Shared and Species-Specific Patterns of Nascent Y Chromosome Evolution in Two Guppy Species


Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Sequencing
2.2. Quality Trimming and Filtering
2.3. k-mer Counting
2.4. Identifying Y-Sequence
2.5. Y-Sequence Characterization
2.6. Origins of Y Genes
2.7. K-mer Composition Comparisons
3. Results
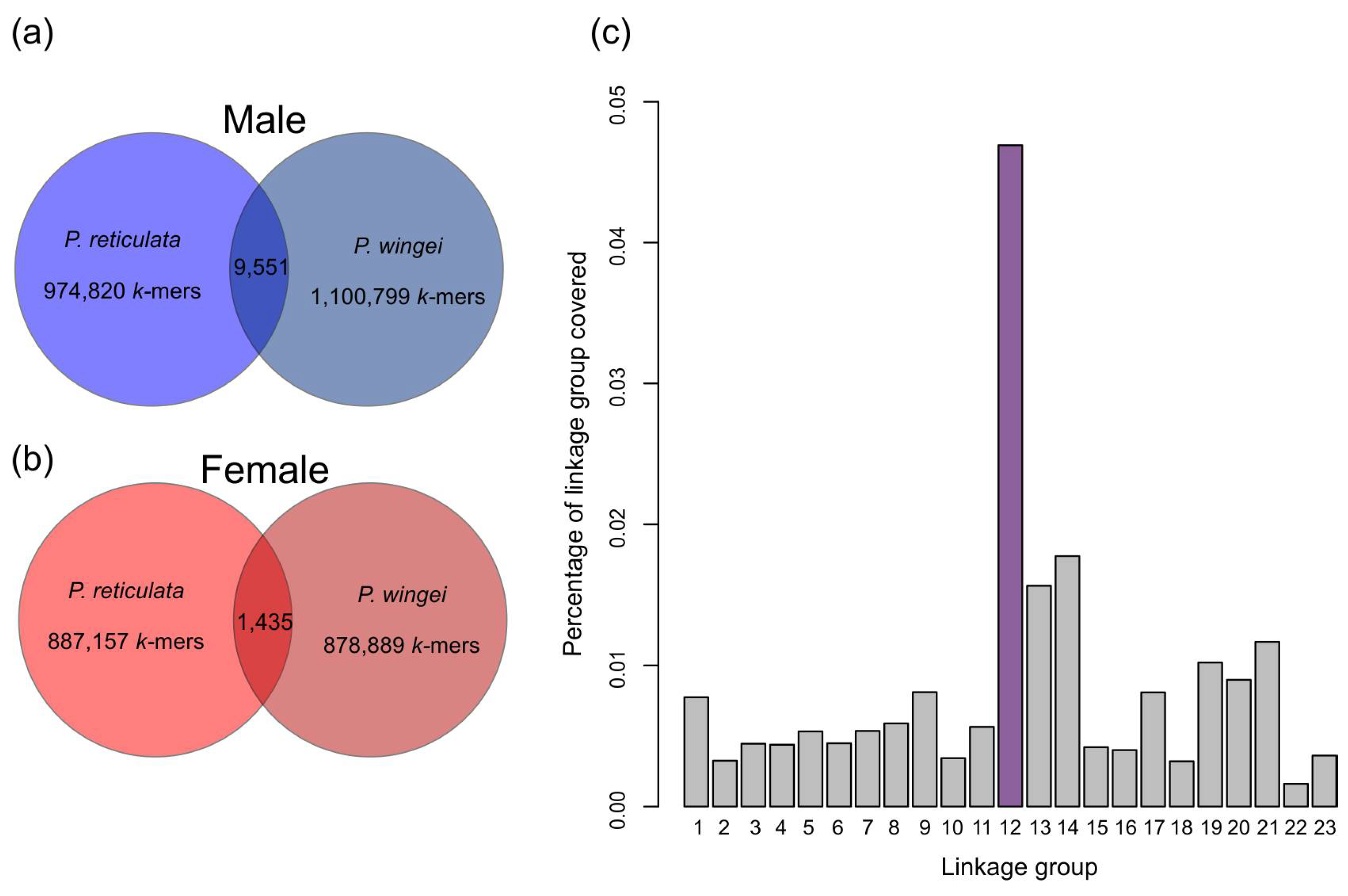
3.1. Y-mer Identification
3.2. Characterization of the Y Sequence
3.3. Origins of Y Genes
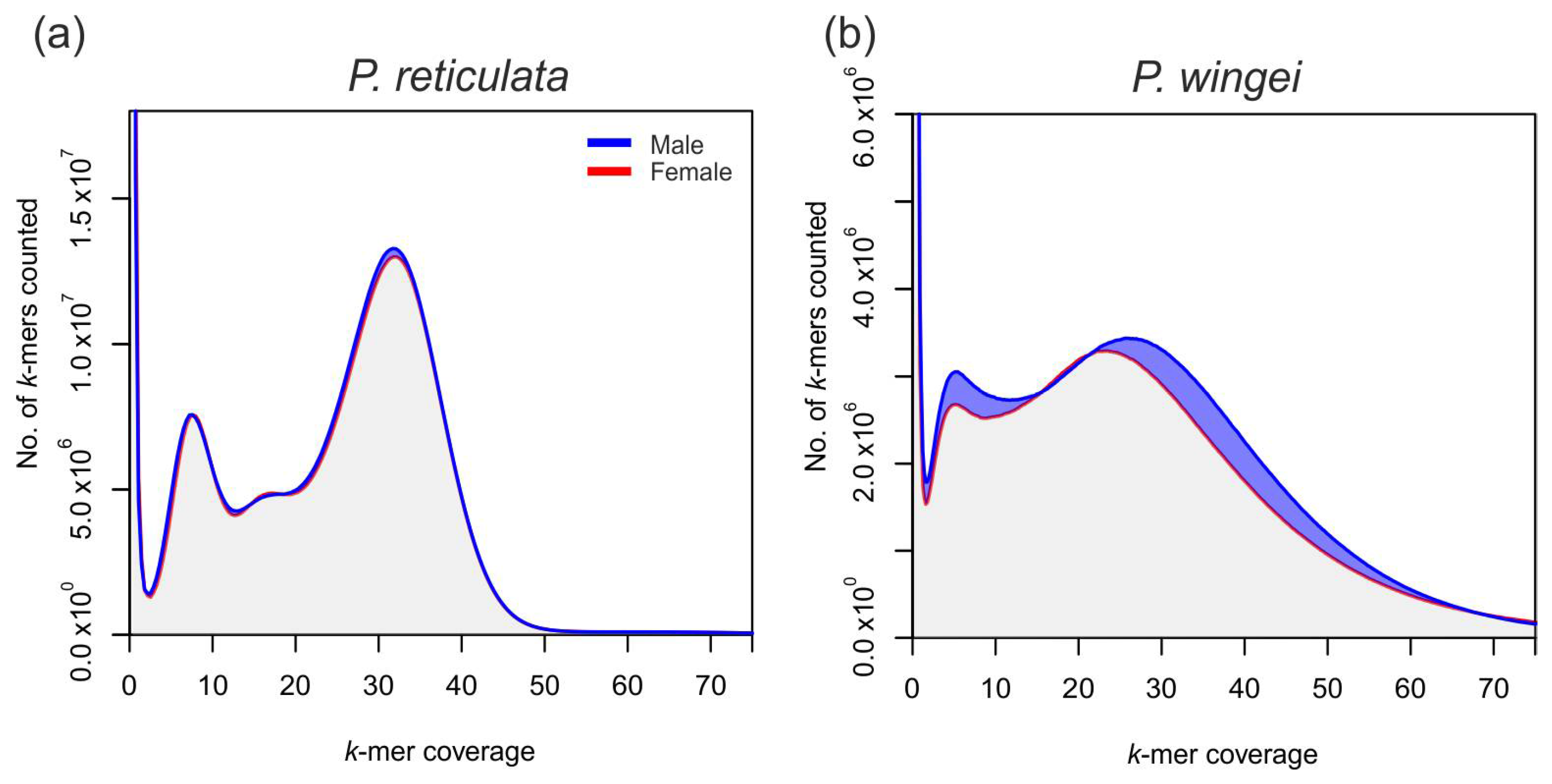
3.4. Poecilia reticulata k-mer Composition
3.5. Poecilia wingei k-mer Composition
4. Discussion
4.1. Young Sex Chromosomes and Mixed Sources of Genic Content
4.2. Non-Linear Degeneration of the Guppy Y Chromosome
4.3. Possible Pigmentation Loci Candidates
4.4. Possible Sex Determination Candidates
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.-L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R.; et al. Sex determination: Why so many ways of doing it? PLoS Biol. 2014, 12, e1001899. [Google Scholar] [CrossRef] [PubMed]
- Beukeboom, L.W.; Perrin, N. The Evolution of Sex Determination; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Rice, W.R. The accumulation of sexually antagonistic genes as a selective agent promoting the evolution of reduced recombination between primitive sex chromosomes. Evolution 1987, 41, 911–914. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.E.; Dean, R.; Zimmer, F.; Mank, J.E. How to make a sex chromosome. Nat. Commun. 2016, 7, 12087. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, B. Evolution of sex chromosomes. Science 1991, 251, 1030–1033. [Google Scholar] [CrossRef] [PubMed]
- Rice, W.R. Evolution of the Y sex in animals: Y chromosomes evolve through the degeneration of autosomes. Bioscience 1996, 46, 331–343. [Google Scholar] [CrossRef]
- Koerich, L.B.; Wang, X.; Clark, A.G.; Carvalho, A.B. Low conservation of gene content in the Drosophila Y. chromosome. Nature 2008, 456, 949–951. [Google Scholar] [CrossRef] [PubMed]
- Yoshido, A.; Marec, F.; Sahara, K. The fate of W chromosomes in hybrids between wild silkmoths, Samia cynthia ssp.: No role in sex determination and reproduction. Heredity 2016, 116, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Perrin, N. Sex reversal: A fountain of youth for sex chromosomes? Evolution 2009, 63, 3043–3049. [Google Scholar] [CrossRef] [PubMed]
- Schartl, M.; Schmid, M.; Nanda, I. Dynamics of vertebrate sex chromosome evolution: From equal size to giants and dwarfs. Chromosoma 2016, 125, 553–571. [Google Scholar] [CrossRef] [PubMed]
