
Genetic Regulation of Virulence and Antibiotic Resistance in Acinetobacter baumannii


Abstract
:1. Introduction
2. Two-Component Systems
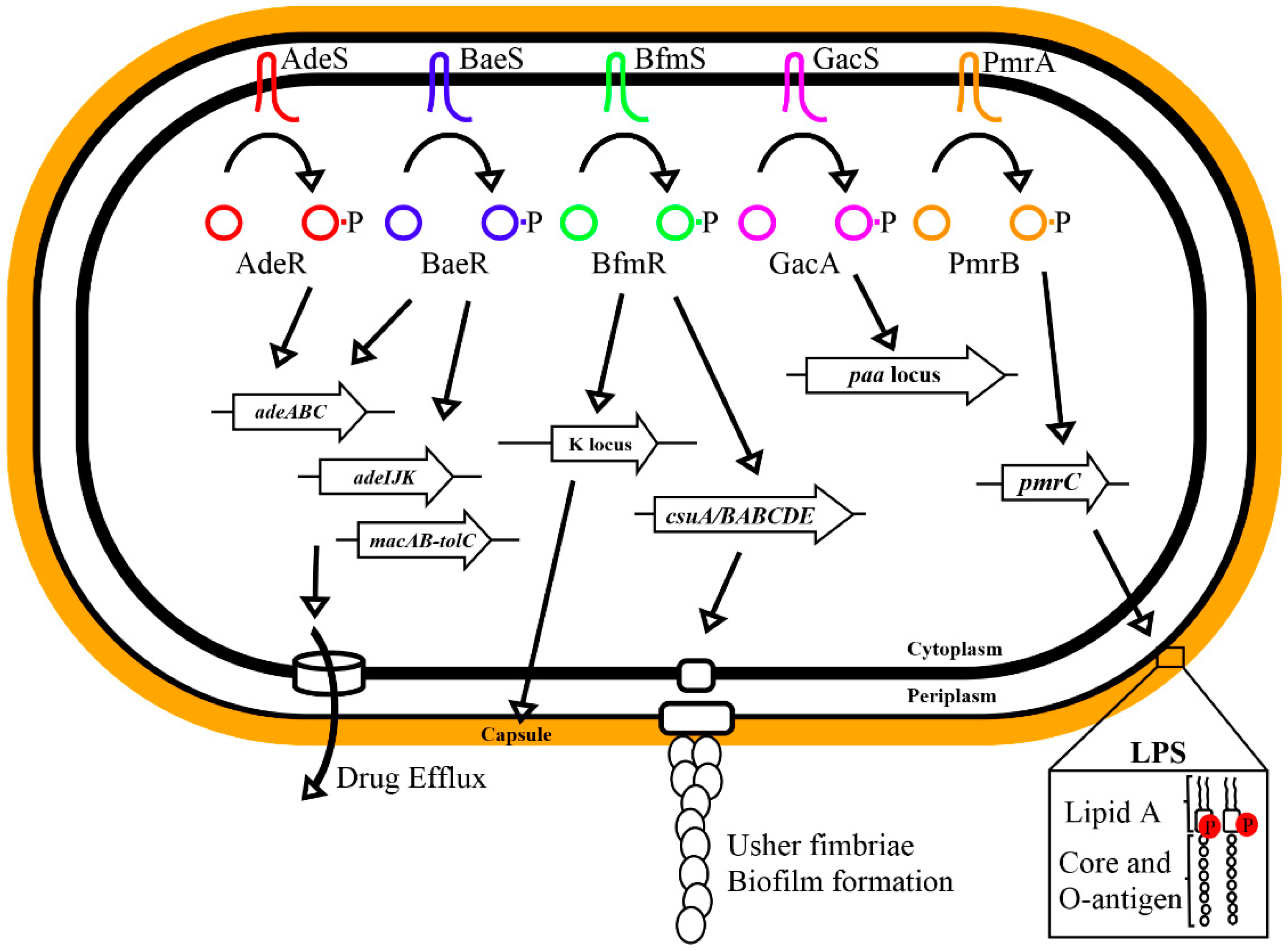
2.1. AdeRS
2.2. BaeSR
2.3. BfmRS
2.4. GacSA
2.5. PmrAB
3. Regulation of Iron and Zinc Acquisition
3.1. Fur—the Ferric Uptake Regulator
3.2. Zur—the Zinc Uptake Regulator
4. Biofilm Formation and Quorum Sensing
5. Other Regulatory Proteins
5.1. AtfA
5.2. Nucleoid-Associated Protein H-NS
6. Global Studies Identifying Important Regulators
7. Do Small Regulatory RNAs Have a Role in A. baumannii Virulence or Antibiotic Resistance?
8. Conclusions and Outlook
Acknowledgments
Conflicts of Interest
References
- Brown, E.D.; Wright, G.D. Antibacterial drug discovery in the resistance era. Nature 2016, 529, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Tommasi, R.; Brown, D.G.; Walkup, G.K.; Manchester, J.I.; Miller, A.A. ESKAPEing the labyrinth of antibacterial discovery. Nat. Rev. Drug Discov. 2015, 14, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad Bugs, No Drugs: No ESKAPE! An Update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.B. Federal Funding for the Study of Antimicrobial Resistance in Nosocomial Pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, B.; Bonomo, R.A. The Deadly Impact of Extreme Drug Resistance in Acinetobacter baumannii. Crit. Care Med. 2014, 42, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Dijkshoorn, L.; Nemec, A.; Seifert, H. An increasing threat in hospitals: Multidrug-resistant Acinetobacter baumannii. Nat. Rev. Microbiol. 2007, 5, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Antunes, L.C.S.; Visca, P.; Towner, K.J. Acinetobacter baumannii: Evolution of a global pathogen. Pathog. Dis. 2014, 71, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Roca, I.; Espinal, P.; Vila-Farrés, X.; Vila, J. The Acinetobacter baumannii Oxymoron: Commensal Hospital Dweller Turned Pan-Drug-Resistant Menace. Front. Microbiol. 2012, 3, 148. [Google Scholar] [CrossRef] [PubMed]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a Successful Pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [PubMed]
- McConnell, M.J.; Actis, L.; Pachón, J. Acinetobacter baumannii: Human infections, factors contributing to pathogenesis and animal models. FEMS Microbiol. Rev. 2012, 37, 130–155. [Google Scholar] [CrossRef] [PubMed]
- Espinal, P.; Roca, I.; Vila, J. Clinical impact and molecular basis of antimicrobial resistance in non-baumannii Acinetobacter. Future Microbiol. 2011, 6, 495–511. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Karveli, E.A.; Kelesidis, I.; Kelesidis, T. Community-acquired Acinetobacter infections. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Dexter, C.; Murray, G.L.; Paulsen, I.T.; Peleg, A.Y. Community-acquired Acinetobacter baumannii: Clinical characteristics, epidemiology and pathogenesis. Expert Rev. Anti Infect. Ther. 2015, 13, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.C.; Thompson, M.G.; Black, C.C.; Kessler, J.L.; Clark, L.P.; McQueary, C.N.; Gancz, H.Y.; Corey, B.W.; Moon, J.K.; Si, Y.; et al. AB5075, a Highly Virulent Isolate of Acinetobacter baumannii, as a Model Strain for the Evaluation of Pathogenesis and Antimicrobial Treatments. mBio 2014, 5, e01076-14. [Google Scholar] [CrossRef] [PubMed]
- Joly-Guillou, M.-L. Clinical impact and pathogenicity of Acinetobacter. Clin. Microbiol. Infect. 2005, 11, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Lemos, E.V.; de la Hoz, F.P.; Einarson, T.R.; McGhan, W.F.; Quevedo, E.; Castañeda, C.; Kawai, K. Carbapenem resistance and mortality in patients with Acinetobacter baumannii infection: Systematic review and meta-analysis. Clin. Microbiol. Infect. 2014, 20, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, B.; Rex, J.H. The value of single-pathogen antibacterial agents. Nat. Rev. Drug Discov. 2013, 12, 963. [Google Scholar] [CrossRef] [PubMed]
- Göttig, S.; Gruber, T.M.; Higgins, P.G.; Wachsmuth, M.; Seifert, H.; Kempf, V.A.J. Detection of pan drug-resistant Acinetobacter baumannii in Germany. J. Antimicrob. Chemother. 2014, 69, 2578–2579. [Google Scholar]
- Chen, Z.; Qiu, S.; Wang, Y.; Wang, Y.; Liu, S.; Wang, Z.; Du, X.; Wang, L.; Guo, J.; Wang, Z.; et al. Coexistence of blaNDM-1 with the Prevalent blaOXA23 and blaIMP in Pan-drug Resistant Acinetobacter baumannii Isolates in China. Clin. Infect. Dis. 2011, 52, 692–693. [Google Scholar] [CrossRef] [PubMed]
- Valencia, R.; Arroyo, L.A.; Conde, M.; Aldana, J.M.; Torres, M.-J.; Fernández-Cuenca, F.; Garnacho-Montero, J.; Cisneros, J.M.; Ortiz, C.; Pachón, J.; et al. Nosocomial Outbreak of Infection With Pan–Drug-Resistant Acinetobacter baumannii in a Tertiary Care University Hospital. Infect. Control Hosp. Epidemiol. 2009, 30, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.H.; Sheng, W.H.; Chang, Y.Y.; Wang, L.H.; Lin, H.C.; Chen, M.L.; Pan, H.J.; Ko, W.J.; Chang, S.C.; Lin, F.Y. Healthcare-associated outbreak due to pan-drug resistant Acinetobacter baumannii in a surgical intensive care unit. J. Hosp. Infect. 2003, 53, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Taherikalani, M.; Etemadi, G.; Geliani, K.N.; Fatollahzadeh, B.; Soroush, S.; Feizabadi, M.M. Emergence of multi and pan-drug resistance Acinetobacter baumannii carrying blaOXA-type -carbapenemase genes among burn patients in Tehran, Iran. Saudi Med. J. 2008, 29, 623–624. [Google Scholar] [PubMed]
- Vila, J.; Pachón, J. Acinetobacter baumannii resistant to everything: What should we do? Clin. Microbiol. Infect. 2011, 17, 955–956. [Google Scholar] [CrossRef] [PubMed]
- Weber, B.S.; Harding, C.M.; Feldman, M.F. Pathogenic Acinetobacter: From the Cell Surface to Infinity and Beyond. J. Bacteriol. 2016, 198, 880–887. [Google Scholar] [CrossRef] [PubMed]
