Overview of Oxidative Stress Response Genes in Selected Halophilic Fungi



Abstract
1. Introduction
2. Materials and Methods
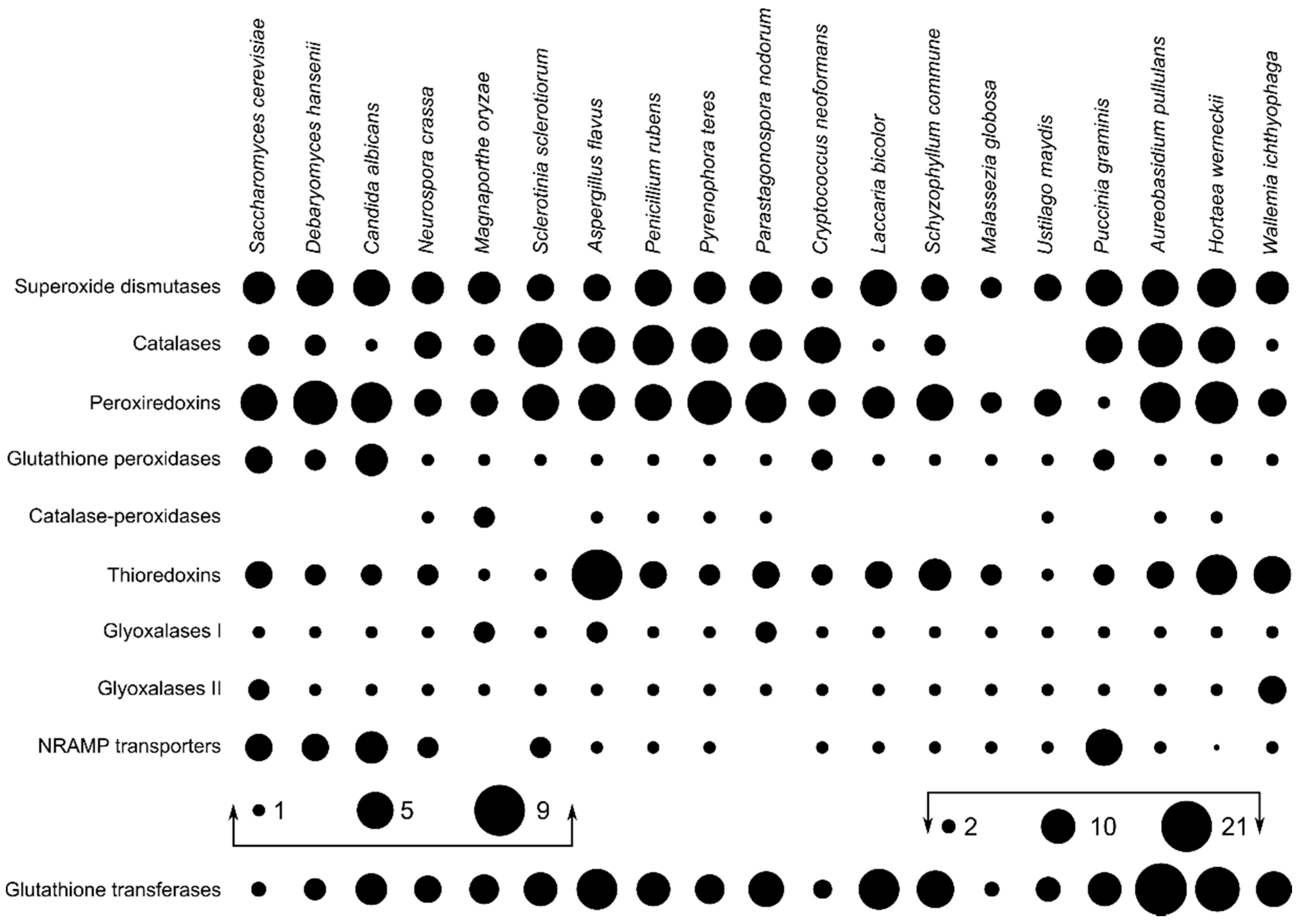
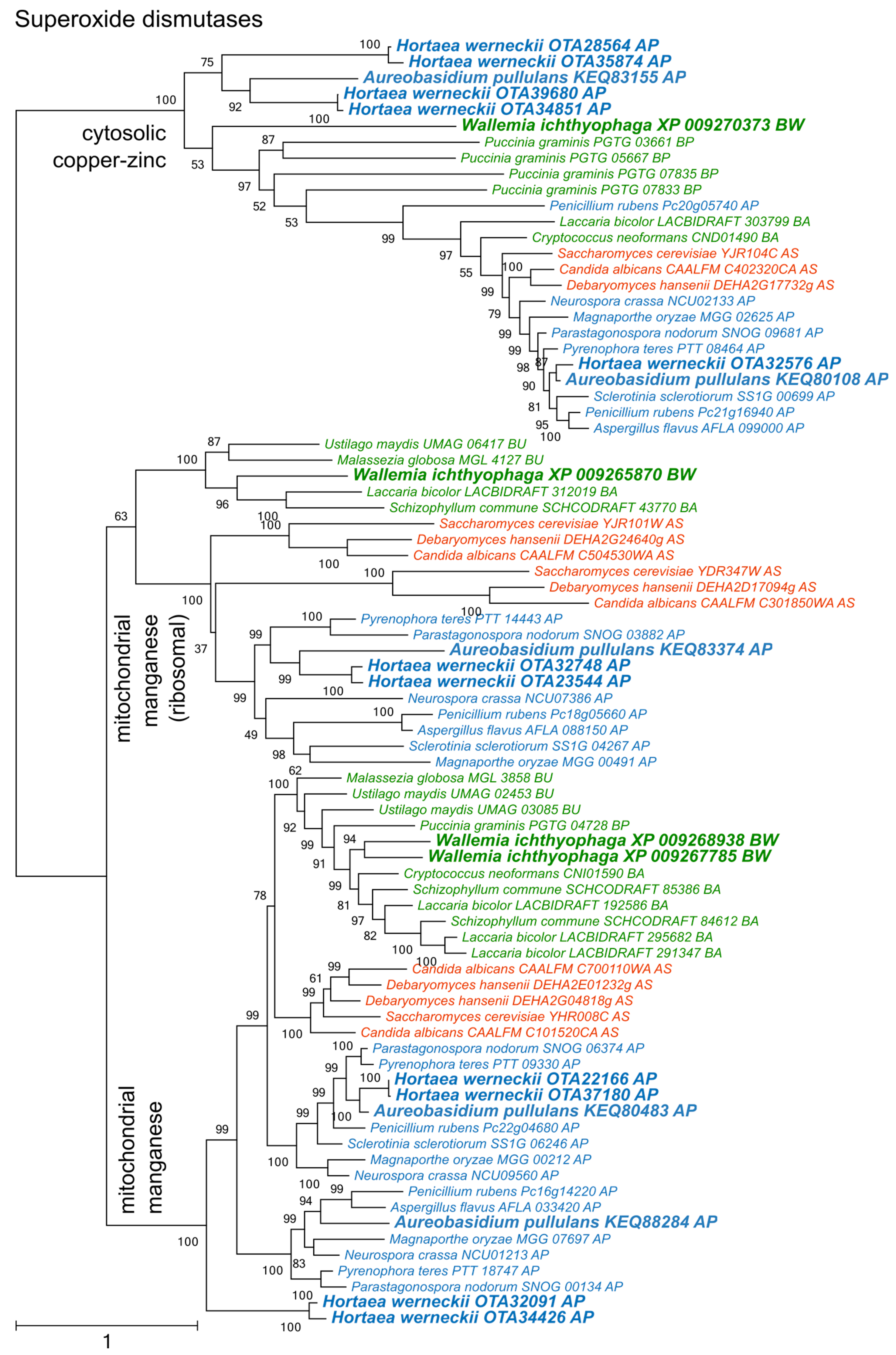
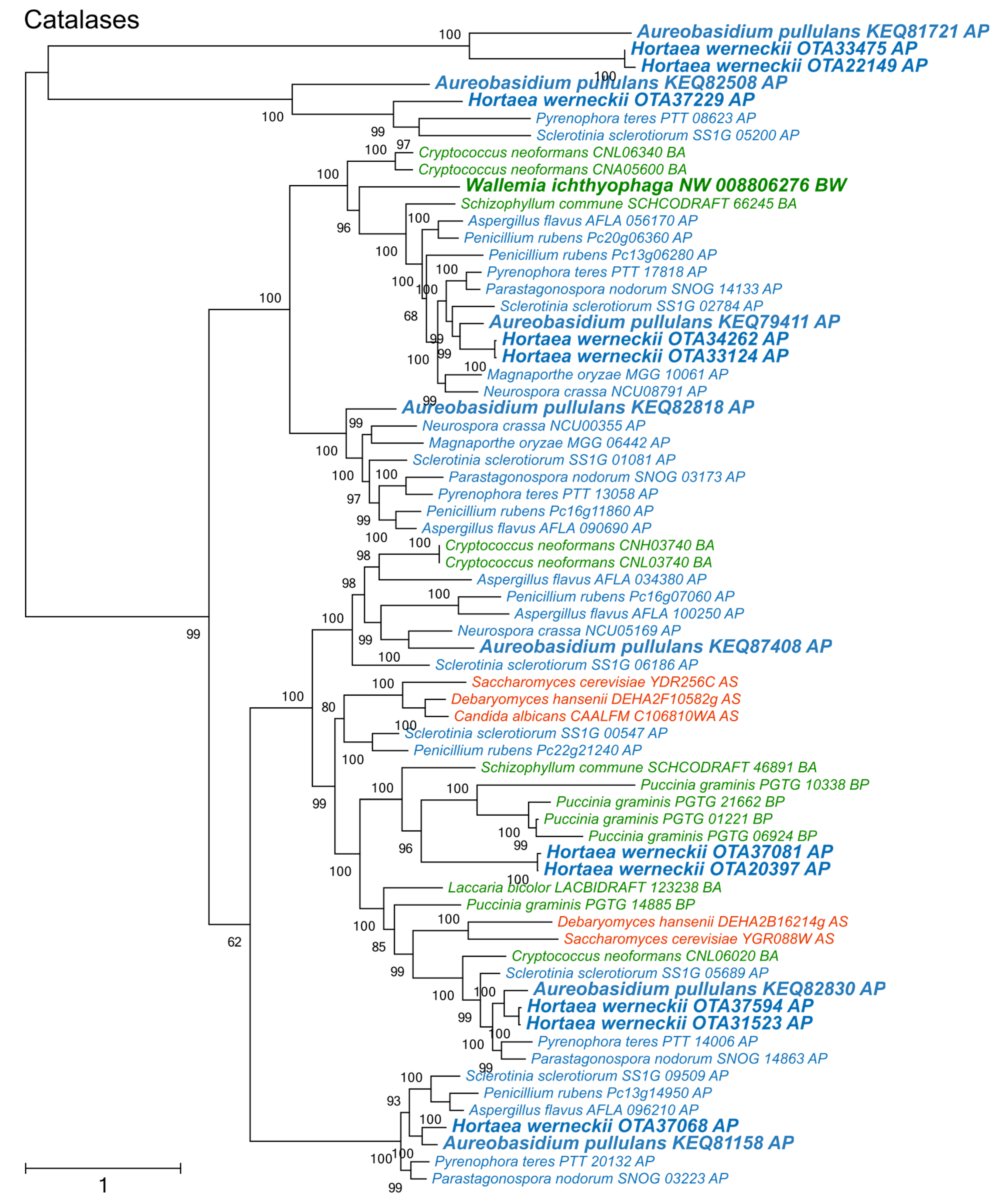
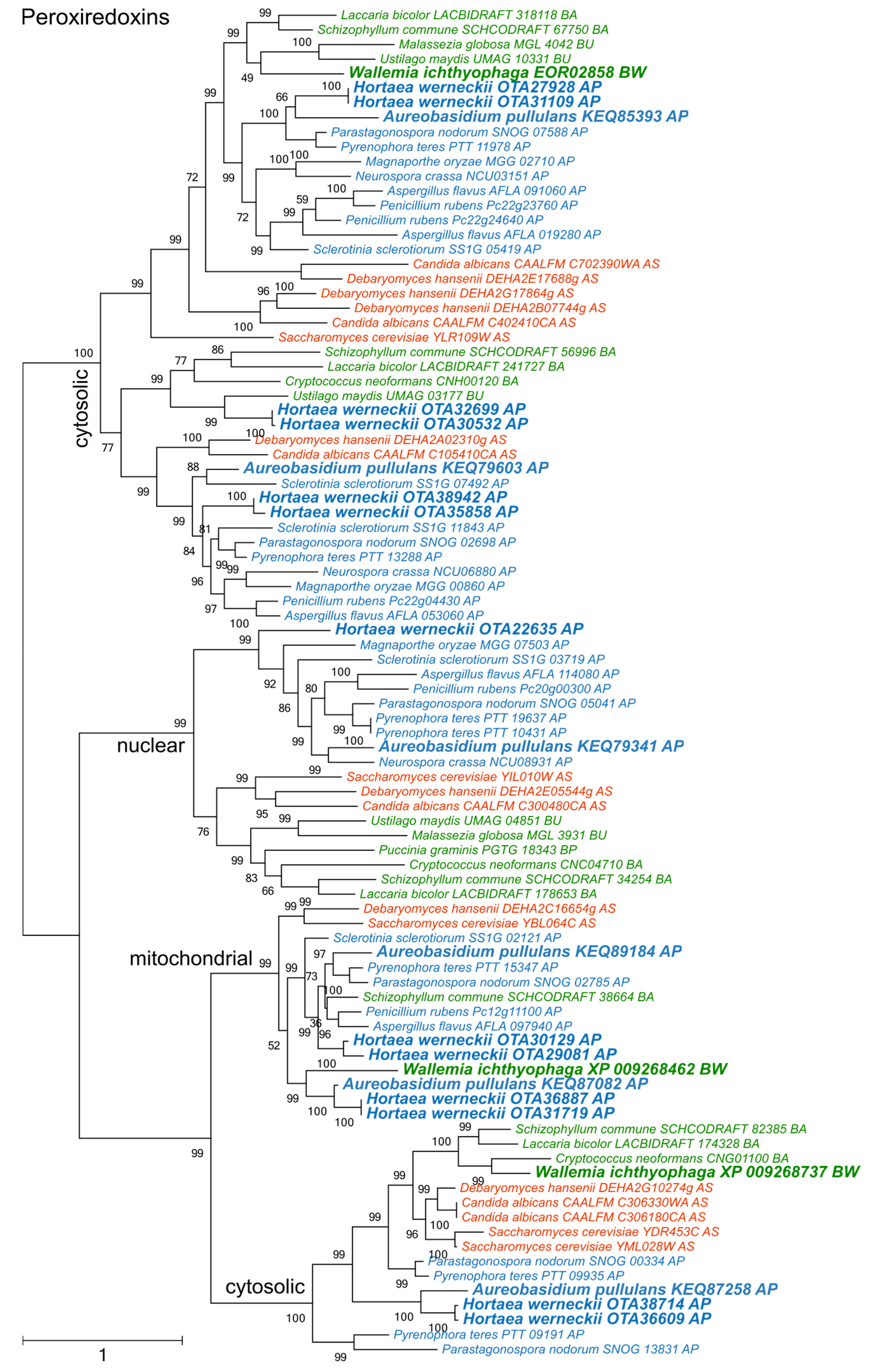
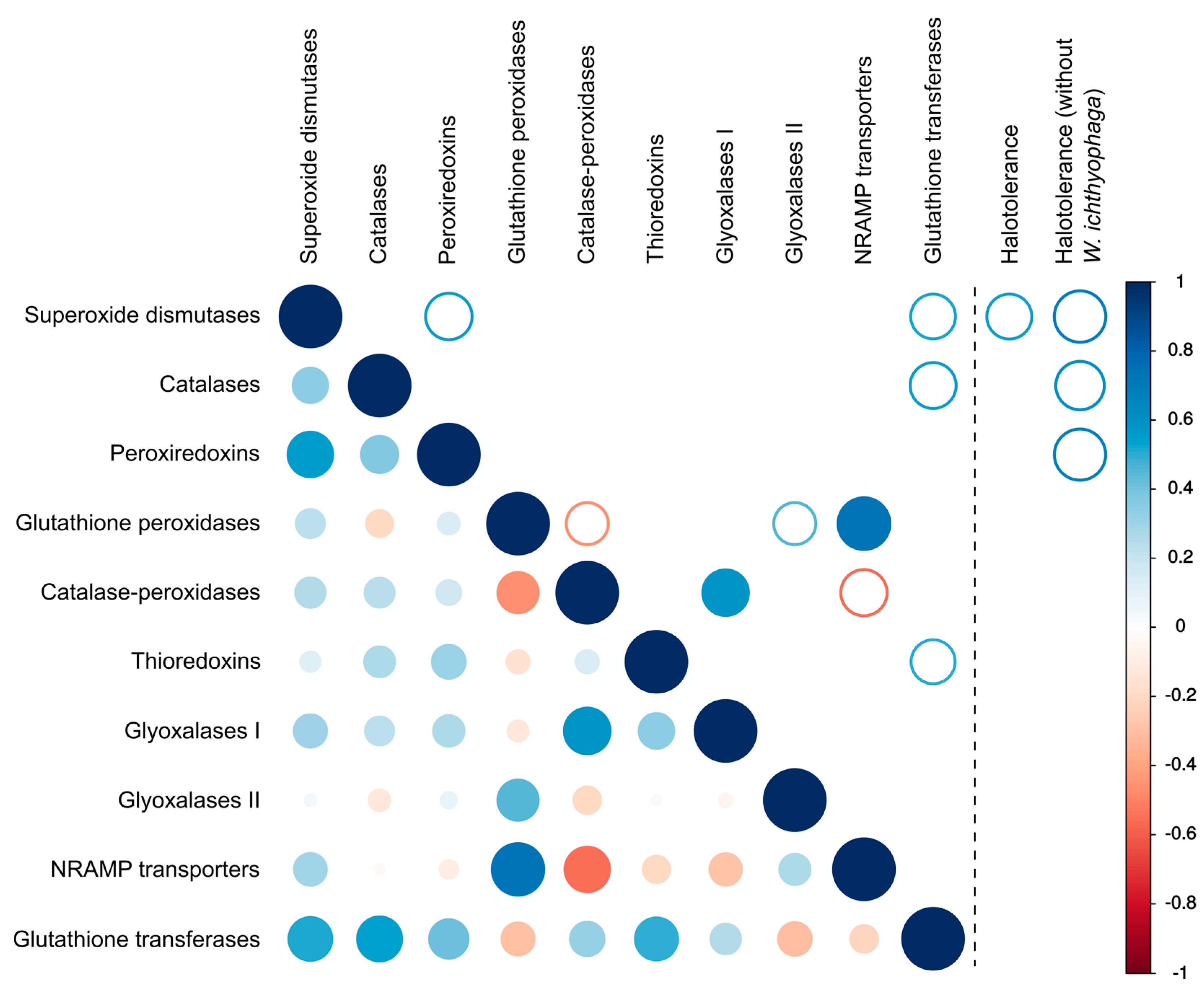
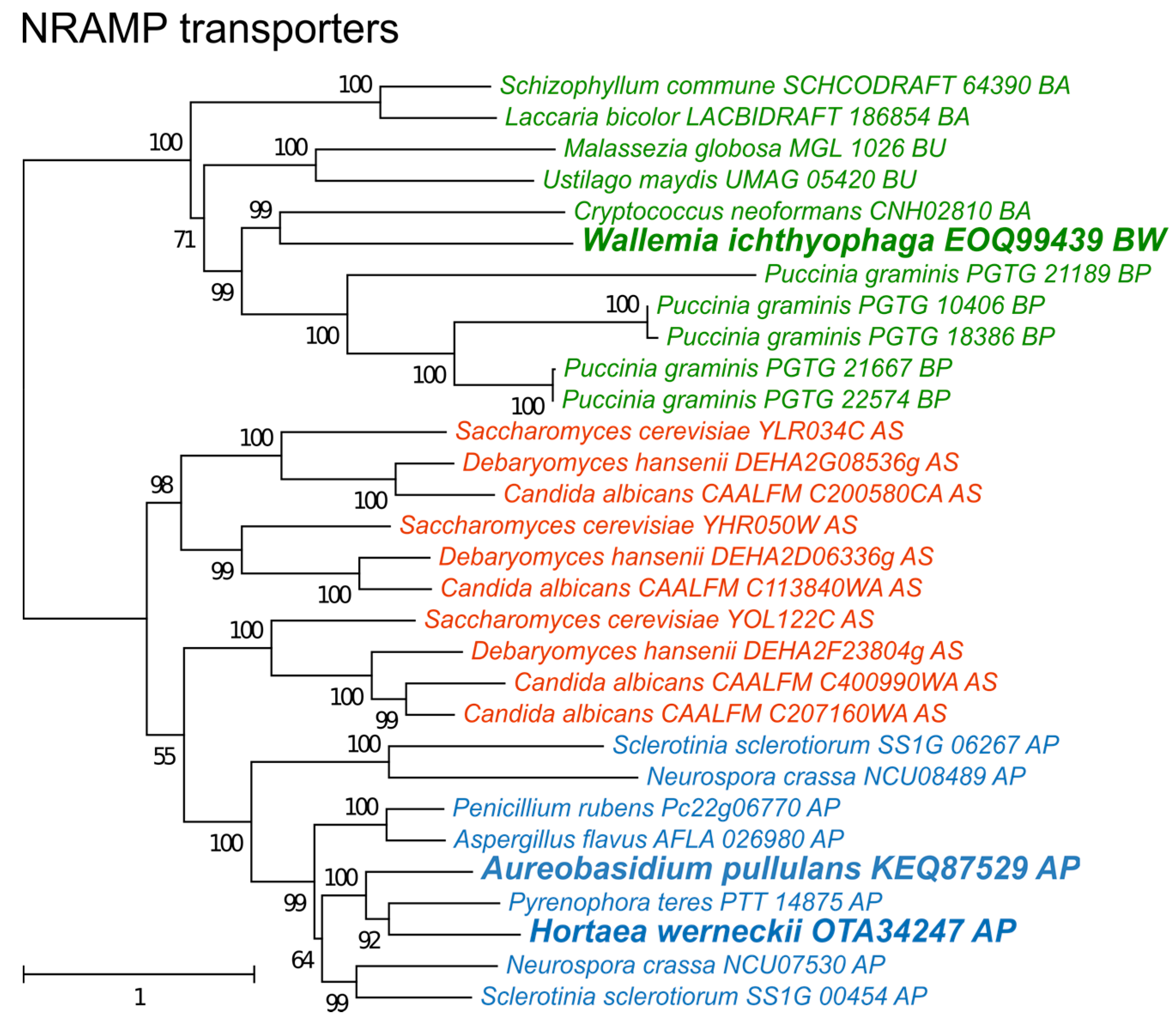
3. Results
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Morano, K.A.; Grant, C.M.; Moye-Rowley, W.S. The response to heat shock and oxidative stress in saccharomyces cerevisiae. Genetics 2012, 190, 1157–1195. [Google Scholar] [CrossRef] [PubMed]
- Garre, E.; Raginel, F.; Palacios, A.; Julien, A.; Matallana, E. Oxidative stress responses and lipid peroxidation damage are induced during dehydration in the production of dry active wine yeasts. Int. J. Food Microbiol. 2010, 136, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Nishio, K.; Usuki, Y.; Fujita, K.-I. Involvement of oxidative stress induction in Na+ toxicity and its relation to the inhibition of a Ca2+-dependent but calcineurin-independent mechanism in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2006, 101, 77–79. [Google Scholar] [CrossRef] [PubMed]
