Ancient DNA Resolves the History of Tetragnatha (Araneae, Tetragnathidae) Spiders on Rapa Nui

 ,
,
Abstract
:1. Introduction
2. Methods
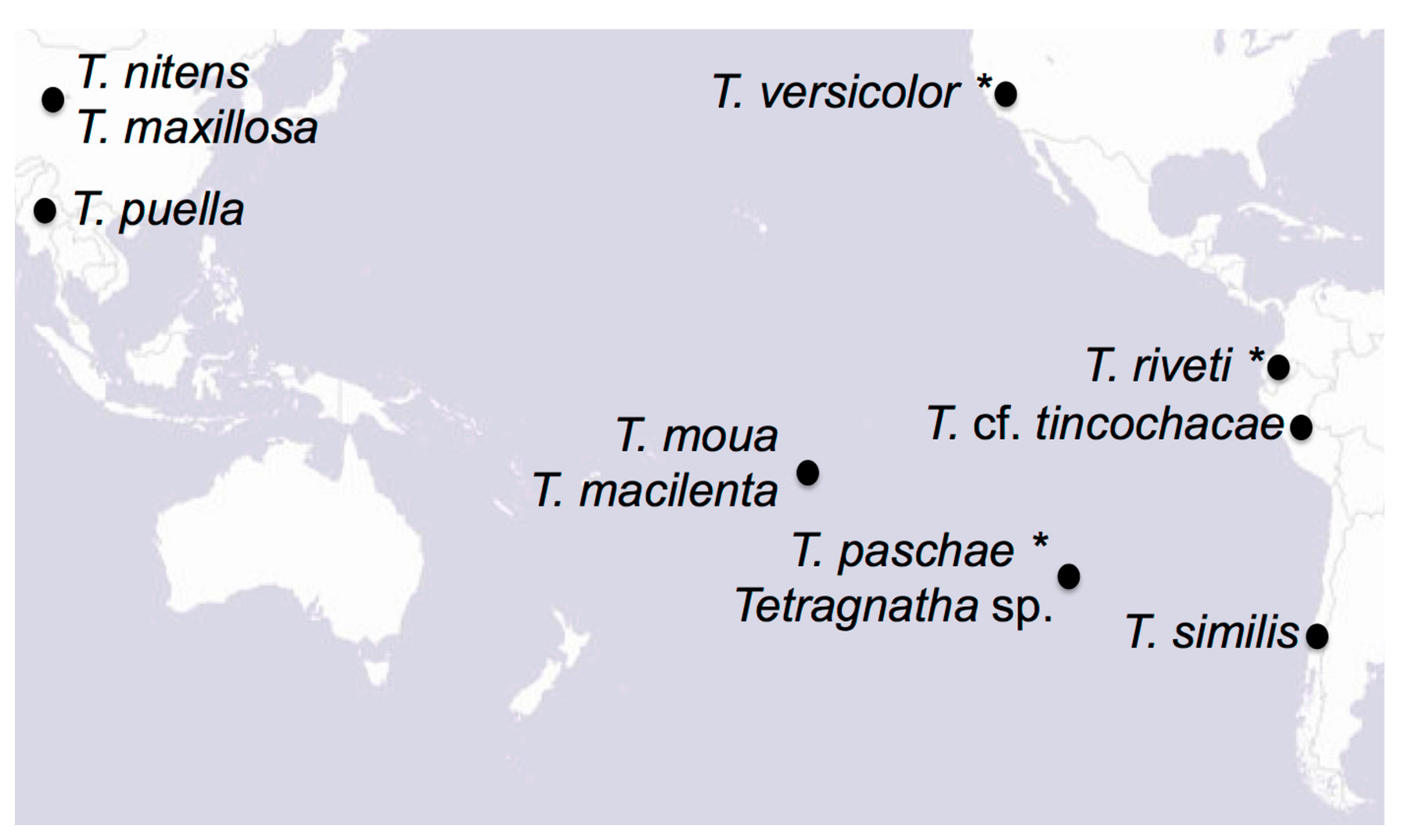
2.1. Samples
2.2. DNA Extraction
2.3. Library Preparation and Sequencing
2.4. Mitochondrial Genome Assembly
2.5. Phylogenetic Reconstruction
3. Results
3.1. DNA Recovery
3.2. Mitochondrial Genome Assemblies
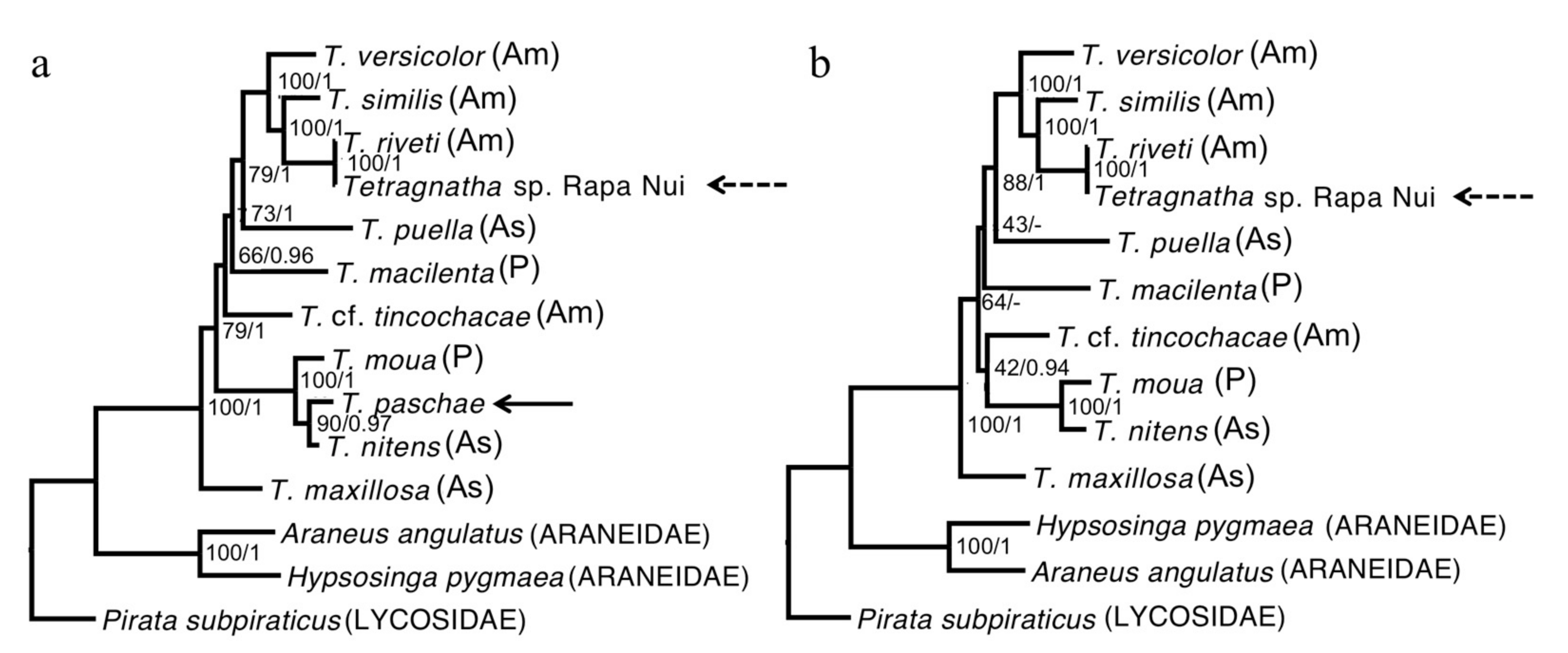
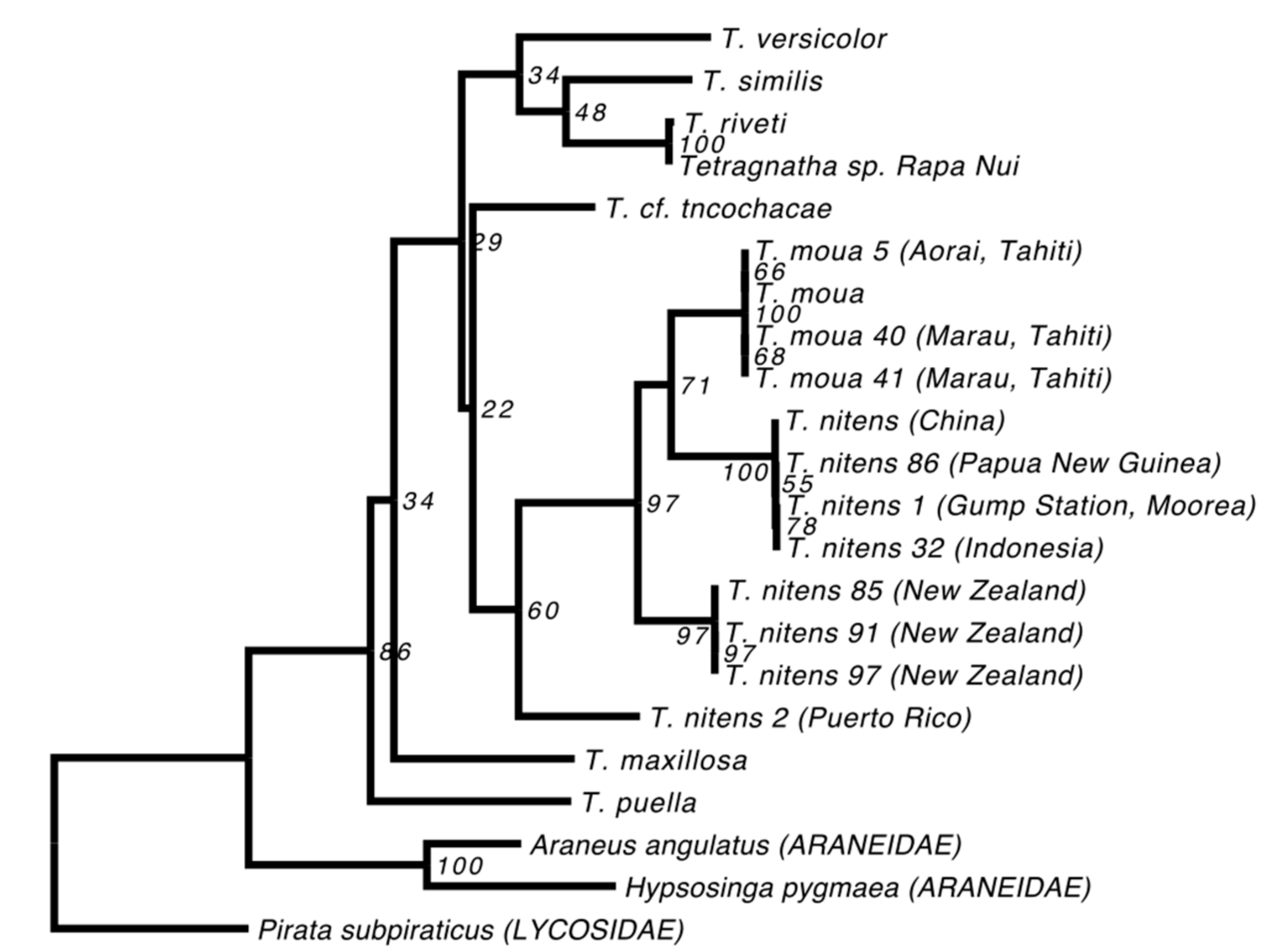
3.3. Phylogenetic Reconstruction
4. Discussion
4.1. Revealing the History of Tetragnatha Species on Rapa Nui
4.2. Recovering Ancient DNA from Museum-Preserved Spiders
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vezzoli, L.; Acocella, V. Easter Island, SE Pacific: An end-member type of hotspot volcanism. Bull. Geol. Soc. Am. 2009, 121, 869–886. [Google Scholar] [CrossRef]
- Azizi, G.; Flenley, J.R. The last glacial maximum climatic conditions on Easter Island. Quat. Int. 2008, 184, 166–176. [Google Scholar] [CrossRef]
- Wynne, J.J.; Bernard, E.C.; Howarth, F.G.; Sommer, S.; Soto-Adames, F.N.; Taiti, S.; Mockford, E.L.; Horrocks, M.; Pakarati, L.; Pakarati-Hotus, V. Disturbance relicts in a rapidly changing world: The rapa nui (Easter Island) factor. Bioscience 2014, 64, 711–718. [Google Scholar] [CrossRef]