- Groisman, E.A. Feedback Control of Two-Component Regulatory Systems. Annu. Rev. Microbiol. 2016, 70, 103–124. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Groisman, E.A. Acidic pH sensing in the bacterial cytoplasm is required for Salmonella virulence. Mol. Microbiol. 2016, 101, 1024–1038. [Google Scholar] [CrossRef] [PubMed]
- Capra, E.J.; Laub, M.T. Evolution of Two-Component Signal Transduction Systems. Annu. Rev. Microbiol. 2012, 66, 325–347. [Google Scholar] [CrossRef] [PubMed]
- Quinn, H.J.; Cameron, A.D.S.; Dorman, C.J. Bacterial Regulon Evolution: Distinct Responses and Roles for the Identical OmpR Proteins of Salmonella Typhimurium and Escherichia coli in the Acid Stress Response. PLoS Genet. 2014, 10, e1004215. [Google Scholar] [CrossRef] [PubMed]
- Walthers, D.; Carroll, R.K.; Navarre, W.W.; Libby, S.J.; Fang, F.C.; Kenney, L.J. The response regulator SsrB activates expression of diverse Salmonella pathogenicity island 2 promoters and counters silencing by the nucleoid-associated protein H-NS. Mol. Microbiol. 2007, 65, 477–493. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.K.; Winardhi, R.S.; Periasamy, S.; Dykas, M.M.; Jie, Y.; Kenney, L.J. The horizontally-acquired response regulator SsrB drives a Salmonella lifestyle switch by relieving biofilm silencing. eLife 2016, 5, e10747. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, L.A.; Ramage, E.; Weiss, E.J.; Radey, M.; Hayden, H.S.; Held, K.G.; Huse, H.K.; Zurawski, D.V.; Brittnacher, M.J.; Manoil, C. Resources for Genetic and Genomic Analysis of Emerging Pathogen Acinetobacter baumannii. J. Bacteriol. 2015, 197, 2027–2035. [Google Scholar] [CrossRef] [PubMed]
- Ashby, M.K. Survey of the number of two-component response regulator genes in the complete and annotated genome sequences of prokaryotes. FEMS Microbiol. Lett. 2004, 231, 277–281. [Google Scholar] [CrossRef]
- Brittnacher, M.J.; Fong, C.; Hayden, H.S.; Jacobs, M.A.; Radey, M.; Rohmer, L. PGAT: A multistrain analysis resource for microbial genomes. Bioinformatics 2011, 27, 2429–2430. [Google Scholar] [CrossRef] [PubMed]
- Magnet, S.; Courvalin, P.; Lambert, T. Resistance-Nodulation-Cell Division-Type Efflux Pump Involved in Aminoglycoside Resistance in Acinetobacter baumannii Strain BM4454. Antimicrob. Agents Chemother. 2001, 45, 3375–3380. [Google Scholar] [CrossRef] [PubMed]
- Marchand, I.; Damier-Piolle, L.; Courvalin, P.; Lambert, T. Expression of the RND-Type Efflux Pump AdeABC in Acinetobacter baumannii Is Regulated by the AdeRS Two-Component System. Antimicrob. Agents Chemother. 2004, 48, 3298–3304. [Google Scholar] [CrossRef] [PubMed]
- Nemec, A.; Maixnerová, M.; van der Reijden, T.J.K.; van den Broek, P.J.; Dijkshoorn, L. Relationship between the AdeABC efflux system gene content, netilmicin susceptibility and multidrug resistance in a genotypically diverse collection of Acinetobacter baumannii strains. J. Antimicrob. Chemother. 2007, 60, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Ruzin, A.; Keeney, D.; Bradford, P.A. AdeABC multidrug efflux pump is associated with decreased susceptibility to tigecycline in Acinetobacter calcoaceticus–Acinetobacter baumannii complex. J. Antimicrob. Chemother. 2007, 59, 1001–1004. [Google Scholar] [CrossRef] [PubMed]
- Coyne, S.; Guigon, G.; Courvalin, P.; Périchon, B. Screening and Quantification of the Expression of Antibiotic Resistance Genes in Acinetobacter baumannii with a Microarray. Antimicrob. Agents Chemother. 2010, 54, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.-J.; Courvalin, P.; Grillot-Courvalin, C. RND-Type Efflux Pumps in Multidrug-Resistant Clinical Isolates of Acinetobacter baumannii: Major Role for AdeABC Overexpression and AdeRS Mutations. Antimicrob. Agents Chemother. 2013, 57, 2989–2995. [Google Scholar] [CrossRef] [PubMed]
- Hornsey, M.; Ellington, M.J.; Doumith, M.; Thomas, C.P.; Gordon, N.C.; Wareham, D.W.; Quinn, J.; Lolans, K.; Livermore, D.M.; Woodford, N. AdeABC-mediated efflux and tigecycline MICs for epidemic clones of Acinetobacter baumannii. J. Antimicrob. Chemother. 2010, 65, 1589–1593. [Google Scholar] [CrossRef] [PubMed]
- Nowak, J.; Schneiders, T.; Seifert, H.; Higgins, P.G. The Asp20-to-Asn Substitution in the Response Regulator AdeR Leads to Enhanced Efflux Activity of AdeB in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2016, 60, 1085–1090. [Google Scholar] [CrossRef] [PubMed]
- Montaña, S.; Vilacoba, E.; Traglia, G.M.; Almuzara, M.; Pennini, M.; Fernández, A.; Sucari, A.; Centrón, D.; Ramírez, M.S. Genetic Variability of AdeRS Two-Component System Associated with Tigecycline Resistance in XDR-Acinetobacter baumannii Isolates. Curr. Microbiol. 2015, 71, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.-R.; Chan, M.-C.; Chang, T.-Y.; Wang, W.-Y.; Chiueh, T.-S. Overexpression of the adeB Gene in Clinical Isolates of Tigecycline-Nonsusceptible Acinetobacter baumannii without Insertion Mutations in adeRS. Antimicrob. Agents Chemother. 2010, 54, 4934–4938. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.-J.; Balloy, V.; Fiette, L.; Chignard, M.; Courvalin, P.; Grillot-Courvalin, C. Contribution of the Ade Resistance-Nodulation-Cell Division-Type Efflux Pumps to Fitness and Pathogenesis of Acinetobacter baumannii. mBio 2016, 7, e00697-16. [Google Scholar] [CrossRef] [PubMed]
- Higgins, P.G.; Schneiders, T.; Hamprecht, A.; Seifert, H. In Vivo Selection of a Missense Mutation in adeR and Conversion of the Novel blaOXA-164 Gene into blaOXA-58 in Carbapenem-Resistant Acinetobacter baumannii Isolates from a Hospitalized Patient. Antimicrob. Agents Chemother. 2010, 54, 5021–5027. [Google Scholar] [CrossRef] [PubMed]