- Bachtrog, D. Y-chromosome evolution: Emerging insights into processes of Y-chromosome degeneration. Nat. Rev. Genet. 2013, 14, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Skaletsky, H.; Kuroda-Kawaguchi, T.; Minx, P.J.; Cordum, H.S.; Hillier, L.; Brown, L.G.; Repping, S.; Pyntikova, T.; Ali, J.; Bieri, T.; et al. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 2003, 423, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Soh, Y.Q.S.; Alföldi, J.; Pyntikova, T.; Brown, L.G.; Graves, T.; Minx, P.J.; Fulton, R.S.; Kremitzki, C.; Koutseva, N.; Mueller, J.L.; et al. Sequencing the mouse Y chromosome reveals convergent gene acquisition and amplification on both sex chromosomes. Cell 2014, 159, 800–813. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, B.; Crow, J.F. Model for evolution of Y chromosomes and dosage compensation. Proc. Natl. Acad. Sci. USA 1978, 75, 5618–5622. [Google Scholar] [CrossRef] [PubMed]
- Tomaszkiewicz, M.; Rangavittal, S.; Cechova, M.; Sanchez, C.; Fescemyer, H.W.; Harris, R.; Ye, D.; Brien, C.M.O.; Chikhi, R.; Ryder, O.A.; et al. A time- and cost-effective strategy to sequence mammalian Y chromosomes: An application to the de novo assembly of gorilla Y. Genome Res. 2016, 26, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.F.; Skaletsky, H.; Pyntikova, T.; Graves, T.A.; Van Daalen, S.K.M.; Minx, P.J.; Fulton, R.S.; McGrath, S.D.; Locke, D.P.; Friedman, C.; et al. Chimpanzee and human Y chromosomes are remarkably divergent in structure and gene content. Nature 2010, 463, 536–539. [Google Scholar] [CrossRef] [PubMed]
- Kichigin, I.G.; Giovannotti, M.; Makunin, A.I.; Ng, B.L.; Kabilov, M.R.; Tupikin, A.E.; Barucchi, V.C.; Splendiani, A.; Ruggeri, P.; Rens, W.; et al. Evolutionary dynamics of Anolis sex chromosomes revealed by sequencing of flow sorting-derived microchromosome-specific DNA. Mol. Genet. Genom. 2016, 291, 1955–1966. [Google Scholar] [CrossRef] [PubMed]
- Traut, W.; Vogel, H.; Glöckner, G.; Hartmann, E.; Heckel, D.G. High-throughput sequencing of a single chromosome: A moth W chromosome. Chromosom. Res. 2013, 21, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Na, J.; Yu, Q.; Gschwend, A.R.; Han, J.; Zeng, F.; Aryal, R.; VanBuren, R.; Murray, J.E.; Zhang, W.; et al. Sequencing papaya X and Yh chromosomes reveals molecular basis of incipient sex chromosome evolution. Proc. Natl. Acad. Sci. USA 2012, 109, 13710–13715. [Google Scholar] [CrossRef] [PubMed]
