
Noncoding RNA Profiles in Tobacco- and Alcohol-Associated Diseases

Abstract
:1. Introduction
2. Noncoding RNAs
2.1. miRNAs
2.2. lncRNAs
3. Diseases and Smoking: An Introduction
3.1. miRNAs in Tobacco-Associated Diseases
3.1.1. Lung Cancer
3.1.2. Other Cancers
3.1.3. Chronic Obstructive Pulmonary Disease
3.1.4. Cardiovascular Diseases
4. Alcohol Abuse and Alcohol-Related Diseases
4.1. miRNAs in Alcohol-Associated Diseases
4.1.1. Liver Cancer
4.1.2. Other Cancers
4.1.3. Other Alcoholic Liver Diseases
5. Long Noncoding RNAs in Tobacco and Alcohol-Associated Diseases
6. Noncoding RNAs Involved in Signaling Pathways
7. Final Considerations
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Health Organization. Global Information System on Alcohol and Health (GISAH). Available online: http://www.who.int/gho/alcohol/en/ (accessed on 20 November 2016).
- Zhou, Y.; Zheng, J.; Li, S.; Zhou, T.; Zhang, P.; Li, H.B. Alcoholic Beverage Consumption and Chronic Diseases. Int. J. Environ. Res. Public Health 2016, 13, 522. [Google Scholar] [CrossRef] [PubMed]
- Leyro, T.M.; Crew, E.E.; Bryson, S.W.; Lembke, A.; Bailey, S.R.; Prochaska, J.J.; Henriksen, L.; Fortmann, S.P.; Killen, J.D.; Killen, D.T.; et al. Retrospective analysis of changing characteristics of treatment-seeking smokers: Implications for further reducing smoking prevalence. BMJ Open 2016, 6, e010960. [Google Scholar] [CrossRef] [PubMed]
- Mathers, C.D.; Loncar, D. Projections of global mortality and burden of disease from 2002 to 2030. PLoS Med. 2006, 3, e442. [Google Scholar] [CrossRef] [PubMed]
- Parkin, D.M.; Boyd, L.; Walker, L.C. The fraction of cancer attributable to lifestyle and environmental factors in the UK in 2010. Br. J. Cancer 2011, 105 (Suppl. S2), S77–S81. [Google Scholar] [CrossRef] [PubMed]
- Alegría-Torres, J.A.; Baccarelli, A.; Bollati, V. Epigenetics and lifestyle. Epigenomics 2011, 3, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Luo, F.; Xu, Y.; Wang, B.; Zhao, Y.; Xu, W.; Shi, L.; Lu, X.; Liu, Q. Epithelial-mesenchymal transition and cancer stem cells, mediated by a long noncoding RNA, HOTAIR, are involved in cell malignant transformation induced by cigarette smoke extract. Toxicol. Appl. Pharmacol. 2015, 282, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Lin, Y.; Zhan, M.; Mann, D.L.; Stass, S.A.; Jiang, F. Differential miRNA expressions in peripheral blood mononuclear cells for diagnosis of lung cancer. Lab. Investig. 2015, 95, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Francis, H.; McDaniel, K.; Han, Y.; Liu, X.; Kennedy, L.; Yang, F.; McCarra, J.; Zhou, T.; Glaser, S.; Venter, J.; et al. Regulation of the extrinsic apoptotic pathway by microRNA-21 in alcoholic liver injury. J. Biol. Chem. 2014, 289, 27526–27539. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Wei, W.; Chen, L.; Xia, Z.; Yu, C. Comprehensive analysis of long noncoding RNA expression profiles in hepatitis B virus-related hepatocellular carcinoma. Oncotarget 2016. [Google Scholar] [CrossRef] [PubMed]
- Vucic, E.A.; Thu, K.L.; Pikor, L.A.; Enfield, K.S.; Yee, J.; English, J.C.; MacAulay, C.E.; Lam, S.; Jurisica, I.; Lam, W.L. Smoking status impacts microRNA mediated prognosis and lung adenocarcinoma biology. BMC Cancer 2014, 14, 778. [Google Scholar] [CrossRef] [PubMed]
- Geng, Q.; Fan, T.; Zhang, B.; Wang, W.; Xu, Y.; Hu, H. Five microRNAs in plasma as novel biomarkers for screening of early-stage non-small cell lung cancer. Respir. Res. 2014, 15, 149. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Luo, F.; Wang, B.; Li, H.; Xu, Y.; Liu, X.; Shi, L.; Lu, X.; Xu, W.; Lu, L.; et al. STAT3-regulated exosomal miR-21 promotes angiogenesis and is involved in neoplastic processes of transformed human bronchial epithelial cells. Cancer Lett. 2016, 370, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Kashi, K.; Henderson, L.; Bonetti, A.; Carninci, P. Discovery and functional analysis of lncRNAs: Methodologies to investigate an uncharacterized transcriptome. Biochim. Biophys. Acta 2016, 1859, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, I.; Ili, C.; Roa, J.C.; Brebi, P. Long noncoding RNAs in gastric cancer: Mechanisms and potential applications. Oncotarget 2016. [Google Scholar] [CrossRef]
- Hafner, M.; Lianoglou, S.; Tuschl, T.; Betel, D. Genome-wide identification of miRNA targets by PAR-CLIP. Methods 2012, 58, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef] [PubMed]