- Coyne, S.; Courvalin, P.; Périchon, B. Efflux-Mediated Antibiotic Resistance in Acinetobacter spp. Antimicrob. Agents Chemother. 2011, 55, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Richmond, G.E.; Evans, L.P.; Anderson, M.J.; Wand, M.E.; Bonney, L.C.; Ivens, A.; Chua, K.L.; Webber, M.A.; Sutton, J.M.; Peterson, M.L.; et al. The Acinetobacter baumannii Two-Component System AdeRS Regulates Genes Required for Multidrug Efflux, Biofilm Formation, and Virulence in a Strain-Specific Manner. mBio 2016, 7, e00430-16. [Google Scholar] [PubMed]
- Wright, M.S.; Iovleva, A.; Jacobs, M.R.; Bonomo, R.A.; Adams, M.D. Genome dynamics of multidrug-resistant Acinetobacter baumannii during infection and treatment. Genome Med. 2016, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.S.; Haft, D.H.; Harkins, D.M.; Perez, F.; Hujer, K.M.; Bajaksouzian, S.; Benard, M.F.; Jacobs, M.R.; Bonomo, R.A.; Adams, M.D. New Insights into Dissemination and Variation of the Health Care-Associated Pathogen Acinetobacter baumannii from Genomic Analysis. mBio 2014, 5, e00963-13. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-F.; Lin, Y.-Y.; Yeh, H.-W.; Lan, C.-Y. Role of the BaeSR two-component system in the regulation of Acinetobacter baumannii adeAB genes and its correlation with tigecycline susceptibility. BMC Microbiol. 2014, 14, 119. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-F.; Lin, Y.-Y.; Lan, C.-Y. The Role of the Two-Component System BaeSR in Disposing Chemicals through Regulating Transporter Systems in Acinetobacter baumannii. PLoS ONE 2015, 10, e0132843. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H.; Fujii, K.; Yamasaki, O.; Oono, T.; Iwatsuki, K. Antibacterial action of several tannins against Staphylococcus aureus. J. Antimicrob. Chemother. 2001, 48, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Chusri, S.; Villanueva, I.; Voravuthikunchai, S.P.; Davies, J. Enhancing antibiotic activity: A strategy to control Acinetobacter infections. J. Antimicrob. Chemother. 2009, 64, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Tomaras, A.P.; Flagler, M.J.; Dorsey, C.W.; Gaddy, J.A.; Actis, L.A. Characterization of a two-component regulatory system from Acinetobacter baumannii that controls biofilm formation and cellular morphology. Microbiology 2008, 154, 3398–3409. [Google Scholar] [CrossRef] [PubMed]
- Gaddy, J.A.; Actis, L.A. Regulation of Acinetobacter baumannii biofilm formation. Future Microbiol. 2009, 4, 273–278. [Google Scholar] [CrossRef] [PubMed]
- de Breij, A.; Gaddy, J.; van der Meer, J.; Koning, R.; Koster, A.; van den Broek, P.; Actis, L.; Nibbering, P.; Dijkshoorn, L. CsuA/BABCDE-dependent pili are not involved in the adherence of Acinetobacter baumannii ATCC19606T to human airway epithelial cells and their inflammatory response. Res. Microbiol. 2009, 160, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Liou, M.-L.; Soo, P.-C.; Ling, S.-R.; Kuo, H.-Y.; Tang, C.Y.; Chang, K.-C. The sensor kinase BfmS mediates virulence in Acinetobacter baumannii. J. Microbiol. Immunol. Infect. 2014, 47, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Russo, T.A.; Manohar, A.; Beanan, J.M.; Olson, R.; MacDonald, U.; Graham, J.; Umland, T.C. The Response Regulator BfmR Is a Potential Drug Target for Acinetobacter baumannii. mSphere 2016, 1, e00082-16. [Google Scholar] [CrossRef] [PubMed]
- Geisinger, E.; Isberg, R.R. Antibiotic Modulation of Capsular Exopolysaccharide and Virulence in Acinetobacter baumannii. PLoS Pathog. 2015, 11, e1004691. [Google Scholar] [CrossRef] [PubMed]
- Dorsey, C.W.; Tomaras, A.P.; Actis, L.A. Genetic and Phenotypic Analysis of Acinetobacter baumannii Insertion Derivatives Generated with a Transposome System. Appl. Environ. Microbiol. 2002, 68, 6353–6360. [Google Scholar] [CrossRef] [PubMed]
- Peleg, A.Y.; Tampakakis, E.; Fuchs, B.B.; Eliopoulos, G.M.; Moellering, R.C.; Mylonakis, E. Prokaryote–eukaryote interactions identified by using Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2008, 105, 14585–14590. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira, G.M.; Kostoulias, X.; Khoo, C.; Aibinu, I.; Qu, Y.; Traven, A.; Peleg, A.Y. A Global Virulence Regulator in Acinetobacter baumannii and Its Control of the Phenylacetic Acid Catabolic Pathway. J. Infect. Dis. 2014, 210, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, M.S.; Ellett, F.; Murray, G.L.; Kostoulias, X.; Cerqueira, G.M.; Schulze, K.E.; Maifiah, M.H.M.; Li, J.; Creek, D.J.; Lieschke, G.J.; et al. Acinetobacter baumannii phenylacetic acid metabolism influences infection outcome through a direct effect on neutrophil chemotaxis. Proc. Natl. Acad. Sci. USA 2016, 113, 9599–9604. [Google Scholar] [CrossRef] [PubMed]
- Teufel, R.; Mascaraque, V.; Ismail, W.; Voss, M.; Perera, J.; Eisenreich, W.; Haehnel, W.; Fuchs, G. Bacterial phenylalanine and phenylacetate catabolic pathway revealed. Proc. Natl. Acad. Sci. USA 2010, 107, 14390–14395. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, L.A.; Herrera, C.M.; Fernandez, L.; Hankins, J.V.; Trent, M.S.; Hancock, R.E.W. The pmrCAB Operon Mediates Polymyxin Resistance in Acinetobacter baumannii ATCC 17978 and Clinical Isolates through Phosphoethanolamine Modification of Lipid A. Antimicrob. Agents Chemother. 2011, 55, 3743–3751. [Google Scholar] [CrossRef] [PubMed]
- Eijkelkamp, B.A.; Hassan, K.A.; Paulsen, I.T.; Brown, M.H. Investigation of the human pathogen Acinetobacter baumannii under iron limiting conditions. BMC Genomics 2011, 12, 126. [Google Scholar] [CrossRef] [PubMed]
- Hood, M.I.; Mortensen, B.L.; Moore, J.L.; Zhang, Y.; Kehl-Fie, T.E.; Sugitani, N.; Chazin, W.J.; Caprioli, R.M.; Skaar, E.P. Identification of an Acinetobacter baumannii Zinc Acquisition System that Facilitates Resistance to Calprotectin-mediated Zinc Sequestration. PLoS Pathog. 2012, 8, e1003068. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, B.L.; Rathi, S.; Chazin, W.J.; Skaar, E.P. Acinetobacter baumannii Response to Host-Mediated Zinc Limitation Requires the Transcriptional Regulator Zur. J. Bacteriol. 2014, 196, 2616–2626. [Google Scholar] [CrossRef] [PubMed]