- Beaudry, F.E.G.; Barrett, S.C.H.; Wright, S.I. Genomic loss and silencing on the Y chromosomes of Rumex. Genome Biol. Evol. 2017, 9, 3345–3355. [Google Scholar] [CrossRef] [PubMed]
- Crowson, D.; Barrett, S.C.H.; Wright, S.I. Purifying and positive selection influence patterns of gene loss and gene expression in the evolution of a plant sex chromosome system. Mol. Biol. Evol. 2018, 34, 1140–1154. [Google Scholar] [CrossRef] [PubMed]
- Pucholt, P.; Wright, A.E.; Conze, L.L.; Mank, J.E.; Berlin, S. Recent sex chromosome divergence despite ancient dioecy in the willow Salix viminalis. Mol. Biol. Evol. 2017, 34, 1991–2001. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, A.; Filipović, I.; Fansiri, T.; Hoffmann, A.A.; Cheng, C.; Kirkpatrick, M.; Rašić, G.; Lambrechts, L. Extensive genetic differentiation between homomorphic sex chromosomes in the mosquito vector, Aedes aegypti. Genome Biol. Evol. 2017, 9, 2322–2335. [Google Scholar] [CrossRef] [PubMed]
- Lambert, M.R.; Skelly, D.K.; Ezaz, T. Sex-linked markers in the North American green frog (Rana clamitans) developed using DArTseq provide early insight into sex chromosome evolution. BMC Genom. 2016, 17, 844. [Google Scholar] [CrossRef] [PubMed]
- Conte, M.A.; Gammerdinger, W.J.; Bartie, K.L.; Penman, D.J.; Kocher, T.D. A high quality assembly of the Nile Tilapia (Oreochromis niloticus) genome reveals the structure of two sex determination regions. BMC Genom. 2017, 18, 341. [Google Scholar] [CrossRef] [PubMed]
- Reichwald, K.; Petzold, A.; Koch, P.; Downie, B.R.; Hartmann, N.; Pietsch, S.; Baumgart, M.; Chalopin, D.; Felder, M.; Bens, M.; et al. Insights into sex chromosome evolution and aging from the genome of a short-lived fish. Cell 2015, 163, 1527–1538. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Huang, R.; Chen, L.; Xiong, L.; He, L.; Li, Y.; Liao, L.; Zhu, Z.; Wang, Y. Computational identification of Y-linked markers and genes in the grass carp genome by using a pool-and-sequence method. Sci. Rep. 2017, 7, 8213. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Pang, M.; Yu, X.; Zhou, Y.; Tong, J.; Fu, B. Sex-specific markers developed by next-generation sequencing confirmed an XX/XY sex determination system in bighead carp (Hypophthalmichehys nobilis) and silver carp (Hypophthalmichthys molitrix). DNA Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ajimura, M.; Chen, Z.; Liu, J.; Chen, E.; Guo, H.; Tadapatri, V.; Reddy, C.G.; Zhang, J.; Kishino, H.; et al. A new approach for comprehensively describing heterogametic sex chromosomes. DNA Res. 2018, dsx054. [Google Scholar] [CrossRef] [PubMed]