- Kunej, T.; Godnic, I.; Horvat, S.; Zorc, M.; Calin, G.A. Cross talk between microRNA and coding cancer genes. Cancer J. 2012, 18, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Eiring, A.M.; Harb, J.G.; Neviani, P.; Garton, C.; Oaks, J.J.; Spizzo, R.; Liu, S.; Schwind, S.; Santhanam, R.; Hickey, C.J.; et al. miR-328 functions as an RNA decoy to modulate hnRNP E2 regulation of mRNA translation in leukemic blasts. Cell 2010, 140, 652–665. [Google Scholar] [CrossRef] [PubMed]
- Garzon, R.; Marcucci, G.; Croce, C.M. Targeting microRNAs in cancer: Rationale, strategies and challenges. Nat. Rev. Drug Discov. 2010, 9, 775–789. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, M.; Paone, A.; Calore, F.; Galli, R.; Gaudio, E.; Santhanam, R.; Lovat, F.; Fadda, P.; Mao, C.; Nuovo, G.J.; et al. MicroRNAs bind to Toll-like receptors to induce prometastatic inflammatory response. Proc. Natl. Acad. Sci. USA 2012, 109, E2110–E2116. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, M.; Garzon, R.; Cimmino, A.; Liu, Z.; Zanesi, N.; Callegari, E.; Liu, S.; Alder, H.; Costinean, S.; Fernandez-Cymering, C.; et al. MicroRNA-29 family reverts aberrant methylation in lung cancer by targeting DNA methyltransferases 3A and 3B. Proc. Natl. Acad. Sci. USA 2007, 104, 15805–15810. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, S.; Tong, Y.; Steitz, J.A. Switching from repression to activation: MicroRNAs can up-regulate translation. Science 2007, 318, 1931–1934. [Google Scholar] [CrossRef] [PubMed]
- Endzeliņš, E.; Melne, V.; Kalniņa, Z.; Lietuvietis, V.; Riekstiņa, U.; Llorente, A.; Linē, A. Diagnostic, prognostic and predictive value of cell-free miRNAs in prostate cancer: A systematic review. Mol. Cancer 2016, 15, 41. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.Y.; Calin, G.A. MicroRNAs as therapeutic targets in human cancers. Wiley Interdiscip. Rev. RNA 2014, 5, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Noncoding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Tuna, M.; Machado, A.S.; Calin, G.A. Genetic and epigenetic alterations of microRNAs and implications for human cancers and other diseases. Genes Chromosomes Cancer 2016, 55, 193–214. [Google Scholar] [CrossRef] [PubMed]
- Borchert, G.M.; Lanier, W.; Davidson, B.L. RNA polymerase III transcribes human microRNAs. Nat. Struct. Mol. Biol. 2006, 13, 1097–1101. [Google Scholar] [CrossRef] [PubMed]
- Gurtner, A.; Falcone, E.; Garibaldi, F.; Piaggio, G. Dysregulation of microRNA biogenesis in cancer: The impact of mutant p53 on Drosha complex activity. J. Exp. Clin. Cancer Res. 2016, 35, 45. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Glaser, S.S.; Francis, H.; Yang, F.; Han, Y.; Stokes, A.; Staloch, D.; McCarra, J.; Liu, J.; Venter, J.; et al. Epigenetic regulation of miR-34a expression in alcoholic liver injury. Am. J. Pathol. 2012, 181, 804–817. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, N.; Ojima, H.; Shirakihara, T.; Shimizu, H.; Kokubu, A.; Urushidate, T.; Totoki, Y.; Kosuge, T.; Miyagawa, S.; Shibata, T. Downregulation of the microRNA biogenesis components and its association with poor prognosis in hepatocellular carcinoma. Cancer Sci. 2013, 104, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.H.; Hsieh, M.J.; Chiou, H.L.; Liou, Y.S.; Yang, C.C.; Yang, S.F.; Kuo, W.H. MicroRNA gene polymorphisms and environmental factors increase patient susceptibility to hepatocellular carcinoma. PLoS ONE 2014, 9, e89930. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Ivan, C.; Yang, D.; Gharpure, K.M.; Wu, S.Y.; Pecot, C.V.; Previs, R.A.; Nagaraja, A.S.; Armaiz-Pena, G.N.; McGuire, M.; et al. Hypoxia-upregulated microRNA-630 targets Dicer, leading to increased tumor progression. Oncogene 2016, 35, 4312–4320. [Google Scholar] [CrossRef] [PubMed]
- Gross, T.J.; Powers, L.S.; Boudreau, R.L.; Brink, B.; Reisetter, A.; Goel, K.; Gerke, A.K.; Hassan, I.H.; Monick, M.M. A microRNA processing defect in smokers’ macrophages is linked to SUMOylation of the endonuclease DICER. J. Biol. Chem. 2014, 289, 12823–12834. [Google Scholar] [CrossRef] [PubMed]
