Antibiotic Resistance Genetic Markers and Integrons in White Soft Cheese: Aspects of Clinical Resistome and Potentiality of Horizontal Gene Transfer


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Processing and DNA Extraction
2.2. Repetitive Extragenic Palindromic Sequences Polymerase Chain Reaction
2.3. Screening of Antibiotic Resistance Genetic Markers and Integrons
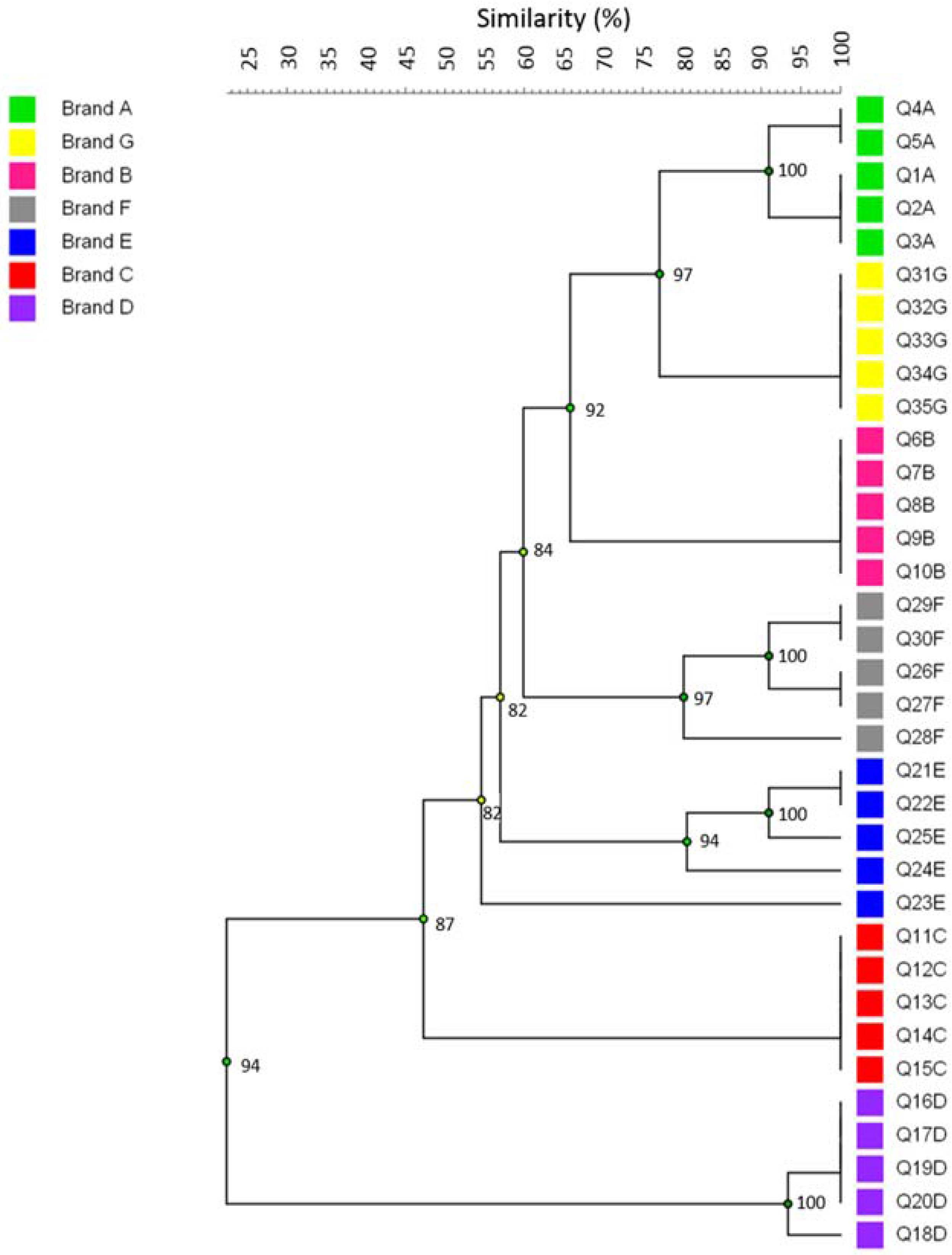
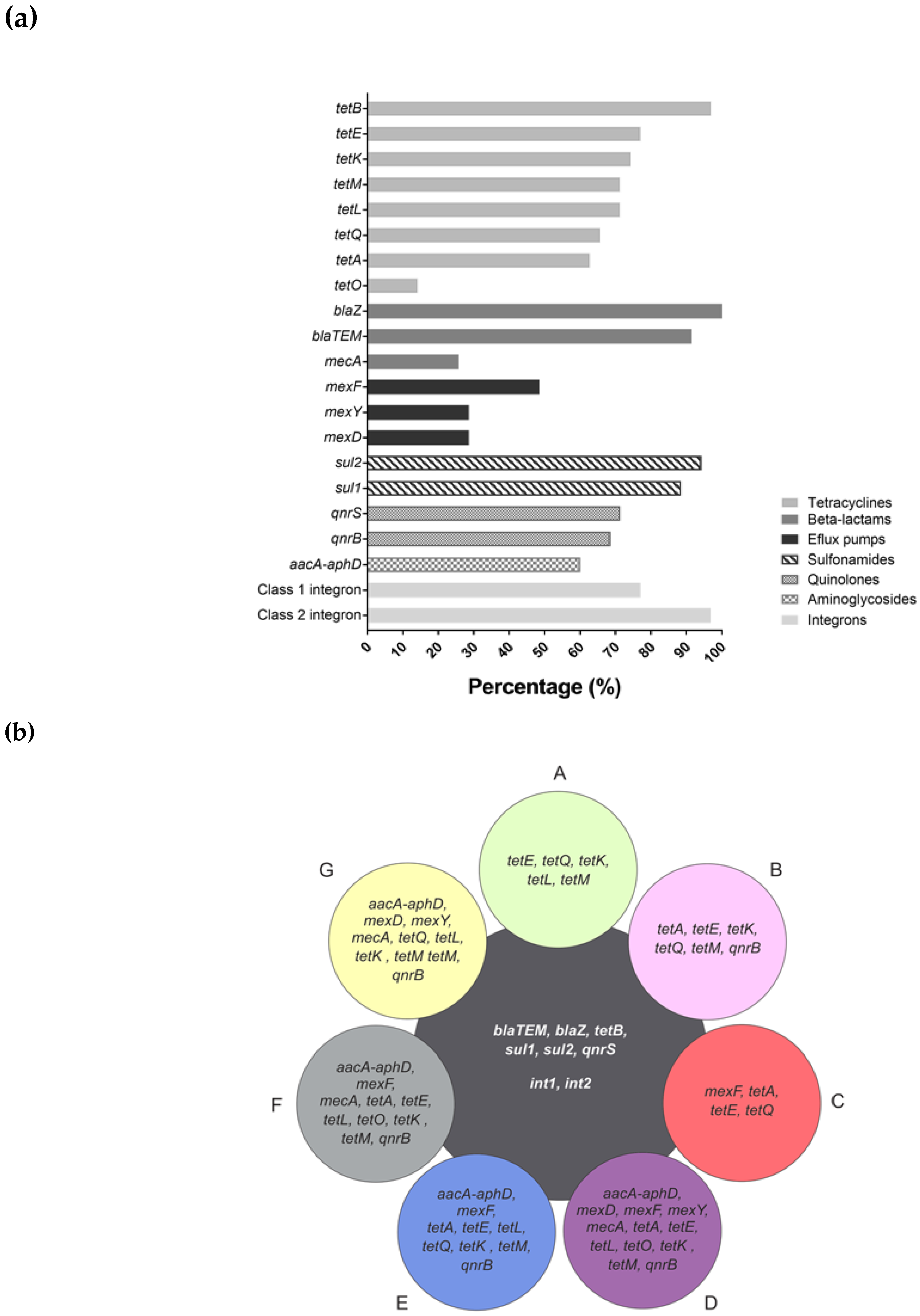
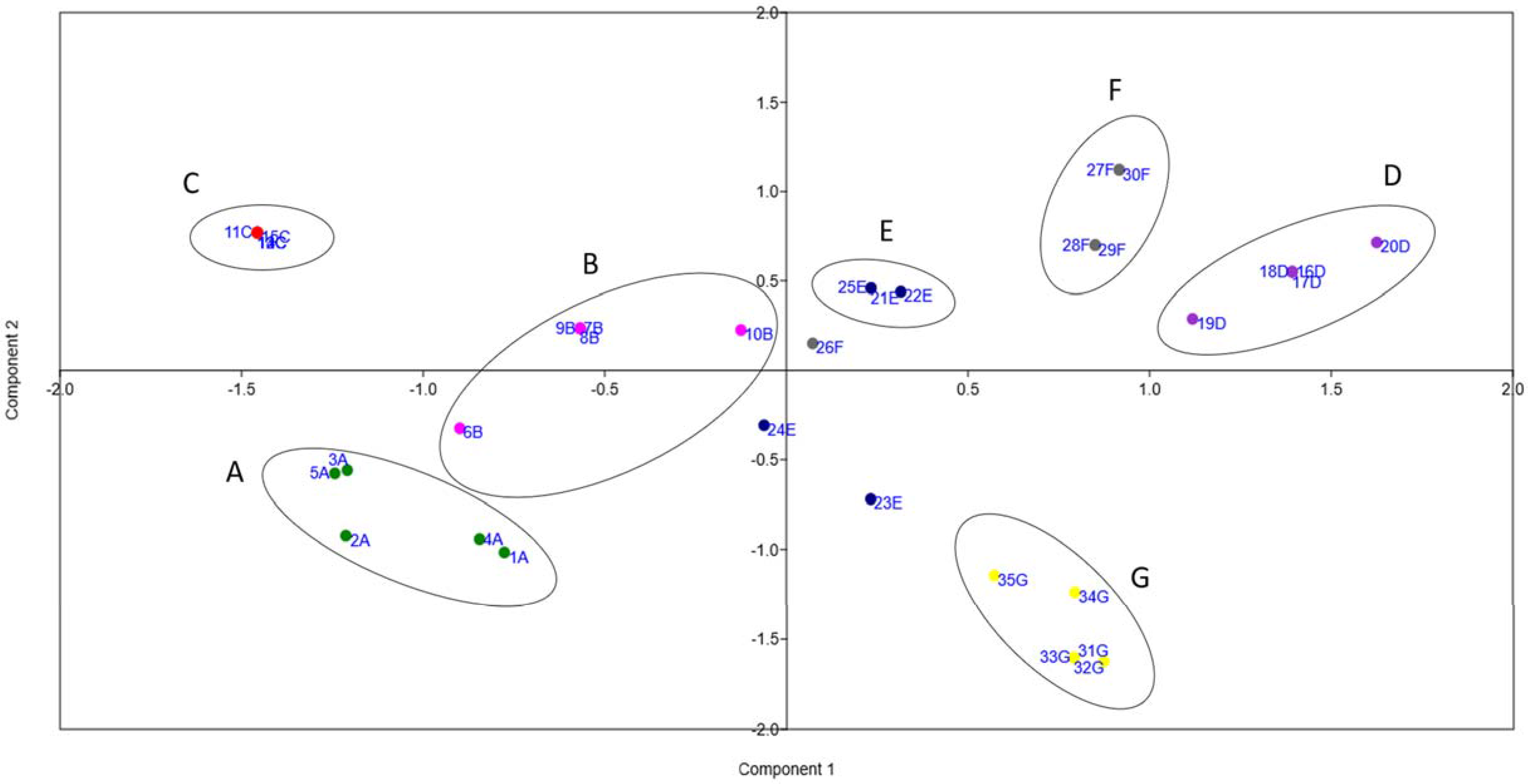
3. Results
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Roca, I.; Akova, M.; Baquero, F.; Carlet, J.; Cavaleri, M.; Coenen, S.; Cohen, J.; Findlay, D.; Gyssens, I.; Heure, O.E.; et al. The global threat of antimicrobial resistance: Science for intervention. New Microbes New Infect. 2015, 6, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Landers, T.F.; Cohen, B.; Wittum, T.E.; Larson, E.L. A Review of antibiotic use in food animals: Perspective, policy, and potential. Public Health Rep. 2012, 127, 4–22. [Google Scholar] [CrossRef] [PubMed]
- Founou, L.L.; Founou, R.C.; Essack, S.Y. Antibiotic resistance in the food chain: A developing country-perspective. Front. Microbiol. 2016, 7, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson-Palme, J. Antibiotic resistance in the food supply chain: Where can sequencing and metagenomics aid risk assessment? Curr. Opin. Food Sci. 2017, 14, 66–71. [Google Scholar] [CrossRef]
- Verraes, C.; van Boxstael, S.; van Meervenne, E.; van Coillie, E.; Butaye, P.; Catry, B.; Schaetzen, M.A.; van Huffel, X.; Imberechts, H.; Dierick, K.; et al. Antimicrobial resistance in the food chain: A review. Int. J. Environ. Res. Public Health 2013, 10, 2643–2669. [Google Scholar] [CrossRef] [PubMed]
- Doyle, M.E. Multidrug-resistant pathogens in the food supply. Foodborne Pathog. Dis. 2015, 12, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Huddleston, J.R. Horizontal gene transfer in the human gastrointestinal tract: Potential spread of antibiotic resistance genes. Infect. Drug Resist. 