- Meredith, R.W.; Pires, M.N.; Reznick, D.N.; Springer, M.S. Molecular phylogenetic relationships and the evolution of the placenta in Poecilia (Micropoecilia) (Poeciliidae: Cyprinodontiformes). Mol. Phylogenet. Evol. 2010, 55, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Nanda, I.; Schories, S.; Tripathi, N.; Dreyer, C.; Haaf, T.; Schmid, M.; Schartl, M. Sex chromosome polymorphism in guppies. Chromosoma 2014, 123, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.E.; Darolti, I.; Bloch, N.I.; Oostra, V.; Sandkam, B.; Buechel, S.D.; Kolm, N.; Breden, F.; Vicoso, B.; Mank, J.E. Convergent recombination suppression suggests role of sexual selection in guppy sex chromosome formation. Nat. Commun. 2017, 8, 14251. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, A.; Breden, F. Sex chromosomes and sexual selection in Poeciliid fishes. Am. Nat. 2002, 160, S214–S224. [Google Scholar] [CrossRef] [PubMed]
- Winge, Ö. The location of eighteen genes in Lebistes reticulatus. J. Genet. 1927, 18, 1–43. [Google Scholar] [CrossRef]
- Rangavittal, S.; Harris, R.S.; Cechova, M.; Tomaszkiewicz, M.; Chikhi, R.; Makova, K.D.; Medvedev, P. RecoverY: K-mer-based read classification for Y-chromosome-specific sequencing and assembly. Bioinformatics 2017, 34, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.; Clark, A. Efficient identification of Y chromosome sequences in the human and Drosophila genomes. Genome Res. 2013, 23, 1894–1907. [Google Scholar] [CrossRef] [PubMed]
- Akagi, T.; Henry, I.M.; Tao, R.; Comai, L. A Y-chromosome-encoded small RNA acts as a sex determinant in persimmons. Science 2014, 346, 646–650. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.F.; Mathew, L.S.; Ahmed, I.; Al-azwani, I.K.; Rivera, D.; Mohamoud, Y.A.; Clark, A.G.; Malek, J.A. Genus-wide sequencing supports a two-locus model for sex-determination in Phoenix. bioRxiv 2018, 245514. [Google Scholar] [CrossRef]
- Tomaszkiewicz, M.; Medvedev, P.; Makova, K.D. Y and W chromosome assemblies: Approaches and discoveries. Trends Genet. 2017, 33, 266–282. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.; Hoffmann, M.; Willing, E.-M.; Lanz, C.; Weigel, D.; Dreyer, C. Genetic linkage map of the guppy, Poecilia reticulata, and quantitative trait loci analysis of male size and colour variation. Proc. Biol. Sci. 2009, 276, 2195–2208. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.; Hoffmann, M.; Weigel, D.; Dreyer, C. Linkage analysis reveals the independent origin of poeciliid sex chromosomes and a case of atypical sex inheritance in the guppy (Poecilia reticulata). Genetics 2009, 182, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Florea, L.; Langmead, B. Lighter: Fast and memory-efficient error correction without counting. Genome Biol. 2014, 15, 509. [Google Scholar] [CrossRef] [PubMed]