- Pang, C.-H.; Wang, B.-S.; Lüttge, U.; Beyschlag, W.; Büdel, B.; Francis, D. Oxidative stress and salt tolerance in plants. In Progress in Botany; Springer: Berlin/Heidelberg, Germany, 2008; Volume 69, pp. 231–245. [Google Scholar]
- Hohmann, S.; Magner, W. Yeast Stress Responses; Springer Science & Business Media: New York, NY, USA, 2003; Volume 53, ISBN 9788578110796. [Google Scholar]
- Daly, M.J. A new perspective on radiation resistance based on Deinococcus radiodurans. Nat. Rev. Microbiol. 2009, 7, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.-Y.; Lee, T.-H. Antioxidant enzymes as redox-based biomarkers: A brief review. BMB Rep. 2015, 48, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Daly, M.J. Death by protein damage in irradiated cells. DNA Repair 2012, 11, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Gaidamakova, E.K.; Grichenko, O.; Matrosova, V.Y.; Hoeke, V.; Klimenkova, P.; Conze, I.H.; Volpe, R.P.; Tkavc, R.; Gostinčar, C.; et al. Across the tree of life, radiation resistance is governed by antioxidant Mn2+, gauged by paramagnetic resonance. Proc. Natl. Acad. Sci. USA 2017, 114, E9253–E9260. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Omelchenko, M.V.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Lapidus, A.; Copeland, A.; Kim, E.; Land, M.; et al. Deinococcus geothermalis: The pool of extreme radiation resistance genes shrinks. PLoS ONE 2007, 2, e955. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Earl, A.M.; Howell, H.A.; Park, M.J.; Eisen, J.A.; Peterson, S.N.; Battista, J.R. Analysis of Deinococcus radiodurans’s transcriptional response to ionizing radiation and desiccation reveals novel proteins that contribute to extreme radioresistance. Genetics 2004, 168, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Gostinčar, C.; Lenassi, M.; Gunde-Cimerman, N.; Plemenitaš, A. Fungal adaptation to extremely high salt concentrations. Adv. Appl. Microbiol. 2011, 77, 71–96. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.D.T.; Jancic, S.; Meijer, M.; Tanney, J.B.; Zalar, P.; Gunde-Cimerman, N.; Seifert, K.A. Application of the phylogenetic species concept to Wallemia sebi from house dust and indoor air revealed by multi-locus genealogical concordance. PLoS ONE 2015, 10, e0120894. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.L.; Li, G.J.; Sánchez-Ramírez, S.; Stata, M.; Yang, Z.L.; Wu, G.; Dai, Y.C.; He, S.H.; Cui, B.K.; Zhou, J.L.; et al. A six-gene phylogenetic overview of Basidiomycota and allied phyla with estimated divergence times of higher taxa and a phyloproteomics perspective. Fungal Divers. 2017, 84, 43–74. [Google Scholar] [CrossRef]
- Zajc, J.; Džeroski, S.; Kocev, D.; Oren, A.; Sonjak, S.; Tkavc, R.; Gunde-Cimerman, N. Chaophilic or chaotolerant fungi: A new category of extremophiles? Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Gostinčar, C.; Grube, M.; Gunde-Cimerman, N. Evolution of fungal pathogens in domestic environments? Fungal Biol. 2011, 115, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, U. Role of oxidative stress in the extremely salt-tolerant yeast Hortaea werneckii. FEMS Yeast Res. 2006, 6, 816–822. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Petrovic, U.; Gunde-Cimerman, N.; Plemenitas, A. Cellular responses to environmental salinity in the halophilic black yeast Hortaea werneckii. Mol. Microbiol. 2002, 45, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Vaupotic, T.; Veranic, P.; Jenoe, P.; Plemenitas, A. Mitochondrial mediation of environmental osmolytes discrimination during osmoadaptation in the extremely halotolerant black yeast Hortaea werneckii. Fungal Genet. Biol. 2008, 45, 994–1007. [Google Scholar] [CrossRef] [PubMed]