2014, 7, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Devirgiliis, C.; Barile, S.; Perozzi, G. Antibiotic resistance determinants in the interplay between food and gut microbiota. Genes Nutr. 2011, 6, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Rolain, J.M. Food and human gut as reservoirs of transferable antibiotic resistance encoding genes. Front. Microbiol. 2013, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Van Hoek, A.H.A.M.; Mevius, D.; Guerra, B.; Mullany, P.; Roberts, A.P.; Aarts, H.J.M. Acquired antibiotic resistance genes: An overview. Front. Microbiol. 2011, 2, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Gillings, M.R. Integrons: Past, present, and future. Microbiol. Mol. Biol. Rev. 2014, 78, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Bao, X.; Ji, L.; Chen, L.; Liu, J.; Miao, J.; Chen, D.; Bian, H.; Li, Y.; Yu, G. Resistance integrons: Class 1, 2 and 3 integrons. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 45. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Singh, R. Antibiotic resistance in food lactic acid bacteria—A review. Int. J. Food Microbiol. 2005, 105, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Gomes, B.C.; Franco, B.D.G.D.M.; De Martinis, E.C.P. Microbiological food safety issues in Brazil: Bacterial pathogens. Foodborne Pathog. Dis. 2013, 10, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, J.D.G.; Viotto, W.H.; Kuaye, A.Y. The quality of Minas Frescal cheese produced by different technological processes. Food Control 2007, 18, 262–267. [Google Scholar] [CrossRef]
- Brito, J.R.F.; Santos, E.M.P.; Arcuri, E.F.; Lange, C.C.; Brito, M.A.V.P.; Souza, G.N.; Cerqueira, M.M.P.O.; Beltran, J.M.S.; Call, J.E.; Liu, Y.; et al. Retail survey of Brazilian milk and Minas Frescal cheese and a contaminated dairy plant to establish prevalence, relatedness, and sources of Listeria monocytogenes isolates. Appl. Environ. Microbiol. 2008, 74, 4954–4961. [Google Scholar] [CrossRef] [PubMed]
- Visotto, R.G.; Oliveira, M.A.; Prado, S.O.T.; Bergamini, A.M.M. Queijo Minas Frescal: Perfil higiênico-sanitário e avaliação da rotulagem. Rev. Inst. Adolfo Lutz 2011, 70, 8–15. [Google Scholar]