- Anbazhagan, D.; Mansor, M.; Yan, G.O.S.; Yusof, M.Y.M.; Hassan, H.; Sekaran, S.D. Detection of Quorum Sensing Signal Molecules and Identification of an Autoinducer Synthase Gene among Biofilm Forming Clinical Isolates of Acinetobacter spp. PLoS ONE 2012, 7, e36696. [Google Scholar] [CrossRef] [PubMed]
- Modarresi, F.; Azizi, O.; Shakibaie, M.R.; Motamedifar, M.; Mosadegh, E.; Mansouri, S. Iron limitation enhances acyl homoserine lactone (AHL) production and biofilm formation in clinical isolates of Acinetobacter baumannii. Virulence 2015, 6, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Withers, R.; Doherty, G.P.; Jordan, M.; Yang, X.; Dixon, N.E.; Lewis, P.J. AtfA, a new factor in global regulation of transcription in Acinetobacter spp. Mol. Microbiol. 2014, 93, 1130–1143. [Google Scholar] [PubMed]
- Tendeng, C.; Krin, E.; Soutourina, O.A.; Marin, A.; Danchin, A.; Bertin, P.N. A Novel H-NS-like Protein from an Antarctic Psychrophilic Bacterium Reveals a Crucial Role for the N-terminal Domain in Thermal Stability. J. Biol. Chem. 2003, 278, 18754–18760. [Google Scholar] [CrossRef] [PubMed]
- Eijkelkamp, B.A.; Stroeher, U.H.; Hassan, K.A.; Elbourne, L.D.H.; Paulsen, I.T.; Brown, M.H. H-NS Plays a Role in Expression of Acinetobacter baumannii Virulence Features. Infect. Immun. 2013, 81, 2574–2583. [Google Scholar] [CrossRef] [PubMed]
- Kaye, K.S.; Pogue, J.M.; Tran, T.B.; Nation, R.L.; Li, J. Agents of Last Resort: Polymyxin Resistance. Infect. Dis. Clin. N. Am. 2016, 30, 391–414. [Google Scholar] [CrossRef] [PubMed]
- Trimble, M.J.; Mlynárčik, P.; Kolář, M.; Hancock, R.E.W. Polymyxin: Alternative Mechanisms of Action and Resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025288. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.D.; Nickel, G.C.; Bajaksouzian, S.; Lavender, H.; Murthy, A.R.; Jacobs, M.R.; Bonomo, R.A. Resistance to Colistin in Acinetobacter baumannii Associated with Mutations in the PmrAB Two-Component System. Antimicrob. Agents Chemother. 2009, 53, 3628–3634. [Google Scholar] [CrossRef] [PubMed]
- Rolain, J.-M.; Diene, S.M.; Kempf, M.; Gimenez, G.; Robert, C.; Raoult, D. Real-Time Sequencing To Decipher the Molecular Mechanism of Resistance of a Clinical Pan-Drug-Resistant Acinetobacter baumannii Isolate from Marseille, France. Antimicrob. Agents Chemother. 2013, 57, 592–596. [Google Scholar] [CrossRef] [PubMed]
- McPhee, J.B.; Lewenza, S.; Hancock, R.E.W. Cationic antimicrobial peptides activate a two-component regulatory system, PmrA-PmrB, that regulates resistance to polymyxin B and cationic antimicrobial peptides in Pseudomonas aeruginosa. Mol. Microbiol. 2003, 50, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Moskowitz, S.M.; Ernst, R.K.; Miller, S.I. PmrAB, a Two-Component Regulatory System of Pseudomonas aeruginosa That Modulates Resistance to Cationic Antimicrobial Peptides and Addition of Aminoarabinose to Lipid A. J. Bacteriol. 2004, 186, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.-Y.; Chen, Y.-F.; Peng, H.-L. Molecular characterization of the PhoPQ-PmrD-PmrAB mediated pathway regulating polymyxin B resistance in Klebsiella pneumoniae CG43. J. Biomed. Sci. 2010, 17, 60. [Google Scholar] [CrossRef] [PubMed]
- Matamouros, S.; Miller, S.I.S. Typhimurium strategies to resist killing by cationic antimicrobial peptides. Biochim. Biophys. Acta BBA - Biomembr. 2015, 1848, 3021–3025. [Google Scholar] [CrossRef] [PubMed]
- Marchal, K.; De Keersmaecker, S.; Monsieurs, P.; van Boxel, N.; Lemmens, K.; Thijs, G.; Vanderleyden, J.; De Moor, B. In silico identification and experimental validation of PmrAB targets in Salmonella typhimurium by regulatory motif detection. Genome Biol. 2004, 5, R9. [Google Scholar] [CrossRef] [PubMed]
- Gunn, J.S.; Lim, K.B.; Krueger, J.; Kim, K.; Guo, L.; Hackett, M.; Miller, S.I. PmrA–PmrB-regulated genes necessary for 4-aminoarabinose lipid A modification and polymyxin resistance. Mol. Microbiol. 1998, 27, 1171–1182. [Google Scholar] [CrossRef] [PubMed]
- Gunn, J.S.; Ryan, S.S.; Velkinburgh, J.C.V.; Ernst, R.K.; Miller, S.I. Genetic and Functional Analysis of a PmrA-PmrB-Regulated Locus Necessary for Lipopolysaccharide Modification, Antimicrobial Peptide Resistance, and Oral Virulence of Salmonella enterica Serovar Typhimurium. Infect. Immun. 2000, 68, 6139–6146. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Hsu, F.-F.; Turk, J.; Groisman, E.A. The PmrA-Regulated pmrC Gene Mediates Phosphoethanolamine Modification of Lipid A and Polymyxin Resistance in Salmonella enterica. J. Bacteriol. 2004, 186, 4124–4133. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, A.O.; Morand, S.; Rolain, J.-M. Mechanisms of polymyxin resistance: Acquired and intrinsic resistance in bacteria. Front. Microbiol. 2014, 5, 643. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.D.; Groisman, E.A. The Biology of the PmrA/PmrB Two-Component System: The Major Regulator of Lipopolysaccharide Modifications. Annu. Rev. Microbiol. 2013, 67, 83–112. [Google Scholar] [CrossRef] [PubMed]
- Beceiro, A.; Llobet, E.; Aranda, J.; Bengoechea, J.A.; Doumith, M.; Hornsey, M.; Dhanji, H.; Chart, H.; Bou, G.; Livermore, D.M.; et al. Phosphoethanolamine Modification of Lipid A in Colistin-Resistant Variants of Acinetobacter baumannii Mediated by the pmrAB Two-Component Regulatory System. Antimicrob. Agents Chemother. 2011, 55, 3370–3379. [Google Scholar] [CrossRef] [PubMed]