- Geisler, S.; Coller, J. RNA in unexpected places: Long non-coding RNA functions in diverse cellular contexts. Nat. Rev. Mol. Cell Biol. 2013, 14, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Loewer, S.; Cabili, M.N.; Guttman, M.; Loh, Y.H.; Thomas, K.; Park, I.H.; Garber, M.; Curran, M.; Onder, T.; Agarwal, S.; et al. Large intergenic non-coding RNA-RoR modulates reprogramming of human induced pluripotent stem cells. Nat. Genet. 2010, 42, 1113–1117. [Google Scholar] [CrossRef] [PubMed]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure; evolution; and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, S.; Kageyama, Y. Nuclear lncRNAs as epigenetic regulators—Beyond skepticism. Biochim. Biophys. Acta 2014, 1839, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Heward, J.A.; Lindsay, M.A. Long non-coding RNAs in the regulation of the immune response. Trends Immunol. 2014, 35, 408–419. [Google Scholar] [CrossRef] [PubMed]
- Wan, P.; Su, W.; Zhuo, Y. The Role of Long Noncoding RNAs in Neurodegenerative Diseases. Mol. Neurobiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Archer, K.; Broskova, Z.; Bayoumi, A.S.; Teoh, J.P.; Davila, A.; Tang, Y.; Su, H.; Kim, I.M. Long Non-Coding RNAs as Master Regulators in Cardiovascular Diseases. Int. J. Mol. Sci. 2015, 16, 23651–23667. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wong, D. Long non-coding RNA-mediated regulation of glucose homeostasis and diabetes. Am. J. Cardiovasc. Dis. 2016, 18, 17–25. [Google Scholar]
- Thai, P.; Statt, S.; Chen, C.H.; Liang, E.; Campbell, C.; Wu, R. Characterization of a novel long noncoding RNA; SCAL1; induced by cigarette smoke and elevated in lung cancer cell lines. Am. J. Respir. Cell Mol. Biol. 2013, 49, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.C.; Yang, Z.; Zhou, L.; Zhu, Q.Q.; Xie, H.Y.; Zhang, F.; Wu, L.M.; Chen, L.M.; Zheng, S.S. Long noncoding RNA MALAT-1 overexpression predicts tumor recurrence of hepatocellular carcinoma after liver transplantation. Med. Oncol. 2012, 29, 1810–1816. [Google Scholar] [CrossRef] [PubMed]
- Doll, R.; Hill, A.B. Smoking and carcinoma of the lung; preliminary report. Br. Med. J. 1950, 2, 739–748. [Google Scholar]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Personal habits and indoor combustions. Volume 100 E. A review of human carcinogens. IARC Monogr. Eval. Carcinog. Risks Hum. 2012, 100E, 1–598. [Google Scholar]
- U.S. Department of Health and Human Services. Fifty Years of Change 1964–2014. In The Health Consequences of Smoking: 50 Years of Progress; A Report of the Surgeon General; U.S. Department of Health and Human Services; Centers for Disease Control and Prevention; National Center for Chronic Disease Prevention and Health Promotion; Office on Smoking and Health: Atlanta, GA, USA, 2014; pp. 15–41. [Google Scholar]
- Gritz, E.R.; Fingeret, M.C.; Vidrine, D.J.; Lazev, A.B.; Mehta, N.V.; Reece, G.P. Successes and failures of the teachable moment: Smoking cessation in cancer patients. Cancer 2006, 106, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, R.M. Exposure to tobacco smoke causes immediate damage: A report of the Surgeon General. Public Health Rep. 2011, 126, 158–159. [Google Scholar] [PubMed]
- Wang, L.; Li, X.; Zhou, Y.; Shi, H.; Xu, C.; He, H.; Wang, S.; Xiong, X.; Zhang, Y.; Du, Z.; et al. Downregulation of miR-133 via MAPK/ERK signaling pathway involved in nicotine-induced cardiomyocyte apoptosis. Naunyn-Schmiedeberg Arch. Pharmacol. 2014, 387, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Maegdefessel, L.; Azuma, J.; Toh, R.; Deng, A.; Merk, D.R.; Raiesdana, A.; Leeper, N.J.; Raaz, U.; Schoelmerich, A.M.; McConnell, M.V.; et al. MicroRNA-21 blocks abdominal aortic aneurysm development and nicotine-augmented expansion. Sci. Transl. Med. 2012, 4, 122ra22. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, R.; Strulovici-Barel, Y.; Salit, J.; Staudt, M.R.; Ahmed, J.; Tilley, A.E.; Yee-Levin, J.; Hollmann, C.; Harvey, B.G.; et al. Persistence of smoking-induced dysregulation of miRNA expression in the small airway epithelium despite smoking cessation. PLoS ONE 2015, 10, e0120824. [Google Scholar] [CrossRef] [PubMed]
- Roth, C.; Stückrath, I.; Pantel, K.; Izbicki, J.R.; Tachezy, M.; Schwarzenbach, H. Low levels of cell-free circulating miR-361-3p and miR-625* as blood-based markers for discriminating malignant from benign lung tumors. PLoS ONE 2012, 7, e38248. [Google Scholar] [CrossRef] [PubMed]
- American Cancer Society (ACS). Cancer Facts & Figures 2012. J. Consum. Health Internet 2012, 16, 366–367. [Google Scholar]