- Hunt, T.L.; Lipo, C.P. Late Colonization of Easter Island. Science 2006, 311, 1603–1606. [Google Scholar] [CrossRef]
- Mann, D.; Edwards, J.; Chase, J.; Beck, W.; Reanier, R.; Mass, M.; Finney, B.; Loret, J. Drought, vegetation change, and human history on Rapa Nui (Isla de Pascua, Easter Island). Quat. Res. 2008, 69, 16–28. [Google Scholar] [CrossRef]
- Diamond, J. Collapse: How Societies Choose to Fail or Succeed. In Collapse: How Societies Choose to Fail or Succeed; Viking: New York, NY, USA, 2005. [Google Scholar]
- Fuentes, M.; Rovano, F.; Orellana, H. Restos industriales de la “Compañía explotadora de Isla de Pascua”. Una aproximación a la organización especial en Rapa Nui durante el ciclo ganadero (1895-1953). Rev. Atlántica-Mediterránea Prehist. y Arqueol. Soc. 2012, 9435. [Google Scholar]
- Berland, L. Araignées de l’île de Pâques et des îles Juan Fernandez. In The Natural History of Juan Fernandez and Easter Island; Skottsberg, C., Ed.; Almqvist & Wiksells Boktryckeri AB: Uppsala, Sweeden, 1924; pp. 419–437. [Google Scholar]
- Baert, L.; Lehtinen, P.; Desender, K. The spiders (Araneae) of Rapa Nui (Easter Island). Bull. Inst. R. Sci. Nat. Belg. Entomol. 1997, 67, 9–32. [Google Scholar]
- Aiken, M.; Coyle, F.A. Habitat Distribution, Life History and Behavior of Tetragnatha Spider Species in the Great Smoky Mountains National Park. J. Arachnol. 2000, 28, 97–106. [Google Scholar] [CrossRef]