- Fontes, C.O.; Silva, V.L.; de Paiva, M.R.B.; Garcia, R.A.; Resende, J.A.; Ferreira-Machado, A.B.; Diniz, C.G. Prevalence, antimicrobial resistance, and virulence characteristics of mecA-encoding coagulase-negative Staphylococci isolated from soft cheese in Brazil. J. Food Sci. 2013, 78, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Nunes, C.S.R.; Souza, C.P.; Pereira, K.S.; Del Aguila, E.M.; Paschoalin, V.M.F. Identification and molecular phylogeny of coagulase-negative staphylococci isolates from Minas Frescal cheese in southeastern Brazil: Superantigenic toxin production and antibiotic resistance. J. Dairy Sci. 2016, 99, 2641–2653. [Google Scholar] [CrossRef] [PubMed]
- Belén Flórez, A.; Alegría, Á.; Rossi, F.; Delgado, S.; Felis, G.E.; Torriani, S.; Mayo, B. Molecular identification and quantification of tetracycline and erythromycin resistance genes in Spanish and Italian retail cheeses. BioMed Res. Int. 2014. [Google Scholar] [CrossRef]
- Amador, P.; Fernandes, R.; Prudêncio, C.; Brito, L. Resistance to β-lactams in bacteria isolated from different types of Portuguese cheese. Int. J. Mol. Sci. 2009, 10, 1538–1551. [Google Scholar] [CrossRef] [PubMed]
- Furlaneto-Maia, L.; Rocha, K.R.; Henrique, F.C.; Giazzi, A.; Furlaneto, M.C. Antimicrobial resistance in Enterococcus sp isolated from soft cheese in Southern Brazil. Adv. Microbiol. 2014, 4, 175–181. [Google Scholar] [CrossRef]
- Kürekci, C. Short communication: Prevalence, antimicrobial resistance, and resistant traits of coagulase-negative staphylococci isolated from cheese samples in Turkey. J. Dairy Sci. 2016, 99, 2675–2679. [Google Scholar] [CrossRef] [PubMed]
- Perry, J.A.; Wright, G.D. The antibiotic resistance “mobilome”: Searching for the link between environment and clinic. Front. Microbiol. 2013, 4, 138. [Google Scholar] [CrossRef] [PubMed]
- Gaber, A.; Hassan, M.M.; Dessoky, E.-D.S.; Attia, A.O. In vitro antimicrobial comparison of Taif and Egyptian Pomegranate Peels and seeds extracts. J. Appl. Biol. Biotechnol. 2015, 3, 12–17. [Google Scholar] [CrossRef]