- Su, M.W.; Yu, S.L.; Lin, W.C.; Tsai, C.H.; Chen, P.H.; Lee, Y.L. Smoking-related microRNAs and mRNAs in human peripheral blood mononuclear cells. Toxicol. Appl. Pharmacol. 2016, 305, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Izzotti, A.; Balansky, R.; Ganchev, G.; Iltcheva, M.; Longobardi, M.; Pulliero, A.; Geretto, M.; Micale, R.T.; La Maestra, S.; Miller, M.S.; et al. Blood and lung microRNAs as biomarkers of pulmonary tumorigenesis in cigarette smoke-exposed mice. Oncotarget 2016. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wu, J.; Li, Y.; Li, X.; Yang, T.; Yang, Q.; Jiang, Y. Deregulation of serum microRNA expression is associated with cigarette smoking and lung cancer. BioMed Res. Int. 2014, 2014, 364316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.K.; Zhu, W.Y.; He, J.Y.; Chen, D.D.; Huang, Y.Y.; Le, H.B.; Liu, X.G. miRNAs expression profiling to distinguish lung squamous-cell carcinoma from adenocarcinoma subtypes. J. Cancer Res. Clin. Oncol. 2012, 138, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.S.; Jeon, H.S.; Sun, Z.; Aubry, M.C.; Tang, H.; Park, C.H.; Rakhshan, F.; Schultz, D.A.; Kolbert, C.P.; Lupu, R.; et al. Increased miR-708 expression in NSCLC and its association with poor survival in lung adenocarcinoma from never smokers. Clin. Cancer Res. 2012, 18, 3658–3667. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.C.; Jung, Y.; Park, J.; Cho, S.; Seo, C.; Kim, J.; Kim, P.; Park, J.; Seo, J.; Kim, J.; et al. A high-dimensional; deep-sequencing study of lung adenocarcinoma in female never-smokers. PLoS ONE 2013, 8, e55596. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Zhang, H.; Su, Y.; Kong, J.; Yu, H.; Qian, B. Up-regulation of microRNA-183-3p is a potent prognostic marker for lung adenocarcinoma of female non-smokers. Clin. Transl. Oncol. 2014, 16, 980–985. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Liu, Z.; Todd, N.W.; Zhang, H.; Liao, J.; Yu, L.; Guarnera, M.A.; Li, R.; Cai, L.; Zhan, M.; et al. Diagnosis of lung cancer in individuals with solitary pulmonary nodules by plasma microRNA biomarkers. BMC Cancer 2011, 11, 374. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Liao, J.; Guarnera, M.A.; Fang, H.; Cai, L.; Stass, S.A.; Jiang, F. Analysis of microRNAs in sputum to improve computed tomography for lung cancer diagnosis. J. Thorac. Oncol. 2014, 9, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Sozzi, G.; Boeri, M.; Rossi, M.; Verri, C.; Suatoni, P.; Bravi, F.; Roz, L.; Conte, D.; Grassi, M.; Sverzellati, N.; et al. Clinical utility of a plasma-based miRNA signature classifier within computed tomography lung cancer screening: A correlative MILD trial study. J. Clin. Oncol. 2014, 32, 768–773. [Google Scholar] [CrossRef] [PubMed]
- Wali, R.K.; Hensing, T.A.; Ray, D.W.; Dela Cruz, M.; Tiwari, A.K.; Radosevich, A.; Jepeal, L.; Fernando, H.C.; Litle, V.R.; Charlot, M.; et al. Buccal microRNA dysregulation in lung field carcinogenesis: Gender-specific implications. Int. J. Oncol. 2014, 45, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Perdomo, C.; Campbell, J.D.; Gerrein, J.; Tellez, C.S.; Garrison, C.B.; Walser, T.C.; Drizik, E.; Si, H.; Gower, A.C.; Vick, J.; et al. MicroRNA 4423 is a primate-specific regulator of airway epithelial cell differentiation and lung carcinogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, 18946–18951. [Google Scholar] [CrossRef] [PubMed]
- Balansky, R.; Izzotti, A.; D’Agostini, F.; Longobardi, M.; Micale, R.T.; La Maestra, S.; Camoirano, A.; Ganchev, G.; Iltcheva, M.; Steele, V.E.; et al. Assay of lapatinib in murine models of cigarette smoke carcinogenesis. Carcinogenesis 2014, 35, 2300–2307. [Google Scholar] [CrossRef] [PubMed]
- Xi, S.; Inchauste, S.; Guo, H.; Shan, J.; Xiao, Z.; Xu, H.; Miettenen, M.; Zhang, M.R.; Hong, J.A.; Raiji, M.T.; et al. Cigarette smoke mediates epigenetic repression of miR-217 during esophageal adenocarcinogenesis. Oncogene 2015, 34, 5548–5559. [Google Scholar] [CrossRef] [PubMed]
- Stánitz, E.; Juhász, K.; Tóth, C.; Gombos, K.; Natali, P.G.; Ember, I. Evaluation of microRNA expression pattern of gastric adenocarcinoma associated with socioeconomic, environmental and lifestyle factors in northwestern Hungary. Anticancer Res. 2013, 33, 3195–3200. [Google Scholar] [PubMed]
- Kolokythas, A.; Zhou, Y.; Schwartz, J.L.; Adami, G.R. Similar Squamous Cell Carcinoma Epithelium microRNA Expression in Never Smokers and Ever Smokers. PLoS ONE 2015, 10, e0141695. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, L.; Sevinç, K.; Papazoglou, I.M.; Tildy, B.; Detillieux, K.; Halayko, A.J.; Chung, K.F.; Perry, M.M. Airway smooth muscle inflammation is regulated by microRNA-145 in COPD. FEBS Lett. 2016, 590, 1324–1334. [Google Scholar] [CrossRef] [PubMed]