- Marçais, G.; Kingsford, C. A fast, lock-free approach for efficient parallel counting of occurrences of k-mers. Bioinformatics 2011, 27, 764–770. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.T.; Wong, K.; Jackman, S.D.; Schein, J.E.; Jones, S.J.M. ABySS: A parallel assembler for short read sequence data. Genome Res. 2009, 19, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Altschup, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Ku, A.; Hoffmann, M.; Fraser, B.A.; Kottler, V.A.; Sharma, E.; Weigel, D.; Dreyer, C. The genome of the Trinidadian guppy, Poecilia reticulata, and variation in the Guanapo population. PLoS ONE 2016, 11, e0169087. [Google Scholar] [CrossRef]
- Carvalho, A.B.; Vicoso, B.; Russo, C.A.M.; Swenor, B.; Clark, A.G. Birth of a new gene on the Y chromosome of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2015, 112, 12450–12455. [Google Scholar] [CrossRef] [PubMed]
- Bull, J.J. Evolution of Sex Determining Mechanisms; The Benjamin/Cummings Publishing Company: Menlo Park, CA, USA, 1983; ISBN 0805304002. [Google Scholar]
- Hall, A.B.; Qi, Y.; Timoshevskiy, V.; Sharakhova, M.V.; Sharakhov, I.V.; Tu, Z. Six novel Y chromosome genes in Anopheles mosquitoes discovered by independently sequencing males and females. BMC Genom. 2013, 14, 273. [Google Scholar] [CrossRef] [PubMed]
- Meisel, R.; Gonzales, C.A.; Luu, H. The house fly Y chromosome is young and undifferentiated from its ancient X chromosome partner. Genome Res. 2017, 27, 1417–1426. [Google Scholar] [CrossRef] [PubMed]
- VanKuren, N.W.; Long, M. Gene duplicates resolving sexual conflict rapidly evolved essential gametogenesis functions. Nat. Ecol. Evol. 2018, 2, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Nanda, I.; Kondo, M.; Hornung, U.; Asakawa, S.; Winkler, C.; Shimizu, A.; Shan, Z.; Haaf, T.; Shimizu, N.; Shima, A.; et al. A duplicated copy of DMRT1 in the sex-determining region of the Y chromosome of the medaka, Oryzias latipes. Proc. Natl. Acad. Sci. USA 2002, 99, 11778–11783. [Google Scholar] [CrossRef] [PubMed]
- Hattori, R.S.; Murai, Y.; Oura, M.; Masuda, S.; Majhi, S.K.; Sakamoto, T.; Fernandino, J.I.; Somoza, G.M.; Yokota, M.; Strussmann, C.A. A Y-linked anti-Mullerian hormone duplication takes over a critical role in sex determination. Proc. Natl. Acad. Sci. USA 2012, 109, 2955–2959. [Google Scholar] [CrossRef] [PubMed]