- Gillespie, R.G. Biogeography of spiders on remote oceanic islands of the Pacific: Archipelagoes as stepping stones? J. Biogeogr. 2002, 29, 655–662. [Google Scholar] [CrossRef]
- Edwards, J.S.; Thornton, I.W.B. Colonization of an Island Volcano, Long Island, Papua New Guinea, and an Emergent Island, Motmot, in Its Caldera Lake. VI. The Pioneer Arthropod Community of Motmot. J. Biogeogr. 2016, 28, 1379–1388. [Google Scholar] [CrossRef]
- Gillespie, R.G. Community assembly through adaptive radiation in Hawaiian spiders. Science 2004, 303, 356–359. [Google Scholar] [CrossRef] [PubMed]
- Cotoras, D.D.; Wynne, J.J.; Flores-Prado, L.; Villagra, C. The spiders of Rapa Nui (Easter island) revisited. Bish. Mus. Occas. Pap. 2017, 120, 1–17. [Google Scholar]
- Miller, J.A.; Beentjes, K.K.; Van Helsdingen, P.; Ijland, S. Which specimens from a museum collection will yield DNA barcodes? A time series study of spiders in alcohol. Zookeys 2013, 365, 245–261. [Google Scholar] [CrossRef] [PubMed]
- Hofreiter, M.; Serre, D.; Poinar, H.N.; Kuch, M.; Pääbo, S. Ancient DNA. Nat. Rev. Genet. 2001, 2, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Alcohol Specifications European Union. Alcohol Specifications European Union. Available online: www.distill.com/specs/EU2.html (accessed on 27 November 2017).