- Friedlander, A.L.; Lynch, D.; Dyar, L.A.; Bowler, R.P. Phenotypes of chronic obstructive pulmonary disease. COPD 2007, 4, 355–384. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.D.; McDonough, J.E.; Zeskind, J.E.; Hackett, T.L.; Pechkovsky, D.V.; Brandsma, C.A.; Suzuki, M.; Gosselink, J.V.; Liu, G.; Alekseyev, Y.O.; et al. A gene expression signature of emphysema-related lung destruction and its reversal by the tripeptide GHK. Genome Med. 2012, 4, 67. [Google Scholar] [CrossRef] [PubMed]
- Van Pottelberge, G.R.; Mestdagh, P.; Bracke, K.R.; Thas, O.; van Durme, Y.M.; Joos, G.F.; Vandesompele, J.; Brusselle, G.G. MicroRNA expression in induced sputum of smokers and patients with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2011, 183, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Oldenburger, A.; van Basten, B.; Kooistra, W.; Meurs, H.; Maarsingh, H.; Krenning, G.; Timens, W.; Schmidt, M. Interaction between Epac1 and miRNA-7 in airway smooth muscle cells. Naunyn-Schmiedeberg Arch. Pharmacol. 2014, 387, 795–797. [Google Scholar] [CrossRef] [PubMed]
- Ezzie, M.E.; Crawford, M.; Cho, J.H.; Orellana, R.; Zhang, S.; Gelinas, R.; Batte, K.; Yu, L.; Nuovo, G.; Galas, D.; et al. Gene expression networks in COPD: MicroRNA and mRNA regulation. Thorax 2012, 67, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Chatila, W.M.; Criner, G.J.; Hancock, W.W.; Akimova, T.; Moldover, B.; Chang, J.K.; Cornwell, W.; Santerre, M.; Rogers, T.J. Blunted expression of miR-199a-5p in regulatory T cells of patients with chronic obstructive pulmonary disease compared to unaffected smokers. Clin. Exp. Immunol. 2014, 177, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Graff, J.W.; Powers, L.S.; Dickson, A.M.; Kim, J.; Reisetter, A.C.; Hassan, I.H.; Kremens, K.; Gross, T.J.; Wilson, M.E.; Monick, M.M. Cigarette smoking decreases global microRNA expression in human alveolar macrophages. PLoS ONE 2012, 7, e44066. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, J.A.; Zago, M.; Nair, P.; Li, P.Z.; Bourbeau, J.; Tan, W.C.; Hamid, Q.; Eidelman, D.H.; Benedetti, A.L.; Baglole, C.J. Decreased expression of the NF-κB family member RelB in lung fibroblasts from smokers with and without COPD potentiates cigarette smoke-induced COX-2 expression. Respir. Res. 2015, 16, 54. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Li, M.; Zhou, S.; Zeng, D.; Xu, X.; Xu, R.; Sun, G. Effect of a single nucleotide polymorphism in miR-146a on COX-2 protein expression and lung function in smokers with chronic obstructive pulmonary disease. Int. J. Chronic Obstr. Pulm. Dis. 2015, 10, 463–473. [Google Scholar]
- Soeda, S.; Ohyashiki, J.H.; Ohtsuki, K.; Umezu, T.; Setoguchi, Y.; Ohyashiki, K. Clinical relevance of plasma miR-106b levels in patients with chronic obstructive pulmonary disease. Int. J. Mol. Med. 2013, 31, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Xin, Q.; Chai, R.; Liu, L.; Ma, Z. Ectopic expressed miR-203 contributes to chronic obstructive pulmonary disease via targeting TAK1 and PIK3CA. Int. J. Clin. Exp. Pathol. 2015, 8, 10662–10670. [Google Scholar] [PubMed]
- World Health Organization. Cardiovascular Disease. Available online: http://www.who.int/cardiovascular_diseases/global-hearts/Global_hearts_initiative/en/ (accessed on 20 November 2016).
- Lachenmeier, D.W.; Przybylski, M.C.; Rehm, J. Comparative risk assessment of carcinogens in alcoholic beverages using the margin of exposure approach. Int. J. Cancer 2012, 131, E995–E1003. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xie, L.; Yang, W.S.; Zhang, W.; Gao, S.; Wang, J.; Xiang, Y.B. Risk factors of hepatocellular carcinoma--current status and perspectives. Asian Pac. J. Cancer Prev. 2012, 13, 743–752. [Google Scholar] [CrossRef] [PubMed]
- McGlynn, K.A.; London, W.T. The global epidemiology of hepatocellular carcinoma: Present and future. Clin. Liver Dis. 2011, 15, 223–243. [Google Scholar] [CrossRef] [PubMed]
- Persson, E.C.; Schwartz, L.M.; Park, Y.; Trabert, B.; Hollenbeck, A.R.; Graubard, B.I.; Freedman, N.D.; McGlynn, K.A. Alcohol consumption, folate intake, hepatocellular carcinoma, and liver disease mortality. Cancer Epidemiol. Biomark. Prev. 2013, 22, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.L.; Qin, S.K. Progress in systemic therapy of advanced hepatocellular carcinoma. World J. Gastroenterol. 2016, 22, 6582–6594. [Google Scholar] [CrossRef] [PubMed]