- Moffatt, J.H.; Harper, M.; Harrison, P.; Hale, J.D.F.; Vinogradov, E.; Seemann, T.; Henry, R.; Crane, B.; Michael, F.S.; Cox, A.D.; et al. Colistin Resistance in Acinetobacter baumannii Is Mediated by Complete Loss of Lipopolysaccharide Production. Antimicrob. Agents Chemother. 2010, 54, 4971–4977. [Google Scholar] [CrossRef] [PubMed]
- López-Rojas, R.; García-Quintanilla, M.; Labrador-Herrera, G.; Pachón, J.; McConnell, M.J. Impaired growth under iron-limiting conditions associated with the acquisition of colistin resistance in Acinetobacter baumannii. Int. J. Antimicrob. Agents 2016, 47, 473–477. [Google Scholar] [CrossRef] [PubMed]
- López-Rojas, R.; Domínguez-Herrera, J.; McConnell, M.J.; Docobo-Peréz, F.; Smani, Y.; Fernández-Reyes, M.; Rivas, L.; Pachón, J. Impaired Virulence and In Vivo Fitness of Colistin-Resistant Acinetobacter baumannii. J. Infect. Dis. 2011, 203, 545–548. [Google Scholar] [CrossRef] [PubMed]
- López-Rojas, R.; McConnell, M.J.; Jiménez-Mejías, M.E.; Domínguez-Herrera, J.; Fernández-Cuenca, F.; Pachón, J. Colistin Resistance in a Clinical Acinetobacter baumannii Strain Appearing after Colistin Treatment: Effect on Virulence and Bacterial Fitness. Antimicrob. Agents Chemother. 2013, 57, 4587–4589. [Google Scholar] [CrossRef] [PubMed]
- Rolain, J.-M.; Roch, A.; Castanier, M.; Papazian, L.; Raoult, D. Acinetobacter baumannii Resistant to Colistin With Impaired Virulence: A Case Report From France. J. Infect. Dis. 2011, 204, 1146–1147. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Reyes, M.; Rodríguez-Falcón, M.; Chiva, C.; Pachón, J.; Andreu, D.; Rivas, L. The cost of resistance to colistin in Acinetobacter baumannii: A proteomic perspective. Proteomics 2009, 9, 1632–1645. [Google Scholar] [CrossRef] [PubMed]
- Hraiech, S.; Roch, A.; Lepidi, H.; Atieh, T.; Audoly, G.; Rolain, J.-M.; Raoult, D.; Brunel, J.-M.; Papazian, L.; Brégeon, F. Impaired Virulence and Fitness of a Colistin-Resistant Clinical Isolate of Acinetobacter baumannii in a Rat Model of Pneumonia. Antimicrob. Agents Chemother. 2013, 57, 5120–5121. [Google Scholar] [CrossRef] [PubMed]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen–host interface. Nat. Rev. Microbiol. 2012, 10, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Ochoa, V.E.; Jellbauer, S.; Klaus, S.; Raffatellu, M. Transition metal ions at the crossroads of mucosal immunity and microbial pathogenesis. Front. Cell. Infect. Microbiol. 2014, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Palmer, L.D.; Skaar, E.P. Transition Metals and Virulence in Bacteria. Annu. Rev. Genet. 2016, 50, 67–91. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Okujo, N.; Sakakibara, Y. Isolation and structure elucidation of acinetobactin, a novel siderophore from Acinetobacter baumannii. Arch. Microbiol. 1994, 162, 249–254. [Google Scholar] [PubMed]
- Antunes, L.C.S.; Imperi, F.; Towner, K.J.; Visca, P. Genome-assisted identification of putative iron-utilization genes in Acinetobacter baumannii and their distribution among a genotypically diverse collection of clinical isolates. Res. Microbiol. 2011, 162, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Mihara, K.; Tanabe, T.; Yamakawa, Y.; Funahashi, T.; Nakao, H.; Narimatsu, S.; Yamamoto, S. Identification and transcriptional organization of a gene cluster involved in biosynthesis and transport of acinetobactin, a siderophore produced by Acinetobacter baumannii ATCC 19606T. Microbiology 2004, 150, 2587–2597. [Google Scholar] [CrossRef] [PubMed]
- Dorsey, C.W.; Tomaras, A.P.; Connerly, P.L.; Tolmasky, M.E.; Crosa, J.H.; Actis, L.A. The siderophore-mediated iron acquisition systems of Acinetobacter baumannii ATCC 19606 and Vibrio anguillarum 775 are structurally and functionally related. Microbiology 2004, 150, 3657–3667. [Google Scholar] [CrossRef] [PubMed]
- Gaddy, J.A.; Arivett, B.A.; McConnell, M.J.; López-Rojas, R.; Pachón, J.; Actis, L.A. Role of Acinetobactin-Mediated Iron Acquisition Functions in the Interaction of Acinetobacter baumannii Strain ATCC 19606T with Human Lung Epithelial Cells, Galleria mellonella Caterpillars, and Mice. Infect. Immun. 2012, 80, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Daniel, C.; Haentjens, S.; Bissinger, M.-C.; Courcol, R.J. Characterization of the Acinetobacter baumannii Fur regulator: Cloning and sequencing of the fur homolog gene. FEMS Microbiol. Lett. 1999, 170, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Zimbler, D.L.; Arivett, B.A.; Beckett, A.C.; Menke, S.M.; Actis, L.A. Functional Features of TonB Energy Transduction Systems of Acinetobacter baumannii. Infect. Immun. 2013, 81, 3382–3394. [Google Scholar] [CrossRef] [PubMed]
- Postle, K.; Kadner, R.J. Touch and go: Tying TonB to transport. Mol. Microbiol. 2003, 49, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Häse, C.C.; Finkelstein, R.A. Bacterial extracellular zinc-containing metalloproteases. Microbiol. Rev. 1993, 57, 823–837. [Google Scholar] [PubMed]
- Antunes, L.C.S.; Imperi, F.; Minandri, F.; Visca, P. In Vitro and In Vivo Antimicrobial Activities of Gallium Nitrate against Multidrug-Resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2012, 56, 5961–5970. [Google Scholar] [CrossRef] [PubMed]
- Runci, F.; Bonchi, C.; Frangipani, E.; Visaggio, D.; Visca, P. Acinetobacter baumannii biofilm formation in Human serum and disruption by gallium. Antimicrob. Agents Chemother. 2016, 61, e01563-16. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, M.Y.; Switzer, B.L.; Goss, C.H.; Aitken, M.L.; Singh, P.K.; Britigan, B.E. Gallium Compounds Exhibit Potential as New Therapeutic Agents against Mycobacterium abscessus. Antimicrob. Agents Chemother. 2015, 59, 4826. [Google Scholar] [CrossRef] [PubMed]
- Chitambar, C.R. Gallium and its competing roles with iron in biological systems. Biochim. Biophys. Acta 2016, 1863, 2044–2053. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.; Clemmer, K.M.; Bonomo, R.A.; Rather, P.N. Isolation and Characterization of an Autoinducer Synthase from Acinetobacter baumannii. J. Bacteriol. 2008, 190, 3386–3392. [Google Scholar] [CrossRef] [PubMed]