- Gilbert, M.T.P.; Moore, W.; Melchior, L.; Worebey, M. DNA extraction from dry museum beetles without conferring external morphological damage. PLoS ONE 2007, 2, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, P.F.; Elias, S.; Gilbert, M.T.P.; Haile, J.; Munch, K.; Kuzmina, S.; Froese, D.G.; Sher, A.; Holdaway, R.N.; Willerslev, E. Non-destructive sampling of ancient insect DNA. PLoS ONE 2009, 4. [Google Scholar] [CrossRef] [PubMed]
- Heintzman, P.D.; Elias, S.A.; Moore, K.; Paszkiewicz, K.; Barnes, I. Characterizing DNA preservation in degraded specimens of Amara alpina (Carabidae: Coleoptera). Mol. Ecol. Resour. 2014, 14, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Tin, M.M.Y.; Economo, E.P.; Mikheyev, A.S. Sequencing degraded DNA from non-destructively sampled museum specimens for RAD-tagging and low-coverage shotgun phylogenetics. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Vink, C.J.; Thomas, S.M.; Paquin, P.; Hayashi, C.Y.; Hedin, M. The effects of preservatives and temperatures on arachnid DNA. Invertebr. Syst. 2005, 19, 99–104. [Google Scholar] [CrossRef]
- Krehenwinkel, H.; Tautz, D. Northern range expansion of European populations of the wasp spider Argiope bruennichi is associated with global warming-correlated genetic admixture and population-specific temperature adaptations. Mol. Ecol. 2013, 22, 2232–2248. [Google Scholar] [CrossRef] [PubMed]
- Dabney, J.; Knapp, M.; Glocke, I.; Gansauge, M.-T.; Weihmann, A.; Nickel, B.; Valdiosera, C.; Garcia, N.; Paabo, S.; Arsuaga, J.-L.; Meyer, M. Complete mitochondrial genome sequence of a Middle Pleistocene cave bear reconstructed from ultrashort DNA fragments. Proc. Natl. Acad. Sci. USA 2013, 110, 15758–15763. [Google Scholar] [CrossRef] [PubMed]
- Fulton, T. L. Setting Up an Ancient DNA Laboratory. In Ancient DNA Methods and Protocols; Shapiro, B., Hofreiter, M., Eds.; Humana Press: New York, NY, USA, 2011; pp. 1–11. [Google Scholar]
- Meyer, M.; Kircher, M. Illumina sequencing library preparation for highly multiplexed target capture and sequencing. Cold Spring Harb. Protoc. 2010, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- SeqPrep. https://github.com/jstjohn/SeqPrep (accessed on 18 December 2017).
- Mitochondrial Iterative Assembler. https://github.com/mpieva/mapping-iterative-assembler (accessed on 18 December 2017).
- Chang, D.; Knapp, M.; Enk, J.; Lippold, S.; Kircher, M.; Lister, A.; MacPhee, R.D.E.; Widga, C.; Czechowski, P.; Sommer, R.; Hodges, E.; et al. The evolutionary and phylogeographic history of woolly mammoths: a comprehensive mitogenomic analysis. Sci. Rep. 2017, 7, 44585. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. The RAxML v8.2.X Manual. Manual 2014, 1–55. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [PubMed]
- Tracer. http://beast.bio.ed.ac.uk/Tracer (accessed on 18 December 2017).
- World Spider Catalog. Natural History Museum Bern. Version 18.5. Available online: http://wsc.nmbe.ch (accessed on 9 August 2017).
- Fig Tree. http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 18 December 2017).
- Kuschel, G. Composition and relationship of the terrestrial faunas of Easter, Juan Fernandez, Desventuradas and Galápagos islands. Occas. Pap. Calif. Acad. Sci. 1963, 79–95. [Google Scholar] [CrossRef]
- Dransfield, J.; Flenley, J.R.; King, S.M.; Harkness, D.D.; Rapu, S. A recently extinct palm from Easter Island. Nature 1984, 312, 750–752. [Google Scholar] [CrossRef]
- Berland, L. Araignées. In Mission du Service Géographique de l’armée Pour la Mesure d’un arc du Méridien équatorial en Amérique du Sud (1899–1906); Gauthier-Villars, Imprimeur Libraire du Bureau des Longitudes, de l’Ecole Polytechnique: Paris, France, 1913; pp. 78–119. [Google Scholar]
- Gillespie, R.G.; Claridge, E.M.; Goodacre, S.L. Biogeography of the fauna of French Polynesia: Diversification within and between a series of hot spot archipelagos. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 3335–3346. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, R.G. Spiders of the Genus Tetragnatha (Araneae, Tetragnathidae) in the Society Islands. J. Arachnol. 2003, 31, 157–172. [Google Scholar] [CrossRef]
- Okuma, C. Notes on the neotropical and Mexican species of Tetragnatha (Araneae: Tetragnathidae) with descriptions of three new species. J. Fac. Agric. Kyushu Univ. 1992, 36, 219–243. [Google Scholar]
- The IUCN Red List of Threatened Species. Version 2017-1. Available online: www.iucnredlist.org.