- Mizuguchi, Y.; Isse, K.; Specht, S.; Lunz, J.G.; Corbitt, N.; Takizawa, T.; Demetris, A.J. Small proline rich protein 2a in benign and malignant liver disease. Hepatology 2014, 59, 1130–1143. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Bukong, T.N.; Kodys, K.; Szabo, G. Alcohol facilitates HCV RNA replication via up-regulation of miR-122 expression and inhibition of cyclin G1 inhuman hepatoma cells. Alcohol. Clin. Exp. Res. 2013, 37, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Bala, S.; Petrasek, J.; Mundkur, S.; Catalano, D.; Levin, I.; Ward, J.; Alao, H.; Kodys, K.; Szabo, G. Circulating microRNAs in exosomes indicate hepatocyte injury and inflammation in alcoholic; drug-induced; and inflammatory liver diseases. Hepatology 2012, 56, 1946–1957. [Google Scholar] [CrossRef] [PubMed]
- Miranda, R.C.; Pietrzykowski, A.Z.; Tang, Y.; Sathyan, P.; Mayfield, D.; Keshavarzian, A.; Sampson, W.; Hereld, D. MicroRNAs: Master regulators of ethanol abuse and toxicity? Alcohol. Clin. Exp. Res. 2010, 34, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Xiao, Q.; Ma, X.P.; Chen, X.; Shi, Z.; Zhang, L.Y.; Chen, H.; Zhang, P.; Ding, D.L.; Huang, H.X.; et al. miR-492G>C polymorphism (rs2289030) is associated with overall survival of hepatocellular carcinoma patients. Tumor Biol. 2016, 37, 8961–8972. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.A.; Kuo, S.Z.; Rahimy, E.; Zou, A.E.; Korrapati, A.; Rahimy, M.; Kim, E.; Zheng, H.; Yu, M.A.; Wang-Rodriguez, J.; et al. Alcohol-dysregulated miR-30a and miR-934 in head and neck squamous cell carcinoma. Mol. Cancer 2015, 14, 181. [Google Scholar] [CrossRef] [PubMed]
- Manikandan, M.; Deva Magendhra Rao, A.K.; Arunkumar, G.; Rajkumar, K.S.; Rajaraman, R.; Munirajan, A.K. Down Regulation of miR-34a and miR-143 May Indirectly Inhibit p53 in Oral Squamous Cell Carcinoma: A Pilot Study. Asian Pac. J. Cancer Prev. 2015, 16, 7619–7625. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Liu, H. MicroRNA-10a-5p and microRNA-34c-5p in laryngeal epithelial premalignant lesions: Differential expression and clinicopathological correlation. Eur. Arch. Otorhinolaryngol. 2015, 272, 391–399. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, R.S.; Dasarathy, S.; McCullough, A.J. Alcoholic liver disease. Am. J. Gastroenterol. 2010, 105, 14–32. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, S.H.; Jin, X.; Li, Y.M. Analysis of differentially expressed genes and microRNAs in alcoholic liver disease. Int. J. Mol. Med. 2013, 31, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.; Momen-Heravi, F.; Kodys, K.; Szabo, G. MicroRNA Cargo of Extracellular Vesicles from Alcohol-exposed Monocytes Signals Naive Monocytes to Differentiate into M2 Macrophages. J. Biol. Chem. 2016, 291, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Momen-Heravi, F.; Saha, B.; Kodys, K.; Catalano, D.; Satishchandran, A.; Szabo, G. Increased number of circulating exosomes and their microRNA cargos are potential novel biomarkers in alcoholic hepatitis. J. Transl. Med. 2015, 13, 261. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Jin, X.; Kong, M.; Li, Y.M. Pattern of microRNA expression associated with different stages of alcoholic liver disease in rat models. Mol. Med. Rep. 2014, 10, 1195–1204. [Google Scholar] [CrossRef] [PubMed]
- Szabo, G.; Mandrekar, P.; Petrasek, J.; Catalano, D. The unfolding web of innate immune dysregulation in alcoholic liver injury. Alcohol. Clin. Exp. Res. 2011, 35, 782–786. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Liang, X.; Jogasuria, A.; Davidson, N.O.; You, M. miR-217 regulates ethanol-induced hepatic inflammation by disrupting sirtuin 1-lipin-1 signaling. Am. J. Pathol. 2015, 185, 1286–1296. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhang, L.; Forsyth, C.B.; Shaikh, M.; Song, S.; Keshavarzian, A. The Role of miR-212 and iNOS in Alcohol-Induced Intestinal Barrier Dysfunction and Steatohepatitis. Alcohol. Clin. Exp. Res. 2015, 39, 1632–1641. [Google Scholar] [CrossRef] [PubMed]