- Case, R.J.; Labbate, M.; Kjelleberg, S. AHL-driven quorum-sensing circuits: Their frequency and function among the Proteobacteria. ISME J. 2008, 2, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Antunes, L.C.M.; Ferreira, R.B.R.; Buckner, M.M.C.; Finlay, B.B. Quorum sensing in bacterial virulence. Microbiology 2010, 156, 2271–2282. [Google Scholar] [CrossRef] [PubMed]
- Patankar, A.V.; González, J.E. Orphan LuxR regulators of quorum sensing. FEMS Microbiol. Rev. 2009, 33, 739–756. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, A.L.; Lappala, C.R.; Morlen, R.P.; Pelletier, D.A.; Lu, T.-Y.S.; Lankford, P.K.; Harwood, C.S.; Greenberg, E.P. LuxR- and LuxI-Type Quorum-Sensing Circuits Are Prevalent in Members of the Populus deltoides Microbiome. Appl. Environ. Microbiol. 2013, 79, 5745–5752. [Google Scholar] [CrossRef] [PubMed]
- Stöveken, T.; Kalscheuer, R.; Malkus, U.; Reichelt, R.; Steinbüchel, A. The Wax Ester Synthase/Acyl Coenzyme A:Diacylglycerol Acyltransferase from Acinetobacter sp. Strain ADP1: Characterization of a Novel Type of Acyltransferase. J. Bacteriol. 2005, 187, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Röttig, A.; Steinbüchel, A. Random mutagenesis of atfA and screening for Acinetobacter baylyi mutants with an altered lipid accumulation. Eur. J. Lipid Sci. Technol. 2013, 115, 394–404. [Google Scholar] [CrossRef]
- Camarena, L.; Bruno, V.; Euskirchen, G.; Poggio, S.; Snyder, M. Molecular Mechanisms of Ethanol-Induced Pathogenesis Revealed by RNA-Sequencing. PLoS Pathog. 2010, 6, e1000834. [Google Scholar] [CrossRef] [PubMed]
- Badrinarayanan, A.; Le, T.B.K.; Laub, M.T. Bacterial Chromosome Organization and Segregation. Annu. Rev. Cell Dev. Biol. 2015, 31, 171–199. [Google Scholar] [CrossRef] [PubMed]
- Dorman, C.J.; Colgan, A.; Dorman, M.J. Bacterial pathogen gene regulation: A DNA-structure-centred view of a protein-dominated domain. Clin. Sci. 2016, 130, 1165–1177. [Google Scholar] [CrossRef] [PubMed]
- Dillon, S.C.; Dorman, C.J. Bacterial nucleoid-associated proteins, nucleoid structure and gene expression. Nat. Rev. Microbiol. 2010, 8, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Barquist, L.; Boinett, C.J.; Cain, A.K. Approaches to querying bacterial genomes with transposon-insertion sequencing. RNA Biol. 2013, 10, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Gray, A.N.; Koo, B.-M.; Shiver, A.L.; Peters, J.M.; Osadnik, H.; Gross, C.A. High-throughput bacterial functional genomics in the sequencing era. Curr. Opin. Microbiol. 2015, 27, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, R.R.; Morgan, E.; Peters, S.E.; Pleasance, S.J.; Hudson, D.L.; Davies, H.M.; Wang, J.; van Diemen, P.M.; Buckley, A.M.; Bowen, A.J.; et al. Comprehensive Assignment of Roles for Salmonella Typhimurium Genes in Intestinal Colonization of Food-Producing Animals. PLoS Genet. 2013, 9, e1003456. [Google Scholar] [CrossRef] [PubMed]
- van Opijnen, T.; Camilli, A. A fine scale phenotype–genotype virulence map of a bacterial pathogen. Genome Res. 2012, 22, 2541–2551. [Google Scholar] [CrossRef] [PubMed]
- Perry, B.J.; Yost, C.K. Construction of a mariner-based transposon vector for use in insertion sequence mutagenesis in selected members of the Rhizobiaceae. BMC Microbiol. 2014, 14, 298. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Ozer, E.A.; Mandel, M.J.; Hauser, A.R. Genome-Wide Identification of Acinetobacter baumannii Genes Necessary for Persistence in the Lung. mBio 2014, 5, e01163-14. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, B.; Kur, J. In vitro interaction of the IHF-like proteins Acinetobacter junii and Proteus vulgaris with ihf sites. FEMS Microbiol. Lett. 1998, 161, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, M.J.; Gallagher, L.A.; Jacobson, R.K.; Usacheva, E.A.; Peterson, L.R.; Zurawski, D.V.; Shuman, H.A. Joint Transcriptional Control of Virulence and Resistance to Antibiotic and Environmental Stress in Acinetobacter baumannii. mBio 2015, 6, e01660-15. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.S.; Larson, M.H.; Gilbert, L.A.; Doudna, J.A.; Weissman, J.S.; Arkin, A.P.; Lim, W.A. Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression. Cell 2013, 152, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Storz, G.; Vogel, J.; Wassarman, K.M. Regulation by Small RNAs in Bacteria: Expanding Frontiers. Mol. Cell 2011, 43, 880–891. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Vanderpool, C.K. Target activation by regulatory RNAs in bacteria. FEMS Microbiol. Rev. 2015, 39, 362–378. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, K.S.; Papenfort, K. Interplay of regulatory RNAs and mobile genetic elements in enteric pathogens. Mol. Microbiol. 2016, 101, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Oliva, G.; Sahr, T.; Buchrieser, C. Small RNAs, 5′ UTR elements and RNA-binding proteins in intracellular bacteria: Impact on metabolism and virulence. FEMS Microbiol. Rev. 2015, 39, 331–349. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Vogel, J. Small RNA functions in carbon metabolism and virulence of enteric pathogens. Front. Cell. Infect. Microbiol. 2014, 4, 91. [Google Scholar] [CrossRef] [PubMed]
- Kröger, C.; Colgan, A.; Srikumar, S.; Händler, K.; Sivasankaran, S.K.; Hammarlöf, D.L.; Canals, R.; Grissom, J.E.; Conway, T.; Hokamp, K.; et al. An Infection-Relevant Transcriptomic Compendium for Salmonella enterica Serovar Typhimurium. Cell Host Microbe 2013, 14, 683–695. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.; Papenfort, K.; Reinhardt, R.; Sharma, C.M.; Vogel, J. An atlas of Hfq-bound transcripts reveals 3’ UTRs as a genomic reservoir of regulatory small RNAs. EMBO J. 2012, 31, 4005–4019. [Google Scholar] [CrossRef] [PubMed]