- Perin, L.M.; Sardaro, M.L. S; Nero, L.A.; Neviani, E.; Gatti, M. Bacterial ecology of artisanal Minas cheeses assessed by culture-dependent and -independent methods. Food Microbiol. 2017, 65, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Perin, L.M.; Bello, B.D.; Belviso, S.; Zeppa, G.; Carvalho, A.F.; Cocolin, F.; Nero, L.A. Microbiota of Minas cheese as influenced by the nisin producer Lactococcus lactis subsp. lactis GLc05. Int. J. Food Microbiol. 2015, 214, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Devirgiliis, C.; Caravelli, A.; Coppola, D.; Barile, S.; Perozzi, G. Antibiotic resistance and microbial composition along the manufacturing process of Mozzarella di Bufala Campana. Int. J. Food Microbiol. 2008, 128, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Grossman, T.H. Tetracycline antibiotics and resistance. Cold Spring Harb. Perspect. Med. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Huys, G.; D’Haene, K.; Collard, J.-M.; Swings, J. Prevalence and molecular characterization of tetracycline resistance in Enterococcus isolates from food. Appl. Environ. Microbiol. 2004, 70, 1555–141032. [Google Scholar] [CrossRef] [PubMed]
- Devirgiliis, C.; Zinno, P.; Stirpe, M.; Barile, S.; Perozzi, G. Functional screening of antibiotic resistance genes from a representative metagenomic library of food fermenting microbiota. BioMed Res. Int. 2014, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Carfora, V.; Caprioli, A.; Marri, N.; Sagrafoli, D.; Boselli, C.; Giacinti, G.; Giangolini, G.; Sorbara, L.; Dottarelli, S.; Battisti, A.; et al. Enterotoxin genes, enterotoxin production, and methicillin resistance in Staphylococcus aureus isolated from milk and dairy products in Central Italy. Int. Dairy J. 2015, 42, 12–15. [Google Scholar] [CrossRef]
- Jamali, H.; Paydar, M.; Radmehr, B.; Ismail, S.; Dadrasnia, A. Prevalence and antimicrobial resistance of Staphylococcus aureus isolated from raw milk and dairy products. Food Control 2006, 54, 383–388. [Google Scholar] [CrossRef]