- Gerlach, J.; Marusik, Y. Arachnida and Myriapoda of the Seychelles Islands; Siri Scientific Press: Manchester, UK, 2010; ISBN 978-0-9558636-8-4. [Google Scholar]
- Shochat, A.E.; Stefanov, W.L.; Whitehouse, M.E.; Faeth, S.H. Urbanization and Spider Diversity : Influences of Human Modification of Habitat Structure and Productivity. Urb. Ecol. 2014, 14, 268–280. [Google Scholar]
- Chapco, W.; Litzenberger, G. A DNA investigation into the mysterious disappearance of the Rocky Mountain grasshopper, mega-pest of the 1800s. Mol. Phylogenet. Evol. 2004, 30, 810–814. [Google Scholar] [CrossRef]
- Brewer, M.S.; Cotoras, D.D.; Croucher, P.J.P.; Gillespie, R. G. New sequencing technologies, the development of genomics tools, and their applications in evolutionary arachnology. J. Arachnol. 2014, 42, 1–15. [Google Scholar] [CrossRef]
- Rohland, N.; Hofreiter, M. Comparison and optimization of ancient DNA extraction. Biotechniques 2007, 42, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Campos, P.F.; Craig, O.E.; Turner-Walker, G.; Peacock, E.; Willerslev, E.; Gilbert, M.T.P. DNA in ancient bone—Where is it located and how should we extract it? Ann. Anat. 2012, 194, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Stevens, R.; Stevens, L.; Price, N. The stabilities of various thiol compounds used in protein purifications. Biochem. Educ. 1983, 11, 70. [Google Scholar] [CrossRef]
- Bio-Rad Technical support FAQ. Available online: www.bio-rad.com/en-us/faq/268440261/technical-support-faq (accessed on 22 May 2017).
- Sanggaard, K.W.; Bechsgaard, J.S.; Fang, X.; Duan, J.; Dyrlund, T.F.; Gupta, V.; Jiang, X.; Cheng, L.; Fan, D.; Feng, Y.; et al. Spider genomes provide insight into composition and evolution of venom and silk. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef]