- Srikumar, S.; Kröger, C.; Hébrard, M.; Colgan, A.; Owen, S.V.; Sivasankaran, S.K.; Cameron, A.D.S.; Hokamp, K.; Hinton, J.C.D. RNA-seq Brings New Insights to the Intra-Macrophage Transcriptome of Salmonella Typhimurium. PLoS Pathog. 2015, 11, e1005262. [Google Scholar] [CrossRef] [PubMed]
- Colgan, A.M.; Kröger, C.; Diard, M.; Hardt, W.-D.; Puente, J.L.; Sivasankaran, S.K.; Hokamp, K.; Hinton, J.C.D. The Impact of 18 Ancestral and Horizontally-Acquired Regulatory Proteins upon the Transcriptome and sRNA Landscape of Salmonella enterica serovar Typhimurium. PLoS Genet. 2016, 12, e1006258. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Arya, S.; Patil, S.D.; Sharma, A.; Jain, P.K.; Navani, N.K.; Pathania, R. Identification of Novel Regulatory Small RNAs in Acinetobacter baumannii. PLoS ONE 2014, 9, e93833. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.; Broach, W.H.; Lee, M.C.; Shaw, L.N. Towards the complete small RNome of Acinetobacter baumannii. Microb. Genomics 2016, 2. [Google Scholar] [CrossRef]
- Vogel, J.; Luisi, B.F. Hfq and its constellation of RNA. Nat. Rev. Microbiol. 2011, 9, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Sonnleitner, E.; Romeo, A.; Bläsi, U. Small regulatory RNAs in Pseudomonas aeruginosa. RNA Biol. 2012, 9, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Sonnleitner, E.; Bläsi, U. Regulation of Hfq by the RNA CrcZ in Pseudomonas aeruginosa Carbon Catabolite Repression. PLoS Genet. 2014, 10, e1004440. [Google Scholar] [CrossRef] [PubMed]
- Schilling, D.; Gerischer, U. The Acinetobacter baylyi hfq Gene Encodes a Large Protein with an Unusual C Terminus. J. Bacteriol. 2009, 191, 5553–5562. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.; Vogel, J. The role of Hfq in bacterial pathogens. Curr. Opin. Microbiol. 2010, 13, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Kakoschke, T.K.; Kakoschke, S.C.; Zeuzem, C.; Bouabe, H.; Adler, K.; Heesemann, J.; Rossier, O. The RNA Chaperone Hfq Is Essential for Virulence and Modulates the Expression of Four Adhesins in Yersinia enterocolitica. Sci. Rep. 2016, 6, 29275. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, J.R.; Grilo, A.M.; Guerreiro, S.I.; Sousa, S.A.; Leitão, J.H. Hfq: A multifaceted RNA chaperone involved in virulence. Future Microbiol. 2015, 11, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Sittka, A.; Pfeiffer, V.; Tedin, K.; Vogel, J. The RNA chaperone Hfq is essential for the virulence of Salmonella typhimurium. Mol. Microbiol. 2007, 63, 193–217. [Google Scholar] [CrossRef] [PubMed]
- Schilling, D.; Findeiß, S.; Richter, A.S.; Taylor, J.A.; Gerischer, U. The small RNA Aar in Acinetobacter baylyi: A putative regulator of amino acid metabolism. Arch. Microbiol. 2010, 192, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Massé, E.; Salvail, H.; Desnoyers, G.; Arguin, M. Small RNAs controlling iron metabolism. Curr. Opin. Microbiol. 2007, 10, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Oglesby-Sherrouse, A.G.; Murphy, E.R. Iron-responsive bacterial small RNAs: Variations on a theme. Metallomics 2013, 5, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Nwugo, C.C.; Gaddy, J.A.; Zimbler, D.L.; Actis, L.A. Deciphering the iron response in Acinetobacter baumannii: A proteomics approach. J. Proteomics 2011, 74, 44–58. [Google Scholar] [CrossRef] [PubMed]
- Yamada, J.; Yamasaki, S.; Hirakawa, H.; Hayashi-Nishino, M.; Yamaguchi, A.; Nishino, K. Impact of the RNA chaperone Hfq on multidrug resistance in Escherichia coli. J. Antimicrob. Chemother. 2010, 65, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Lalaouna, D.; Eyraud, A.; Chabelskaya, S.; Felden, B.; Massé, E. Regulatory RNAs Involved in Bacterial Antibiotic Resistance. PLoS Pathog. 2014, 10, e1004299. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Bak, G.; Lee, J.; Kim, K. Systematic analysis of the role of bacterial Hfq-interacting sRNAs in the response to antibiotics. J. Antimicrob. Chemother. 2015, 70, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, T.; Chou, M.Y.; Inouye, M. A unique mechanism regulating gene expression: Translational inhibition by a complementary RNA transcript (micRNA). Proc. Natl. Acad. Sci. USA 1984, 81, 1966–1970. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Pfeiffer, V.; Mika, F.; Lucchini, S.; Hinton, J.C.D.; Vogel, J. σE-dependent small RNAs of Salmonella respond to membrane stress by accelerating global omp mRNA decay. Mol. Microbiol. 2006, 62, 1674–1688. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, V.; Papenfort, K.; Lucchini, S.; Hinton, J.C.D.; Vogel, J. Coding sequence targeting by MicC RNA reveals bacterial mRNA silencing downstream of translational initiation. Nat. Struct. Mol. Biol. 2009, 16, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Udekwu, K.I.; Darfeuille, F.; Vogel, J.; Reimegård, J.; Holmqvist, E.; Wagner, E.G.H. Hfq-dependent regulation of OmpA synthesis is mediated by an antisense RNA. Genes Dev. 2005, 19, 2355–2366. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, V.; Sittka, A.; Tomer, R.; Tedin, K.; Brinkmann, V.; Vogel, J. A small non-coding RNA of the invasion gene island (SPI-1) represses outer membrane protein synthesis from the Salmonella core genome. Mol. Microbiol. 2007, 66, 1174–1191. [Google Scholar] [CrossRef] [PubMed]
- Guillier, M.; Gottesman, S.; Storz, G. Modulating the outer membrane with small RNAs. Genes Dev. 2006, 20, 2338–2348. [Google Scholar] [CrossRef] [PubMed]