- Cazzoli, R.; Buttitta, F.; Di Nicola, M.; Malatesta, S.; Marchetti, A.; Rom, W.N.; Pass, H.I. MicroRNAs derived from circulating exosomes as noninvasive biomarkers for screening and diagnosing lung cancer. J. Thorac. Oncol. 2013, 8, 1156–1162. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Su, B.; Zhang, P.; Xie, H.; Zheng, H.; Xu, Y.; Du, Q.; Zeng, H.; Zhou, X.; Chen, C.; et al. Expression of miR-150 and miR-3940-5p is reduced in non-small cell lung carcinoma and correlates with clinicopathological features. Oncol. Rep. 2013, 29, 704–712. [Google Scholar] [PubMed]
- Mascaux, C.; Feser, W.J.; Lewis, M.T.; Barón, A.E.; Coldren, C.D.; Merrick, D.T.; Kennedy, T.C.; Eckelberger, J.I.; Rozeboom, L.M.; Franklin, W.A.; et al. Endobronchial miRNAs as biomarkers in lung cancer chemoprevention. Cancer Prev. Res. 2013, 6, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Luo, F.; Liu, Y.; Liu, X.; Shi, L.; Lu, X.; Liu, Q. Posttranscriptional silencing of the lncRNA MALAT1 by miR-217 inhibits the epithelial-mesenchymal transition via enhancer of zeste homolog 2 in the malignant transformation of HBE cells induced by cigarette smoke extract. Toxicol. Appl. Pharmacol. 2015, 289, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Liu, X.; Zhou, Q.; Huang, C.; Meng, X.; Xu, F.; Li, J. Silent information regulator 1 (SIRT1) ameliorates liver fibrosis via promoting activated stellate cell apoptosis and reversion. Toxicol. Appl. Pharmacol. 2015, 289, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Hu, M.; Liang, X.; Ajmo, J.M.; Li, X.; Bataller, R.; Odena, G.; Stevens, S.M.; You, M. Deletion of SIRT1 from hepatocytes in mice disrupts lipin-1 signaling and aggravates alcoholic fatty liver. Gastroenterology 2014, 146, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.J.; Yin, J.Y.; Li, X.P.; Fang, C.; Xiao, D.; Zhang, W.; Zhou, H.H.; Li, X.; Liu, Z.Q. Association of well-characterized lung cancer lncRNA polymorphisms with lung cancer susceptibility and platinum-based chemotherapy response. Tumor Biol. 2016, 37, 8349–8358. [Google Scholar] [CrossRef] [PubMed]
- ENCODE Project Consortium. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar]
- Li, X.; Zhang, Y.; Zhang, H.; Liu, X.; Gong, T.; Li, M.; Sun, L.; Ji, G.; Shi, Y.; Han, Z.; et al. miRNA-223 promotes gastric cancer invasion and metastasis by targeting tumor suppressor EPB41L3. Mol. Cancer Res. 2011, 9, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Du, W.W.; Yang, W.; Rutnam, Z.J.; Peng, C.; Li, H.; O’Malley, Y.Q.; Askeland, R.W.; Sugg, S.; Liu, M.; et al. miR-93 enhances angiogenesis and metastasis by targeting LATS2. Cell Cycle 2012, 11, 4352–4365. [Google Scholar] [CrossRef] [PubMed]
- Malzkorn, B.; Wolter, M.; Liesenberg, F.; Grzendowski, M.; Stühler, K.; Meyer, H.E.; Reifenberger, G. Identification and functional characterization of microRNAs involved in the malignant progression of gliomas. Brain Pathol. 2010, 20, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Nakada, C.; Tsukamoto, Y.; Matsuura, K.; Nguyen, T.L.; Hijiya, N.; Uchida, T.; Sato, F.; Mimata, H.; Seto, M.; Moriyama, M. Overexpression of miR-210, a downstream target of HIF1α, causes centrosome amplification in renal carcinoma cells. J. Pathol. 2011, 224, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Bansal, A.; Lee, I.H.; Hong, X.; Anand, V.; Mathur, S.C.; Gaddam, S.; Rastogi, A.; Wani, S.B.; Gupta, N.; Visvanathan, M.; et al. Feasibility of microRNAs as biomarkers for Barrett’s esophagus progression: A pilot cross-sectional, phase 2 biomarker study. Am. J. Gastroenterol. 2011, 106, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Ding, L.; Bennewith, K.L.; Tong, R.T.; Welford, S.M.; Ang, K.K.; Story, M.; Le, Q.T.; Giaccia, A.J. Hypoxia-inducible miR-210 regulates normoxic gene expression involved in tumor initiation. Mol. Cell 2009, 35, 856–867. [Google Scholar] [CrossRef] [PubMed]
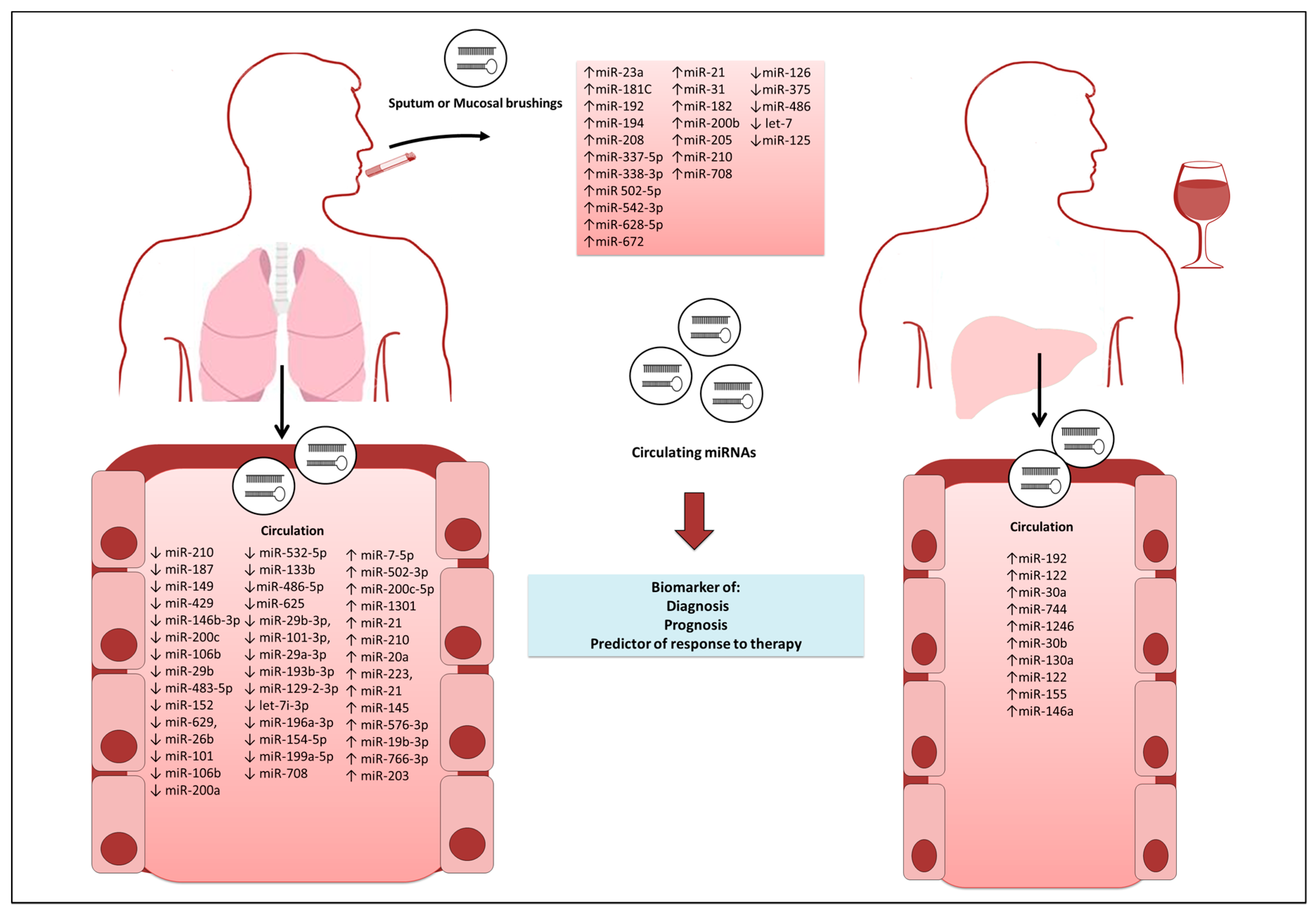

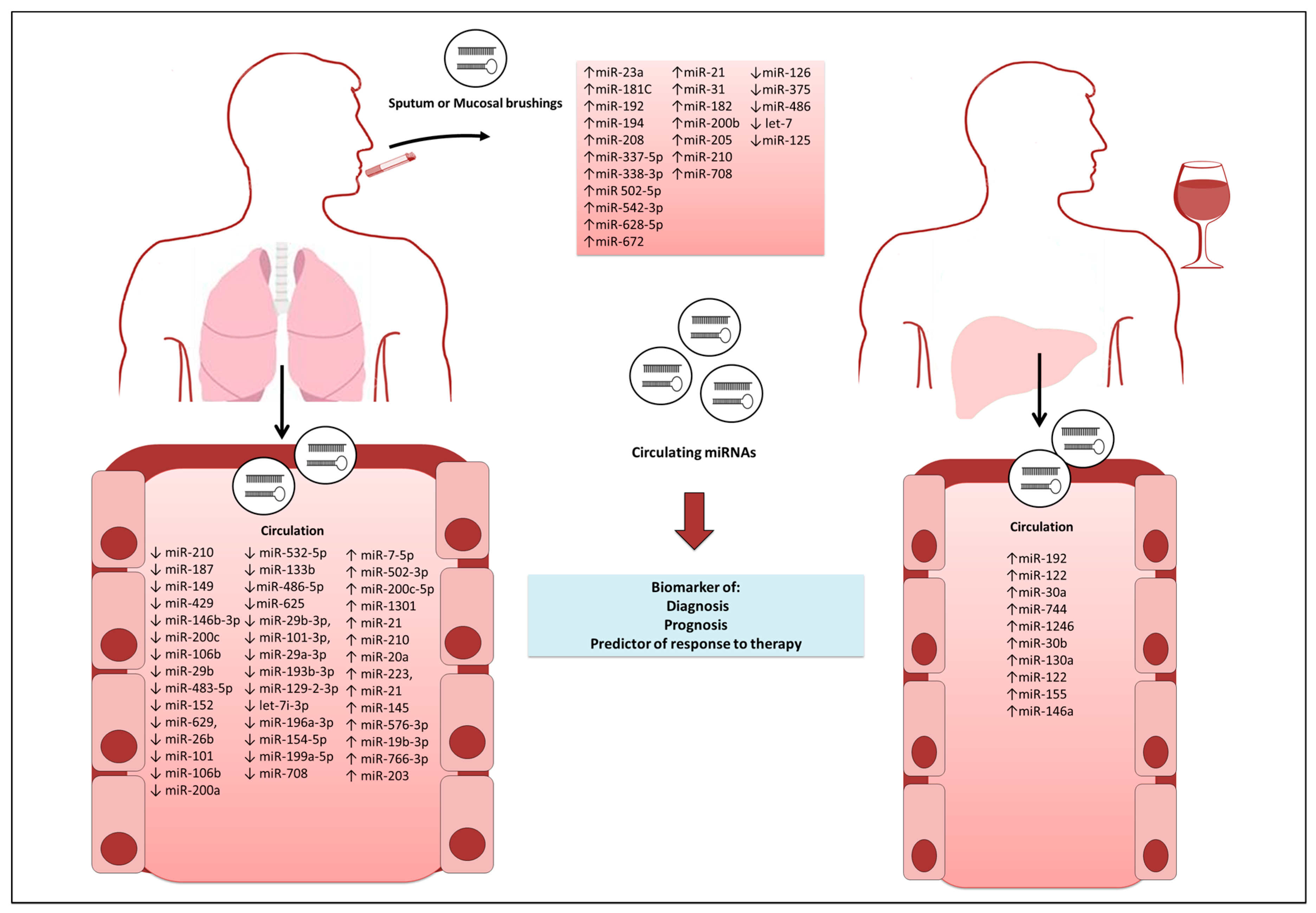{kind=link}
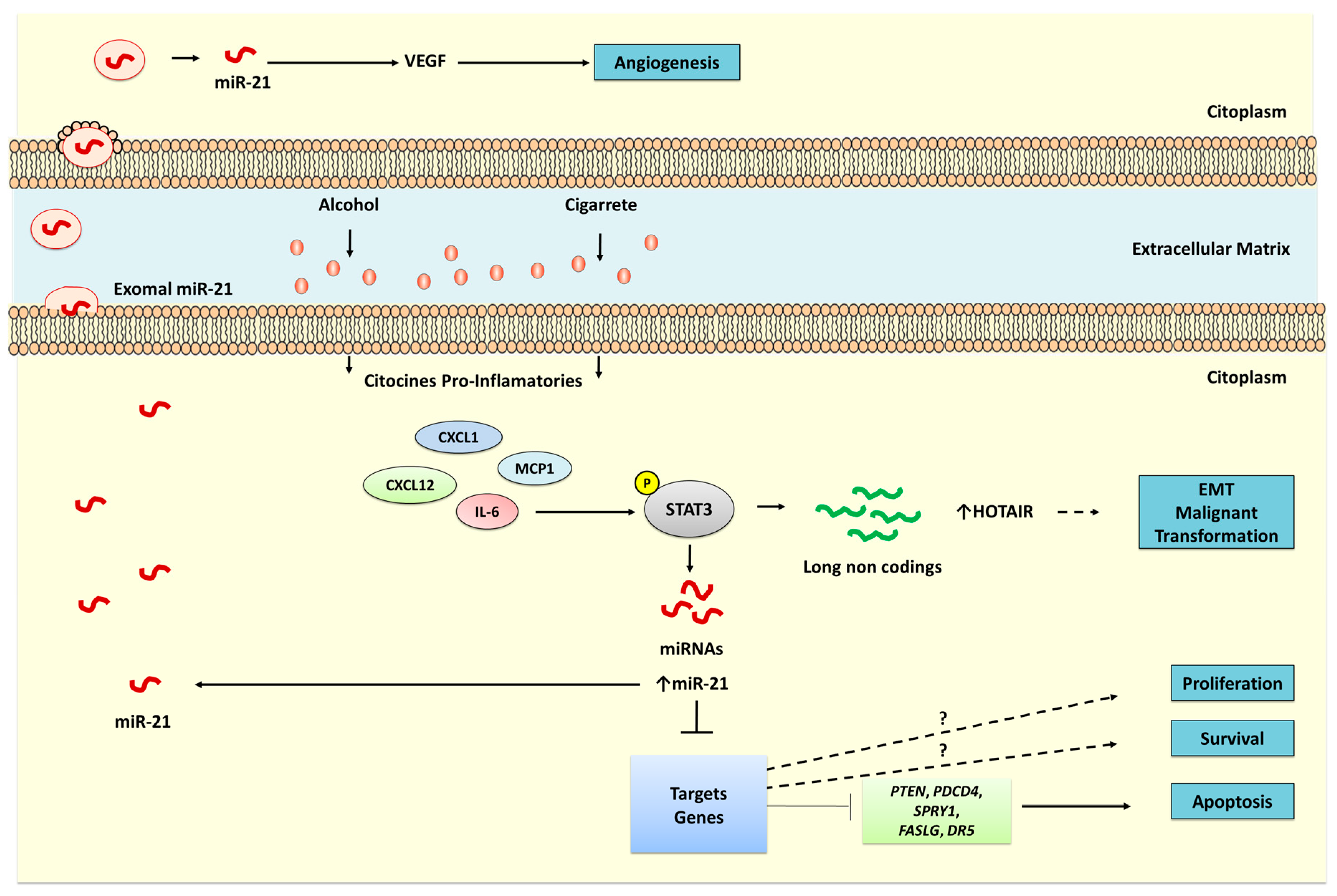
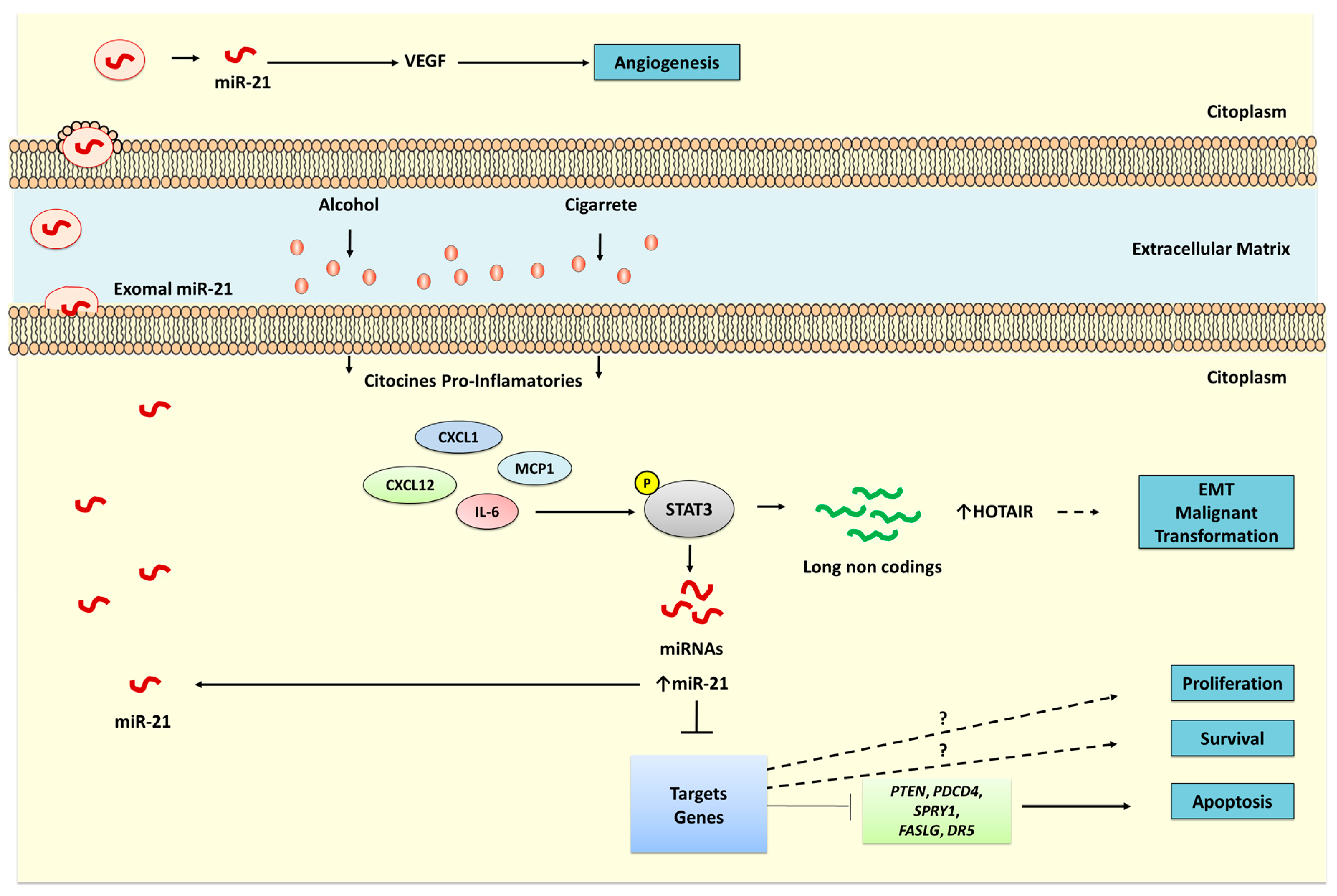{kind=link}
| Cancer | miRNA | Regulation * | Associated Risk Factor | Samples | Ref. # |
|---|---|---|---|---|---|
| Lung | miR-486-5p | ↓ | Tobacco | Plasma | [63] |
| Lung | miR-625 | ↓ | Tobacco | Serum and cell lines | [54] |
| Lung | miR-150 | ↓ | Tobacco | Tumor tissues | [108] |
| Lung | miR-142-3p, miR-34c | ↓ | Tobacco | Tumor tissues | [109] |
| Lung | miR-126, miR-375, miR-486 | ↓ | Tobacco | Sputum | [64] |
| Lung | miR-322, miR-326 | ↓ | Tobacco | Mice | [68] |