- Babb, P. L.; Lahens, N.F.; Correa-Garhwal, S.M.; Nicholson, D.N.; Kim, E.J.; Hogenesch, J.B.; Kuntner, M.; Higgins, L.; Hayashi, C.Y.; Agnarsson, I.; Voight, B.F. The Nephila clavipes genome highlights the diversity of spider silk genes and their complex expression. Nat. Genet. 2017, 49, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Schwager, E.E.; Sharma, P.P.; Clarke, T.; Leite, D.J.; Wierschin, T.; Pechmann, M.; Akiyama-Oda, Y.; Esposito, L.; Bechsgaard, J.; Bilde, T.; et al. The house spider genome reveals an ancient whole-genome duplication during arachnid evolution. BMC Biol. 2017, 15, 62. [Google Scholar] [CrossRef] [PubMed]
- Garrison, N.L.; Rodriguez, J.; Agnarsson, I.; Coddington, J. A.; Griswold, C.E.; Hamilton, C.A.; Hedin, M.; Kocot, K.M.; Ledford, J.M.; Bond, J.E. Spider phylogenomics: Untangling the Spider Tree of Life. PeerJ 2016, 4, e1719. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, S.; McPherson, J.D.; McCombie, W.R. Coming of age: Ten years of next-generation sequencing technologies. Nat. Rev. Genet. 2016, 17, 333–351. [Google Scholar] [CrossRef] [PubMed]
- Putnam, N.H.; Connell, B.O.; Stites, J.C.; Rice, B.J.; Hartley, P.D.; Sugnet, C.W.; Haussler, D.; Rokhsar, D.S. Chromosome-scale shotgun assembly using an in vitro method for long-range linkage. Genome Res. 2016, 26, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Sohn, J.; Nam, J.-W. The present and future of de novo whole-genome assembly. Brief. Bioinform. 2016. [Google Scholar] [CrossRef] [PubMed]
- Prebble, M.; Whitau, R.; Meyer, J.Y.; Sibley-Punnett, L.; Fallon, S.; Porch, N. Abrupt late Pleistocene ecological and climate change on Tahiti (French Polynesia). J. Biogeogr. 2016, 43, 2438–2453. [Google Scholar] [CrossRef]
- Liebherr, J.K.; Porch, N. Reassembling a lost lowland carabid beetle assemblage (Coleoptera) from Kauai, Hawaiian Islands. Invertebr. Syst. 2015, 29, 191–213. [Google Scholar] [CrossRef]