- Gallach, M.; Betrán, E. Intralocus sexual conflict resolved through gene duplication. Trends Ecol. Evol. 2011, 26, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Grossen, C.; Neuenschwander, S.; Perrin, N. The evolution of XY recombination: Sexually antagonistic selection versus deleterious mutation load. Evolution 2012, 66, 3155–3166. [Google Scholar] [CrossRef] [PubMed]
- Parichy, D.M.; Rawls, J.F.; Pratt, S.J.; Whitfield, T.T.; Johnson, S.L. Zebrafish sparse corresponds to an orthologue of c-kit and is required for the morphogenesis of a subpopulation of melanocytes, but is not essential for hematopoiesis or primordial germ cell development. Development 1999, 126, 3425–3436. [Google Scholar] [PubMed]
- Kottler, V.A.; Fadeev, A.; Weige, D.; Dreyer, C. Pigment pattern formation in the guppy, Poecilia reticulata, involves the Kita and Csf1ra receptor tyrosine kinases. Genetics 2013, 194, 631–646. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A. The evolution of dominance. Biol. Rev. 1931, 6, 345–368. [Google Scholar] [CrossRef]
- Hiort, O. The differential role of androgens in early human sex development. BMC Med. 2013, 11, 152. [Google Scholar] [CrossRef] [PubMed]
- Oike, A.; Kodama, M.; Yasumasu, S.; Yamamoto, T.; Nakamura, Y.; Ito, E.; Nakamura, M. Participation of androgen and its receptor in sex determination of an amphibian species. PLoS ONE 2017, 12, e0178067. [Google Scholar] [CrossRef] [PubMed]
- Uno, Y.; Hosaka, S.; Yamazaki, H. Identification and analysis of CYP27A, CYP17A1, CYP20A1, CYP27A1 and CYP51A1 in Cynomolgus Macaques. J. Vet. Med. Sci. 2014, 76, 1647–1650. [Google Scholar] [CrossRef] [PubMed]
- Mast, N.; Lin, J.B.; Pikuleva, I.A. Marketed drugs can inhibit cytochrome p450 27A1, a potential new target for breast cancer adjuvant therapy. Mol. Pharmacol. 2015, 88, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Kohno, S.; Bernhard, M.C.; Katsu, Y.; Zhu, J.; Bryan, T.A.; Doheny, B.M.; Iguchi, T.; Guillette, L.J. Estrogen receptor 1 (ESR1; ERα), not ESR2 (ERβ), modulates estrogen-induced sex reversal in the American alligator, a species with temperature-dependent sex determination. Endocrinology 2015, 156, 1887–1899. [Google Scholar] [CrossRef] [PubMed]