| Lung | miR-29b-3p, miR-101-3p, miR-29a-3p, miR-193b-3p | ↓ | Tobacco | Mononuclear cells | [8] |
| Lung | miR-129-2-3p, let-7i-3p, miR-196a-3p, miR-154-5p | ↓ | Tobacco | Serum | [58] |
| Cholangio | miR-200 | ↓ | Alcohol | Tumor tissues | [91] |
| Laryngeal ** | miR-34c-5p | ↓ | Alcohol | Tumor tissues | [98] |
| Esophageal | miR-217 | ↓ | Tobacco | Tumors and cell lines | [60] |
| Gastric | miR-143 | ↓ | Tobacco | Tumor tissues | [70] |
| Lung | miR-7-5p, miR-502-3p, miR-200c-5p, miR-1301 | ↑ | Tobacco | Serum | [58] |
| Lung | miR-23a, miR-181c, miR-192, miR-194, miR-208, miR-337-5p, miR-338-3p, miR-502-5p, miR-542-3p, miR-628-5p, miR-672 | ↑ | Tobacco | Mucosal brushings | [66] |
| Lung | miR-21, miR-210 | ↑ | Tobacco | Plasma | [63] |
| Lung | miR-100 | ↑ | Tobacco | Tumor tissues | [59] |
| Lung | miR-224, miR-375, miR-452 | ↑ | Tobacco | Tumor tissues | [109] |
| Lung | miR-20a, miR-223, miR-21, miR-145 | ↑ | Tobacco | Plasma | [12] |
| Lung | miR-21, miR-31, miR-182, miR-200b, miR-205, miR-210, miR-708 | ↑ | Tobacco | Sputum | [64] |
| Lung | miR-576-3p, miR-19b-3p, miR-766-3p | ↑ | Tobacco | Mononuclear cells | [8] |
| Lung | miR-21 | ↑ | Tobacco | Serum and cell lines | [13] |
| Gastric | miR-21 | ↑ | Tobacco | Brush cytology | [70] |
| Oral | miR-637 | ↑ | Tobacco | Tumor tissues | [71] |
| Liver | miR-34 | ↑ | Alcohol | Mice and cell lines | [30] |
| Liver | miR-21 | ↑ | Alcohol | Mice and cell lines | [9] |
| Oral | miR-34 | ↑ | Alcohol | Tumor tissues | [97] |
| Head and Neck | miR-30a-5p, miR-934, miR-3164, miR-3178 | ↑ | Alcohol | Tumor tissues | [96] |
| Gastric | miR-203, miR-205, miR-223 | ↑ | Alcohol | Tumor tissues | [70] |
| Disease | miRNA | Regulation * | Associated Risk Factor | Samples | Ref. # |
|---|---|---|---|---|---|
| Cardiovascular Diseases | |||||
| Myocardial Fibrosis | miR-133 | ↓ | Tobacco | Cell line | [51] |
| Abdominal aortic aneurysms | miR-21 | ↑ | Tobacco | Tissue, mice, cell line | [52] |
| Intestinal barrier dysfunction | miR-212 | ↑ | Alcohol | Mice and cell line | [106] |
| Respiratory diseases | |||||
| COPD | miR-199a-5p | ↓ | Tobacco | Mononuclear cells | [78] |
| COPD | miR-146b-3p, miR-150, miR-210 | ↓ | Tobacco | Bronchoalveolar lavage | [79] |
| COPD | miR-708, miR-200a, miR-210, miR-187, miR-149, miR-429, miR-146b-3p, miR-200c | ↓ | Tobacco | Alveolar macrophages | [34] |
| COPD | let-7, miR-125 | ↓ | Tobacco | Sputum | [75] |
| COPD | miR-106b, miR-29b, miR-483-5p, miR-152, miR-629, miR 26b, miR-101, miR-106b, miR-532-5p, miR-133b | ↓ | Tobacco | Plasma | [82] |
| COPD | miR-7 | ↑ | Tobacco | Cell line | [76] |
| COPD | miR-203 | ↑ | Tobacco | Blood | [83] |
| COPD | miR-146a | ↑ | Tobacco | Cell line | [80] |
| Liver diseases | |||||
| Alcoholic liver diseases | miR-199-3p, miR-214, miR-93, miR-146a, miR-191, let-7b | ↓ | Alcohol | Tumor tissues | [103] |
| Steatosis | miR-217 | ↑ | Alcohol | Mice and cell line | [105] |
| Alcoholic hepatitis | miR-27a | ↑ | Alcohol | Cell line and plasma | [101] |
| Alcoholic hepatitis | miR-192, miR-122, miR-30A, miR-744, miR-1246, miR-30, miR-130a | ↑ | Alcohol | Human plasma and mice | [102] |
| Hepatitis C Virus | miR-122 | ↑ | Alcohol | Cell line | [92] |
| Alcoholic liver diseases | miR-122, miR-155, miR-146a | ↑ | Alcohol | Plasma and serum of mice | [93] |
| Alcoholic liver diseases | miR-570, miR-122, miR-34b, miR-29c, miR-922, miR-185 | ↑ | Alcohol | Tumor tissues | [100] |
| Alcoholic liver diseases | miR-129, miR-490, miR-21, miR-503, miR-183, miR-185 | ↑ | Alcohol | Tumor tissues | [103] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares do Amaral, N.; Cruz e Melo, N.; De Melo Maia, B.; Malagoli Rocha, R. Noncoding RNA Profiles in Tobacco- and Alcohol-Associated Diseases. Genes 2017, 8, 6. https://doi.org/10.3390/genes8010006
Soares do Amaral N, Cruz e Melo N, De Melo Maia B, Malagoli Rocha R. Noncoding RNA Profiles in Tobacco- and Alcohol-Associated Diseases. Genes. 2017; 8(1):6. https://doi.org/10.3390/genes8010006
Chicago/Turabian StyleSoares do Amaral, Nayra, Natalia Cruz e Melo, Beatriz De Melo Maia, and Rafael Malagoli Rocha. 2017. "Noncoding RNA Profiles in Tobacco- and Alcohol-Associated Diseases" Genes 8, no. 1: 6. https://doi.org/10.3390/genes8010006
APA StyleSoares do Amaral, N., Cruz e Melo, N., De Melo Maia, B., & Malagoli Rocha, R. (2017). Noncoding RNA Profiles in Tobacco- and Alcohol-Associated Diseases. Genes, 8(1), 6. https://doi.org/10.3390/genes8010006