- Gostinčar, C.; Ohm, R.A.; Kogej, T.; Sonjak, S.; Turk, M.; Zajc, J.; Zalar, P.; Grube, M.; Sun, H.; Han, J.; et al. Genome sequencing of four Aureobasidium pullulans varieties: Biotechnological potential, stress tolerance, and description of new species. BMC Genom. 2014, 15, 549. [Google Scholar] [CrossRef] [PubMed]
- Zajc, J.; Liu, Y.; Dai, W.; Yang, Z.; Hu, J.; Gostinčar, C.; Gunde-Cimerman, N. Genome and transcriptome sequencing of the halophilic fungus Wallemia ichthyophaga: Haloadaptations present and absent. BMC Genom. 2013, 14, 617. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Flibotte, S.; Neira, M.; Formby, S.; Plemenitas, A.; Cimerman, N.G.; Lenassi, M.; Gostincar, C.; Stajich, J.E.; Nislow, C. Insight into the recent genome duplication of the halophilic yeast Hortaea werneckii: Combining an improved genome with gene expression and chromatin structure. G3 Genes Genomes Genet. 2017, 7, 2015–2022. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M. Toward pathway engineering: a new database of genetic and molecular pathways. Sci. Technol. Jpn. 1996, 59, 34–38. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Shaffer, A.A.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- NCBI Resource Coordinators. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2017, 45, D12–D17. [Google Scholar] [CrossRef]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage, 2nd ed.; Aspen Publishers, Inc.: Dordrecht, UK, 1999; ISBN 978-0-387–92206–5. [Google Scholar]
- Krauke, Y.; Sychrova, H. Four pathogenic Candida species differ in salt tolerance. Curr. Microbiol. 2010, 61, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Charlang, G.W.; Horowitz, N.H. Germination and growth of Neurospora at low water activities. Proc. Natl. Acad. Sci. USA 1971, 68, 260–262. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, K.K.; Sankaralingam, A.; Nakkeeran, S. Effect of temperature and salinity on the growth of Sclerotinia sclerotiorum causing head rot of cabbage. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 950–954. [Google Scholar] [CrossRef]
- El-Mougith, A.A. The effect of salinity on some halophilic soil fungi. In Towards the Rational Use of High Salinity Tolerant Plants Tasks for Vegetation Science; Springer Science & Business Media: New York, NY, USA, 1993; Volume 27, pp. 473–477. ISBN 978-94–010–4821–7. [Google Scholar]
- Castillo, G.; Demoulin, V. NaCl salinity and temperature effects on growth of three wood-rotting basidiomycetes from a Papua New Guinea coastal forest. Mycol. Res. 1997, 101, 341–344. [Google Scholar] [CrossRef]
- Baroni, A.; De Rosa, R.; De Rosa, A.; Donnarumma, G.; Catalanotti, P. New strategies in dandruff treatment: Growth control of Malassezia ovalis. Dermatology 2000, 201, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Benito, B.; Garciadeblás, B.; Pérez-Martín, J.; Rodríguez-Navarro, A. Growth at high pH and sodium and potassium tolerance in media above the cytoplasmic pH depend on ENA ATPases in Ustilago maydis. Eukaryot. Cell 2009, 8, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Kuma, K.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.; Doallo, R.; Posada, D. ProtTest 3: fast selection of best-fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing 2017; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix (Version 0.84). Available online: https://github.com/taiyun/corrplot (accessed on 27 December 2017).