- Maruo, K.; Suda, M.; Yokoyama, S.; Oshima, Y.; Nakamura, M. Steroidogenic gene expression during sex determination in the frog Rana rugosa. Gen. Comp. Endocrinol. 2008, 158, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M. Is a sex-determining gene(s) necessary for sex-determination in amphibians? Steroid hormones may be the key factor. Sex. Dev. 2012, 7, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.-F.; Li, J.-T.; Sun, X.-W. Complete mitochondrial genome of the guppy (Poecilia reticulata). Mitochondrial DNA 2016, 27, 228–229. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene | Genome Position |
|---|---|
| DNAJC1 2 | Unpl. Scaff. |
| CAMTA2 4 | Unpl. Scaff. |
| CDK5R1-like 1 | LG8 |
| CECR5-like 1 | Multi. hits |
| CNRIP1-like 3 | LG1 |
| CYP27A 6 | LG12 |
| jockey/pol-like 2 | Multi. hits |
| ELF2-like 5 | Multi. hits |
| EPHB4-like 2 | Unpl. Scaff. |
| gastrula zinc finger protein XlCGF17.1-like 3 | Multi. hits |
| GSN 1,5 | Unpl. Scaff. |
| KCNV2 2 | LG12 |
| LRRC39 1 | LG4 |
| MDH2 1 | LG16 |
| MED13LL 1 | LG12 |
| MLPH-like 1 | LG2 |
| NFIL3 1 | LG12 |
| NLRC3-like 5 | LG12 |
| nonfunctional kita gene 1 | Multi. hits |
| OR6N2-like 1 | Multi. hits |
| PARP4-like 1 | LG2 |
| PDE8B 3 | LG12 |
| RAB27B-like 1 | LG12 |
| RASGRF2-like 1,2 | LG12 |
| RSU1-like 4 | LG20 |
| S1PR3-like 4 | LG12 |
| SAPCD2 1 | LG12 |
| SNAPC4 1 | Unpl. Scaff. |
| stonustoxin subunit alpha-like 4 | LG14 |
| transposon Helitron gene-like 5 | Unpl. Scaff. |
| TRIM16-like 1,5 | LG17 |
| TRIM39-like 6 | Multi. hits |
| TTF1 1 | Unpl. Scaff. |
| UFM1 1 | LG21 |
| UGT2B31-like 1 | LG18 |
| uncharacterized protein K02A2.6-like 2,5 | LG7 |
| XYLB 4 | Unpl. Scaff. |
| ZBED1-like4 | LG12 |
| ZCCHC18-like2 | LG9 |
| ZNF146 4 | LG2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morris, J.; Darolti, I.; Bloch, N.I.; Wright, A.E.; Mank, J.E. Shared and Species-Specific Patterns of Nascent Y Chromosome Evolution in Two Guppy Species. Genes 2018, 9, 238. https://doi.org/10.3390/genes9050238
Morris J, Darolti I, Bloch NI, Wright AE, Mank JE. Shared and Species-Specific Patterns of Nascent Y Chromosome Evolution in Two Guppy Species. Genes. 2018; 9(5):238. https://doi.org/10.3390/genes9050238
Chicago/Turabian StyleMorris, Jake, Iulia Darolti, Natasha I. Bloch, Alison E. Wright, and Judith E. Mank. 2018. "Shared and Species-Specific Patterns of Nascent Y Chromosome Evolution in Two Guppy Species" Genes 9, no. 5: 238. https://doi.org/10.3390/genes9050238
APA StyleMorris, J., Darolti, I., Bloch, N. I., Wright, A. E., & Mank, J. E. (2018). Shared and Species-Specific Patterns of Nascent Y Chromosome Evolution in Two Guppy Species. Genes, 9(5), 238. https://doi.org/10.3390/genes9050238