- Spanu, V.; Spanu, C.; Virdis, S.; Cossu, F.; Scarano, C.; De Santis, E.P.L. Virulence factors and genetic variability of Staphylococcus aureus strains isolated from raw sheep’s milk cheese. Int. J. Food Microbiol. 2012, 153, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Jaimee, G.; Halami, P.M. Emerging resistance to aminoglycosides in lactic acid bacteria of food origin—An impending menace. App. Microbiol. Biotechnol. 2016, 100, 1137–1151. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Martínez, J.M.; Cano, M.E.; Velasco, C.; Martínez-Martínez, L.; Pascual, Á. Plasmid-mediated quinolone resistance: An update. J. Infect. Chemother. 2011, 17, 149–182. [Google Scholar] [CrossRef] [PubMed]
- Phuong Hoa, P.T.; Nonaka, L.; Hung Viet, P.; Suzuki, S. Detection of the sul1, sul2, and sul3 genes in sulfonamide-resistant bacteria from wastewater and shrimp ponds of north Vietnam. Sci. Total Environ. 2008, 405, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Sköld, O. Sulfonamide resistance: Mechanisms and trends. Drug Resist. Updat. 2000, 3, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Piddock, L.J.V. Multidrug-resistance efflux pumps—Not just for resistance. Nat. Rev. Microbiol. 2006, 4, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Blanco, P.; Hernando-Amado, S.; Reales-Calderon, J.A.; Corona, F.; Lira, F.; Alcalde-Rico, M.; Bernardini, A.; Sanchez, M.B.; Martinez, J.L. Bacterial multidrug efflux pumps: Much more than antibiotic resistance determinants. Microorganisms 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Kievit, T.; Parkins, M.; Gillis, R.; Srikumar, R.; Ceri, H.; Poole, K.; Iglewski, B.H.; Storey, D. Multidrug efflux pumps: Expression patterns and contribution to antibiotic resistance in Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 2001, 45, 1761–1770. [Google Scholar] [CrossRef] [PubMed]
- Ueda, O.; Wexler, H.M.; Hirai, K.; Shibata, Y.; Yoshimura, F.; Fujimura, S. Sixteen homologs of the Mex-type multidrug resistance efflux pump in Bacteroides fragilis. Antimicrob. Agents Chemother. 2005, 49, 2807–2815. [Google Scholar] [CrossRef] [PubMed]
- Galvani, J.W.C.; Tondo, E.C.; Brandelli, A. Commonly screened antibiotics in raw milk from dairy plants under State Inspection in Rio Grande do Sul, Brazil. Braz. J. Vet. Res. Anim. Sci. 2017, 57, 147–153. [Google Scholar] [CrossRef]