- Reiss, R.A. Ancient DNA from ice age insects: Proceed with caution. Quat. Sci. Rev. 2006, 25, 1877–1893. [Google Scholar] [CrossRef]
- King, G.A.; Gilbert, M.T.P.; Willerslev, E.; Collins, M.J.; Kenward, H. Recovery of DNA from archaeological insect remains: First results, problems and potential. J. Archaeol. Sci. 2009, 36, 1179–1183. [Google Scholar] [CrossRef]

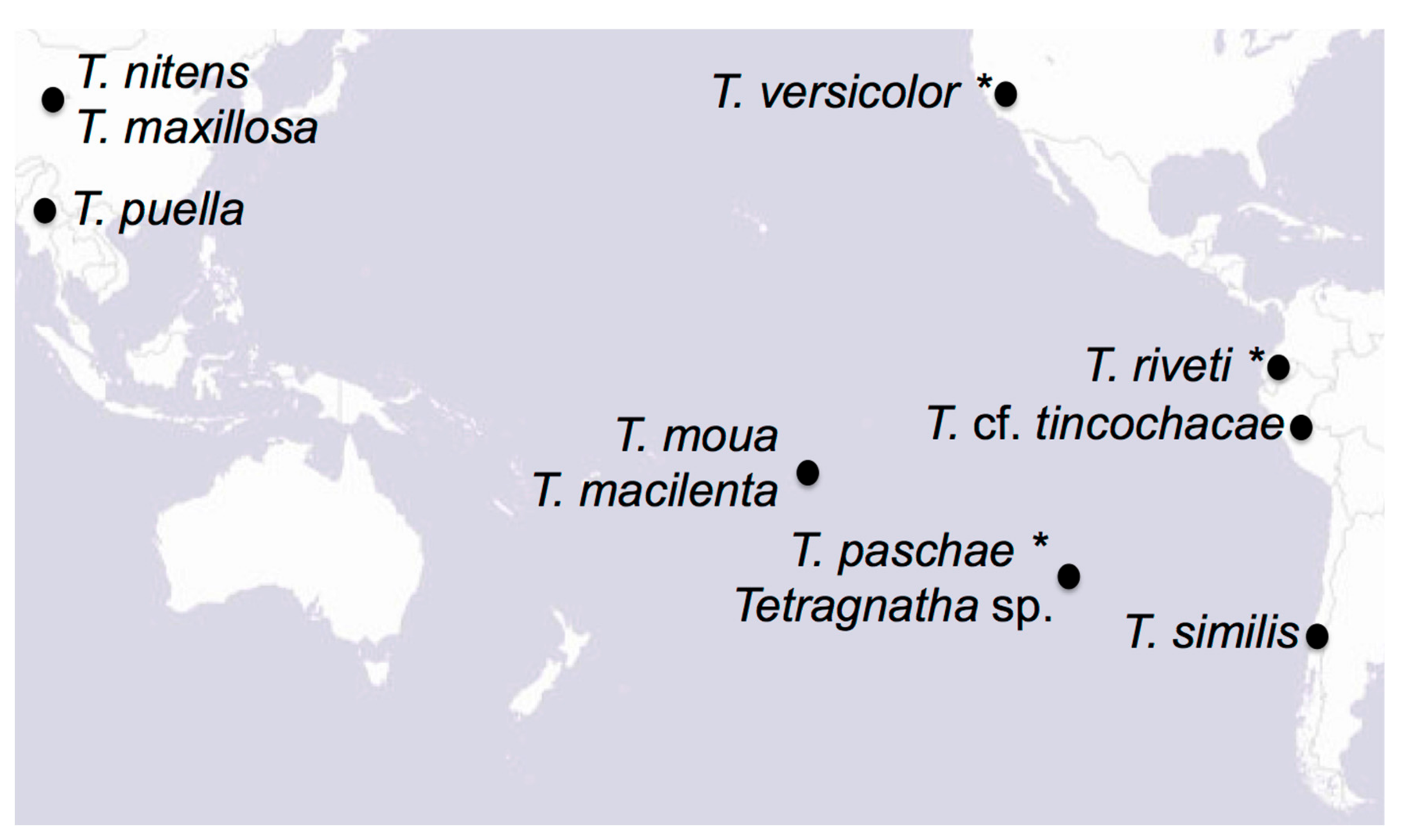{kind=link}
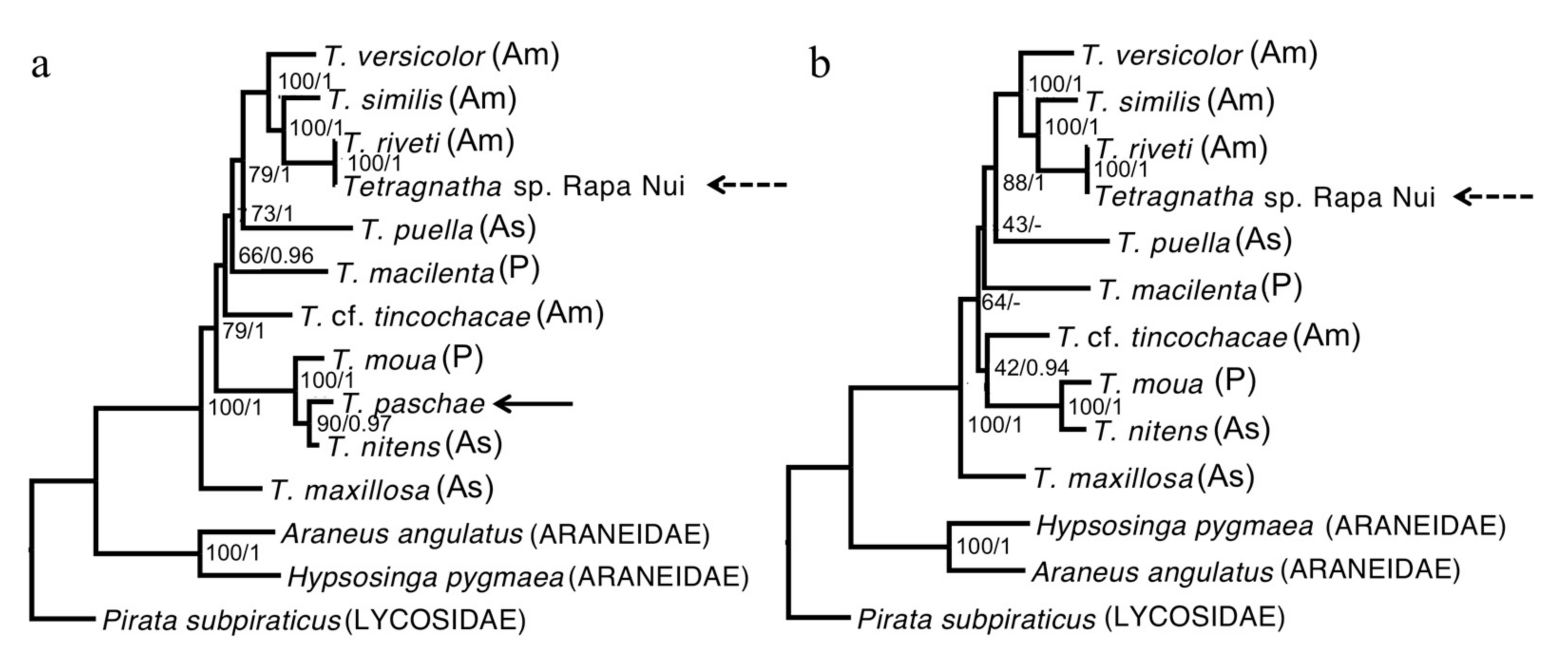{kind=link}
{kind=link}
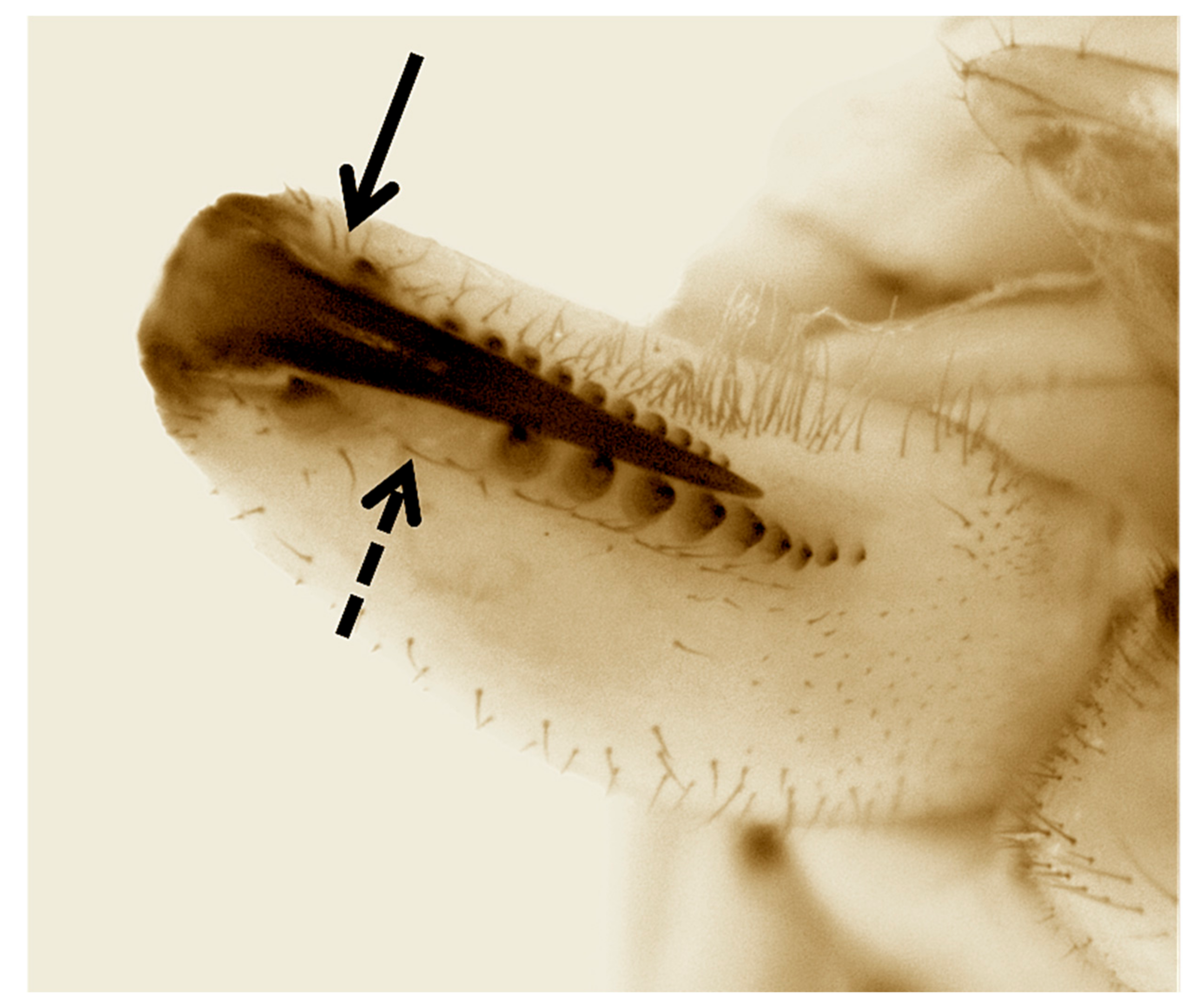{kind=link}
| Species Name | Geographic Origin | Collection/GenBank | Preservation Conditions | Collection Year |
|---|---|---|---|---|
| Tetragnatha paschae | Rapa Nui | Natural History Museum of London | Ethanol 70% originally and currently 80% IDA (industrial denatured alcohol), room temperature | 1925 |
| Tetragnatha riveti | Ecuador | California Academy of Sciences | Ethanol 70%, room temperature | 1943 |
| Tetragnatha versicolor * | United States (California) | California Academy of Sciences | Ethanol 70%, room temperature | 1947 |
| Tetragnatha puella | Myanmar | California Academy of Sciences | Ethanol 95%, −20 °C | 2003 |
| Tetragnatha cf. tincochacae | Perú | California Academy of Sciences | Ethanol 95%, −20 °C | 2010 |
| Tetragnatha sp. | Rapa Nui | This study | Ethanol 95%, −20 °C | 2012 |
| Tetragnatha similis | Chile | This study | Ethanol 95%, −20 °C | 2015 |
| Tetragnatha macilenta | Tahiti | This study | Ethanol 95%, −20 °C | 2015 |
| Tetragnatha moua | Tahiti | This study | Ethanol 95%, −20 °C | 2015 |
| Tetragnatha maxillosa | China | KP306789.1 | - | - |
| Tetragnatha nitens | China | KP306790.1 | - | - |
| Hypsosinga pygmaea | China | NC028078.1 | - | - |
| Araneus angulatus | China | KU365988.1 | - | - |
| Pirata subpiraticus | China | KM486623.1 | - | - |