- Parker, A.; Gottesman, S. Small RNA Regulation of TolC, the Outer Membrane Component of Bacterial Multidrug Transporters. J. Bacteriol. 2016, 198, 1101–1113. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.H.; Hyun, S.H.; Lee, J.Y.; Lee, J.S.; Lee, Y.S.; Kim, S.A.; Chae, J.-P.; Yoo, S.M.; Lee, J.C. Acinetobacter baumannii outer membrane protein A targets the nucleus and induces cytotoxicity. Cell. Microbiol. 2008, 10, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.H.; Lee, J.S.; Lee, Y.C.; Park, T.I.; Lee, J.C. Acinetobacter baumannii invades epithelial cells and outer membrane protein A mediates interactions with epithelial cells. BMC Microbiol. 2008, 8, 216. [Google Scholar] [CrossRef] [PubMed]
- Smani, Y.; Fàbrega, A.; Roca, I.; Sánchez-Encinales, V.; Vila, J.; Pachón, J. Role of OmpA in the Multidrug Resistance Phenotype of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2014, 58, 1806–1808. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Gene Name | Type | Function and Phenotype | Relevant References |
|---|---|---|---|
| adeRS | Two-component system | Directly or indirectly controls 579 genes. Regulates the expression of the AdeABC drug efflux pump. Required for biofilm formation biofilm. | [35,47] |
| bfmRS | Two-component system | Controls the expression of the K locus (exopolysaccharide production/capsule). Regulates expression of CsuA/BABCDE chaperone-usher secretion system, biofilm formation on abiotic surfaces. | [54,58,59] |
| gacSA | Two-component system | Directly or indirectly controls 674 genes, including virulence genes, biofilm and pili formation, resistance against human serum, motility, and metabolism of aromatic compounds (paa locus). | [60,62,63] |
| baeSR | Two-component system | Deletion results in reduced expression of AdeABC, AdeIJK, MacAB-TolC drug efflux pumps and increases susceptibility to tannic acid. | [50,51] |
| pmrAB | Two-component system | Mutations in pmrAB can confer resistance to colistin. | [65] |
| fur | Transcription factor | Transcriptional repressor of genes involved in iron homeostasis. | [66] |
| zur | Transcription factor | Transcriptional repressor of genes involved in zinc homeostasis. | [67,68] |
| abaR | Transcription factor | AHL-responsive transcription factor involved in quorum sensing and biofilm formation. | [69,70] |
| atfA | RNA polymerase binding protein | Deletion of atfA results in cell enlargement, loss of biofilm formation and increased susceptibility to antibiotics and ethanol. | [71] |
| hns | Nucleoid-associated protein | Deletion results in a hyper-motile phenotype, enhanced adherence to human pneumocytes and enhanced virulence in a nematode infection assay. | [72,73] |
| Gene Name | Gene Identifier | Product | ||
|---|---|---|---|---|
| ATCC17978 | ATCC17978-mff | AB5075-UW | ||
| gacA | A1S_0236 | ACX60_RS16900 | ABUW_RS17720 | Two-component response regulator |
| phoU | A1S_0256 | ACX60_RS16800 | ABUW_RS17620 | High affinity phosphate uptake transcriptional repressor |
| ihfA | A1S_0603 | ACX60_RS15110 | ABUW_RS15930 | Integration host factor subunit alpha |
| bfmR | A1S_0748 | ACX60_RS14635 | ABUW_RS15450 | Two-component response regulator |
| bfmS | A1S_0749 | ACX60_RS14630 | ABUW_RS15445 | Two-component sensor histidine kinase |
| - | A1S_2122 | ACX60_RS06950 | ABUW_RS07810 | LysR family transcriptional regulator |
| - | A1S_2537 | ACX60_RS04880 | ABUW_RS04985 | LysR family transcriptional regulator |
| Gene Name | Gene Identifier | Product | ||
|---|---|---|---|---|
| ATCC17978 | ATCC17978-mff | AB5075-UW | ||
| - | A1S_2082 | ACX60_RS11775 | ABUW_RS08030 | TetR family transcriptional regulator |
| - | A1S_2064 | ACX60_RS07270 | ABUW_RS08145 | LysR family transcriptional regulator |
| - | A1S_2042 | ACX60_RS07365 | ABUW_RS08240 | TetR family transcriptional regulator |
| - | A1S_1958 | ACX60_RS08015 | ABUW_RS08550 | AsnC family transcriptional regulator |
| - | A1S_1948 | ACX60_RS08070 | ABUW_RS08610 | MarR family transcriptional regulator |
| - | A1S_1874 | ACX60_RS08460 | ABUW_RS09000 | LysR family transcriptional regulator |
| - | not present | not present | ABUW_RS09560 | LysR family transcriptional regulator |
| - | not present | not present | ABUW_RS10070 | Fur family transcriptional regulator |
| alkR | A1S_1640 | ACX60_RS09755 | ABUW_RS10675 | AraC family transcriptional regulator |
| - | A1S_1578 | ACX60_RS10205 | ABUW_RS10870 | AraC family transcriptional regulator |
| arsR | A1S_1453 | ACX60_RS10865 | ABUW_RS11530 | ArsR family transcriptional regulator |
| - | A1S_1350 | ACX60_RS11395 | ABUW_RS12250 | TetR family transcriptional regulator |
| - | A1S_1330 | ACX60_RS11505 | ABUW_RS12365 | AraC family transcriptional regulator |
| soxR | A1S_1320 | ACX60_RS11550 | ABUW_RS12410 | Redox-sensitive transcriptional activator |
| - | A1S_0768 | ACX60_RS14535 | ABUW_RS15350 | LysR family transcriptional regulator |
| bfmS | A1S_0749 | ACX60_RS14630 | ABUW_RS15445 | Two-component sensor histidine kinase |
| - | A1S_0621 | ACX60_RS15010 | ABUW_RS15830 | Putative two-component response regulator |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kröger, C.; Kary, S.C.; Schauer, K.; Cameron, A.D.S. Genetic Regulation of Virulence and Antibiotic Resistance in Acinetobacter baumannii. Genes 2017, 8, 12. https://doi.org/10.3390/genes8010012
Kröger C, Kary SC, Schauer K, Cameron ADS. Genetic Regulation of Virulence and Antibiotic Resistance in Acinetobacter baumannii. Genes. 2017; 8(1):12. https://doi.org/10.3390/genes8010012
Chicago/Turabian StyleKröger, Carsten, Stefani C. Kary, Kristina Schauer, and Andrew D. S. Cameron. 2017. "Genetic Regulation of Virulence and Antibiotic Resistance in Acinetobacter baumannii" Genes 8, no. 1: 12. https://doi.org/10.3390/genes8010012
APA StyleKröger, C., Kary, S. C., Schauer, K., & Cameron, A. D. S. (2017). Genetic Regulation of Virulence and Antibiotic Resistance in Acinetobacter baumannii. Genes, 8(1), 12. https://doi.org/10.3390/genes8010012