr.; Smith, R.E.; Bolt, M.R. A melanistic female mosquitofish, Gambusia affinis holbrooki. Am. Midl. Nat. 1986, 115, 413–415. [Google Scholar] [CrossRef]
- Salzburger, W. The interaction of sexually and naturally selected traits in the adaptive radiations of cichlid fishes. Mol. Ecol. 2009, 18, 169–185. [Google Scholar] [CrossRef] [PubMed]
- Maan, M.E.; Sefc, K.M. Colour variation in cichlid fish: Developmental mechanisms, selective pressures and evolutionary consequences. Semin. Cell Dev. Biol. 2013, 24, 516–528. [Google Scholar] [CrossRef] [PubMed]
- Parnell, N.F.; Streelman, J.T. Genetic interactions controlling sex and color establish the potential for sexual conflict in Lake Malawi cichlid fishes. Heredity 2013, 110, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Ser, J.R.; Roberts, R.B.; Kocher, T.D. Multiple interacting loci control sex determination in Lake Malawi cichlid fish. Evolution 2010, 64, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Streelman, J.T.; Albertson, R.C.; Kocher, T.D. Genome mapping of the orange blotch colour pattern in cichlid fishes. Mol. Ecol. 2003, 12, 2465–2471. [Google Scholar] [CrossRef] [PubMed]
- Seehausen, O.; van Alphen, J.J.M.; Lande, R. Color polymorphism and sex ratio distortion in a cichlid fish as an incipient stage in sympatric speciation by sexual selection. Ecol. Lett. 1999, 2, 367–378. [Google Scholar] [CrossRef]
- Takahashi, T.; Sota, T.; Hori, M. Genetic basis of male colour dimorphism in a Lake Tanganyika cichlid fish. Mol. Ecol. 2013, 22, 3049–3060. [Google Scholar] [CrossRef] [PubMed]
- Salzburger, W.; Braasch, I.; Meyer, A. Adaptive sequence evolution in a color gene involved in the formation of the characteristic egg-dummies of male haplochromine cichlid fishes. BMC Biol. 2007, 5, 51. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.E.; Braasch, I.; Boileau, N.; Meyer, B.S.; Sauteur, L.; Bohne, A.; Belting, H.G.; Affolter, M.; Salzburger, W. The evolution of cichlid fish egg-spots is linked with a cis-regulatory change. Nat. Commun. 2014, 5, 5149. [Google Scholar] [CrossRef] [PubMed]
- Gunter, H.M.; Clabaut, C.; Salzburger, W.; Meyer, A. Identification and characterization of gene expression involved in the coloration of cichlid fish using microarray and qRT-PCR approaches. J. Mol. Evol. 2011, 72, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Hornung, U.; Nanda, I.; Imai, S.; Sasaki, T.; Shimizu, A.; Asakawa, S.; Hori, H.; Schmid, M.; Shimizu, N.; et al. Genomic organization of the sex-determining and adjacent regions of the sex chromosomes of medaka. Genome Res. 2006, 16, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Nanda, I.; Kondo, M.; Hornung, U.; Asakawa, S.; Winkler, C.; Shimizu, A.; Shan, Z.; Haaf, T.; Shimizu, N.; Shima, A.; et al. A duplicated copy of DMRT1 in the sex-determining region of the Y chromosome of the medaka, Oryzias latipes. Proc. Natl. Acad. Sci. USA 2002, 99, 11778–11783. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; Nagahama, Y.; Shinomiya, A.; Sato, T.; Matsuda, C.; Kobayashi, T.; Morrey, C.E.; Shibata, N.; Asakawa, S.; Shimizu, N.; et al. DMY is a Y-specific DM-domain gene required for male development in the medaka fish. Nature 2002, 417, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Nanda, I.; Hornung, U.; Schmid, M.; Schartl, M. Evolutionary origin of the medaka Y chromosome. Curr. Biol. 2004, 14, 1664–1669. [Google Scholar] [CrossRef] [PubMed]
- Wada, H.; Shimada, A.; Fukamachi, S.; Naruse, K.; Shima, A. Sex-linked inheritance of the lf locus in the medaka fish (Oryzias latipes). Zool. Sci. 1998, 15, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; Matsuda, C.; Hamaguchi, S.; Sakaizumi, M. Identification of the sex chromosomes of the medaka, Oryzias latipes, by fluorescence in situ hybridization. Cytogenet. Cell Genet. 1998, 82, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Schartl, M.; Schmid, M.; Nanda, I. Dynamics of vertebrate sex chromosome evolution: From equal size to giants and dwarfs. Chromosoma 2016, 125, 553–571. [Google Scholar] [CrossRef] [PubMed]
- Sedlacek, O.; Baciakova, B.; Kratochvil, L. Evolution of body colouration in killifishes (Cyprinodontiformes: Aplocheilidae, Nothobranchiidae, Rivulidae): Is male ornamentation constrained by intersexual genetic correlation? Zool. Anz. 2014, 253, 207–215. [Google Scholar] [CrossRef]
- Kunte, K.; Zhang, W.; Tenger-Trolander, A.; Palmer, D.H.; Martin, A.; Reed, R.D.; Mullen, S.P.; Kronforst, M.R. Doublesex is a mimicry supergene. Nature 2014, 507, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Küpper, C.; Stocks, M.; Risse, J.E.; Dos Remedios, N.; Farrell, L.L.; McRae, S.B.; Morgan, T.C.; Karlionova, N.; Pinchuk, P.; Verkuil, Y.I.; et al. A supergene determines highly divergent male reproductive morphs in the ruff. Nat. Genet. 2016, 48, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Lamichhaney, S.; Fan, G.; Widemo, F.; Gunnarsson, U.; Thalmann, D.S.; Hoeppner, M.P.; Kerje, S.; Gustafson, U.; Shi, C.; Zhang, H.; et al. Structural genomic changes underlie alternative reproductive strategies in the ruff (Philomachus pugnax). Nat. Genet. 2016, 48, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Joron, M.; Frezal, L.; Jones, R.T.; Chamberlain, N.L.; Lee, S.F.; Haag, C.R.; Whibley, A.; Becuwe, M.; Baxter, S.W.; Ferguson, L.; et al. Chromosomal rearrangements maintain a polymorphic supergene controlling butterfly mimicry. Nature 2011, 477, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Le Poul, Y.; Whibley, A.; Chouteau, M.; Prunier, F.; Llaurens, V.; Joron, M. Evolution of dominance mechanisms at a butterfly mimicry supergene. Nat. Commun. 2014, 5, 5644. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kottler, V.A.; Schartl, M. The Colorful Sex Chromosomes of Teleost Fish. Genes 2018, 9, 233. https://doi.org/10.3390/genes9050233
Kottler VA, Schartl M. The Colorful Sex Chromosomes of Teleost Fish. Genes. 2018; 9(5):233. https://doi.org/10.3390/genes9050233
Chicago/Turabian StyleKottler, Verena A., and Manfred Schartl. 2018. "The Colorful Sex Chromosomes of Teleost Fish" Genes 9, no. 5: 233. https://doi.org/10.3390/genes9050233
APA StyleKottler, V. A., & Schartl, M. (2018). The Colorful Sex Chromosomes of Teleost Fish. Genes, 9(5), 233. https://doi.org/10.3390/genes9050233