| Species Name | Merged Reads | Unmerged Read Pairs | Mapped Reads | % Mapped Reads | Coverage * |
|---|---|---|---|---|---|
| T. paschae | 2,035,294 | 127,651 | 274 | 0.012 | 1.142 |
| T. riveti | 1,152,000 | 191,722 | 1717 | 0.112 | 7.754 |
| T. versicolor | 1,664,113 | 81,418 | 19,793 | 1.08 | 96.616 |
| T. puella | 203,936 | 464,142 | 4695 | 0.415 | 27.776 |
| T. cf. tincochacae | 205,455 | 501,781 | 2811 | 0.232 | 16.923 |
| Tetragnatha sp. Rapa Nui | 187,032 | 411,508 | 2944 | 0.291 | 17.622 |
| T. similis | 109,240 | 580,116 | 2003 | 0.158 | 11.481 |
| T. macilenta | 159,505 | 729,383 | 2023 | 0.125 | 11.866 |
| T. moua | 307,068 | 732,206 | 3663 | 0.207 | 21.920 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cotoras, D.D.; Murray, G.G.R.; Kapp, J.; Gillespie, R.G.; Griswold, C.; Simison, W.B.; Green, R.E.; Shapiro, B. Ancient DNA Resolves the History of Tetragnatha (Araneae, Tetragnathidae) Spiders on Rapa Nui. Genes 2017, 8, 403. https://doi.org/10.3390/genes8120403
Cotoras DD, Murray GGR, Kapp J, Gillespie RG, Griswold C, Simison WB, Green RE, Shapiro B. Ancient DNA Resolves the History of Tetragnatha (Araneae, Tetragnathidae) Spiders on Rapa Nui. Genes. 2017; 8(12):403. https://doi.org/10.3390/genes8120403
Chicago/Turabian StyleCotoras, Darko D., Gemma G. R. Murray, Joshua Kapp, Rosemary G. Gillespie, Charles Griswold, W. Brian Simison, Richard E. Green, and Beth Shapiro. 2017. "Ancient DNA Resolves the History of Tetragnatha (Araneae, Tetragnathidae) Spiders on Rapa Nui" Genes 8, no. 12: 403. https://doi.org/10.3390/genes8120403
APA StyleCotoras, D. D., Murray, G. G. R., Kapp, J., Gillespie, R. G., Griswold, C., Simison, W. B., Green, R. E., & Shapiro, B. (2017). Ancient DNA Resolves the History of Tetragnatha (Araneae, Tetragnathidae) Spiders on Rapa Nui. Genes, 8(12), 403. https://doi.org/10.3390/genes8120403