- Gostinčar, C.; Stajich, J.E.; Zupančič, J.; Zalar, P.; Gunde-Cimerman, N. Genomic evidence for intraspecific hybridization in a clonal and extremely halotolerant yeast. BMC Genom. 2018. submitted. [Google Scholar]
- Krantz, M.; Becit, E.; Hohmann, S. Comparative genomics of the HOG-signalling system in fungi. Curr. Genet. 2006, 49, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Vaupotič, T.; Plemenitaš, A. Differential gene expression and HogI interaction with osmoresponsive genes in the extremely halotolerant black yeast Hortaea werneckii. BMC Genom. 2007, 8, 280–295. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, C.; Siles, A.; Martinez, J.L.; Calero, F.; Ramos, J. Oxidative stress sensitivity in Debaryomyces hansenii. FEMS Yeast Res. 2009, 9, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Zámocký, M.; Gasselhuber, B.; Furtmüller, P.G.; Obinger, C. Molecular evolution of hydrogen peroxide degrading enzymes. Arch. Biochem. Biophys. 2012, 525, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Zalar, P.; Sybren de Hoog, G.; Schroers, H.-J.; Frank, J.M.; Gunde-Cimerman, N. Taxonomy and phylogeny of the xerophilic genus Wallemia (Wallemiomycetes and Wallemiales, cl. et ord. nov.). Antonie Van Leeuwenhoek 2005, 87, 311–328. [Google Scholar] [CrossRef] [PubMed]
- Zajc, J.; Kogej, T.; Ramos, J.; Galinski, E.A.; Gunde-Cimerman, N. The osmoadaptation strategy of the most halophilic fungus Wallemia ichthyophaga, growing optimally at salinities above 15% NaCl. Appl. Environ. Microbiol. 2014, 80, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, S.; Ishii-Minami, N.; Saitoh, K.-I.; Otake, Y.; Kaku, H.; Shibuya, N.; Nishizawa, Y.; Minami, E. The role of catalase-peroxidase secreted by Magnaporthe oryzae during early infection of rice cells. Mol. Plant Microbe Interact. 2011, 24, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Molina, L.; Kahmann, R. An Ustilago maydis gene involved in H2O2 detoxification is required for virulence. Plant Cell 2007, 19, 2293–2309. [Google Scholar] [CrossRef] [PubMed]
- Omelchenko, M.V.; Wolf, Y.I.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Daly, M.J.; Koonin, E.V.; Makarova, K.S. Comparative genomics of Thermus thermophilus and Deinococcus radiodurans: Divergent routes of adaptation to thermophily and radiation resistance. BMC Evol. Biol. 2005, 5, 57. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Phylogenetic Position | Maximum Tolerated Concentration of NaCl (w/v) |
|---|---|---|
| Saccharomyces cerevisiae | Ascomycota, Saccharomycotina | 10 (our unpublished data) |
| Debaryomyces hansenii | Ascomycota, Saccharomycotina | 24 [26] |
| Candida albicans | Ascomycota, Saccharomycotina | 12.2 [27] |
| Neurospora crassa | Ascomycota, Pezizomycotina | 12.8 [28] |
| Magnaporthe oryzae | Ascomycota, Pezizomycotina | 8 [29] |
| Sclerotinia sclerotiorum | Ascomycota, Pezizomycotina | N.D. 1 |
| Aspergillus flavus | Ascomycota, Pezizomycotina | 10 [30] |
| Penicillium rubens | Ascomycota, Pezizomycotina | N.D. 1 (20 for related P. notatum and P. chrysogenum [30]) |
| Pyrenophora teres | Ascomycota, Pezizomycotina | N.D. 1 |
| Parastagonospora nodorum | Ascomycota, Pezizomycotina | N.D. 1 |
| Cryptococcus neoformans var. neoformans | Basidiomycota, Agaricomycotina | 16 (our unpublished data) |
| Laccaria bicolor | Basidiomycota, Agaricomycotina | N.D. 1 |
| Schizophyllum commune | Basidiomycota, Agaricomycotina | 7 [31] |
| Malassezia globosa | Basidiomycota, Ustilaginomycotina | N.D. 1 (5.5 for related M. furfur [32]) |
| Ustilago maydis | Basidiomycota, Ustilaginomycotina | 3.5 [33] |
| Puccinia graminis | Basidiomycota, Pucciniomycotina | N.D. 1 |
| Hortaea werneckii | Ascomycota, Pezizomycotina | 28 [12] |
| Aureobasidium pullulans | Ascomycota, Pezizomycotina | 17 [16] |
| Wallemia ichthyophaga | Basidiomycota, Wallemiomycotina | 35.9 [12] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gostinčar, C.; Gunde-Cimerman, N. Overview of Oxidative Stress Response Genes in Selected Halophilic Fungi. Genes 2018, 9, 143. https://doi.org/10.3390/genes9030143
Gostinčar C, Gunde-Cimerman N. Overview of Oxidative Stress Response Genes in Selected Halophilic Fungi. Genes. 2018; 9(3):143. https://doi.org/10.3390/genes9030143
Chicago/Turabian StyleGostinčar, Cene, and Nina Gunde-Cimerman. 2018. "Overview of Oxidative Stress Response Genes in Selected Halophilic Fungi" Genes 9, no. 3: 143. https://doi.org/10.3390/genes9030143
APA StyleGostinčar, C., & Gunde-Cimerman, N. (2018). Overview of Oxidative Stress Response Genes in Selected Halophilic Fungi. Genes, 9(3), 143. https://doi.org/10.3390/genes9030143