- Cambray, G.; Guerout, A.-M.; Mazel, D. Integrons. Ann. Rev. Genet. 2010, 44, 141–166. [Google Scholar] [CrossRef] [PubMed]
- Mazel, D. Integrons: Agents of bacterial evolution. Nat. Rev. Microbiol. 2006, 4, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Tsafnat, G.; Coiera, E.; Iredell, J.R. Gene cassettes and cassette arrays in mobile resistance integrons: Review article. FEMS Microbiol. Rev. 2009, 33, 757–784. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.M.; Shimamoto, T.; Shimamoto, T. Characterization of integrons and resistance genes in multidrug-resistant Salmonella enterica isolated from meat and dairy products in Egypt. Int. J. Food Microbiol. 2014, 189, 39–44. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Paula, A.C.L.; Medeiros, J.D.; De Azevedo, A.C.; De Assis Chagas, J.M.; Da Silva, V.L.; Diniz, C.G. Antibiotic Resistance Genetic Markers and Integrons in White Soft Cheese: Aspects of Clinical Resistome and Potentiality of Horizontal Gene Transfer. Genes 2018, 9, 106. https://doi.org/10.3390/genes9020106
De Paula ACL, Medeiros JD, De Azevedo AC, De Assis Chagas JM, Da Silva VL, Diniz CG. Antibiotic Resistance Genetic Markers and Integrons in White Soft Cheese: Aspects of Clinical Resistome and Potentiality of Horizontal Gene Transfer. Genes. 2018; 9(2):106. https://doi.org/10.3390/genes9020106
Chicago/Turabian StyleDe Paula, Ana Caroline L., Julliane D. Medeiros, Analice C. De Azevedo, Jéssica M. De Assis Chagas, Vânia L. Da Silva, and Cláudio G. Diniz. 2018. "Antibiotic Resistance Genetic Markers and Integrons in White Soft Cheese: Aspects of Clinical Resistome and Potentiality of Horizontal Gene Transfer" Genes 9, no. 2: 106. https://doi.org/10.3390/genes9020106
APA StyleDe Paula, A. C. L., Medeiros, J. D., De Azevedo, A. C., De Assis Chagas, J. M., Da Silva, V. L., & Diniz, C. G. (2018). Antibiotic Resistance Genetic Markers and Integrons in White Soft Cheese: Aspects of Clinical Resistome and Potentiality of Horizontal Gene Transfer. Genes, 9(2), 106. https://doi.org/10.3390/genes9020106