
Sugar Allocation to Metabolic Pathways is Tightly Regulated and Affects the Virulence of Streptococcus mutans

Abstract
:1. Introduction
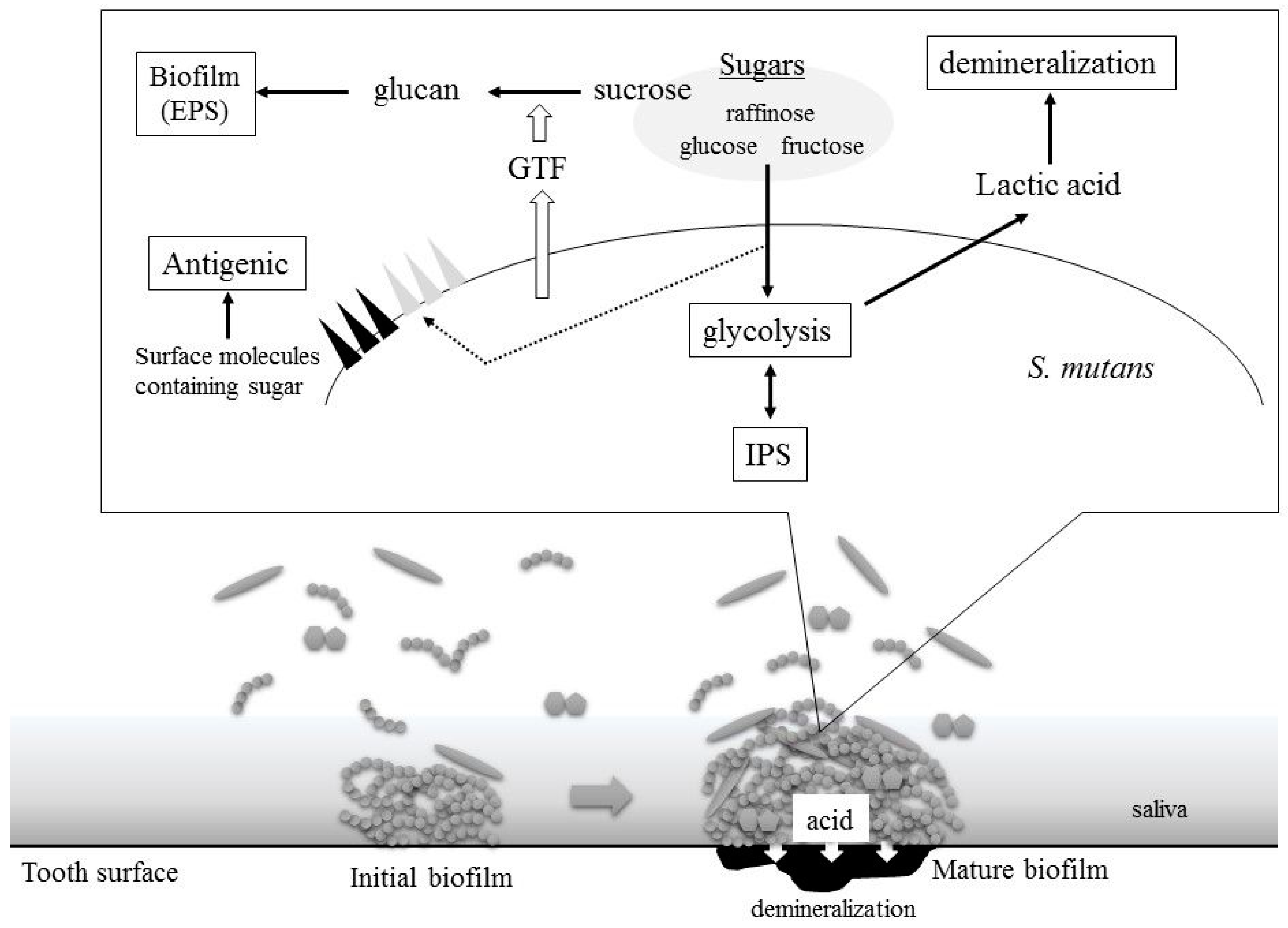
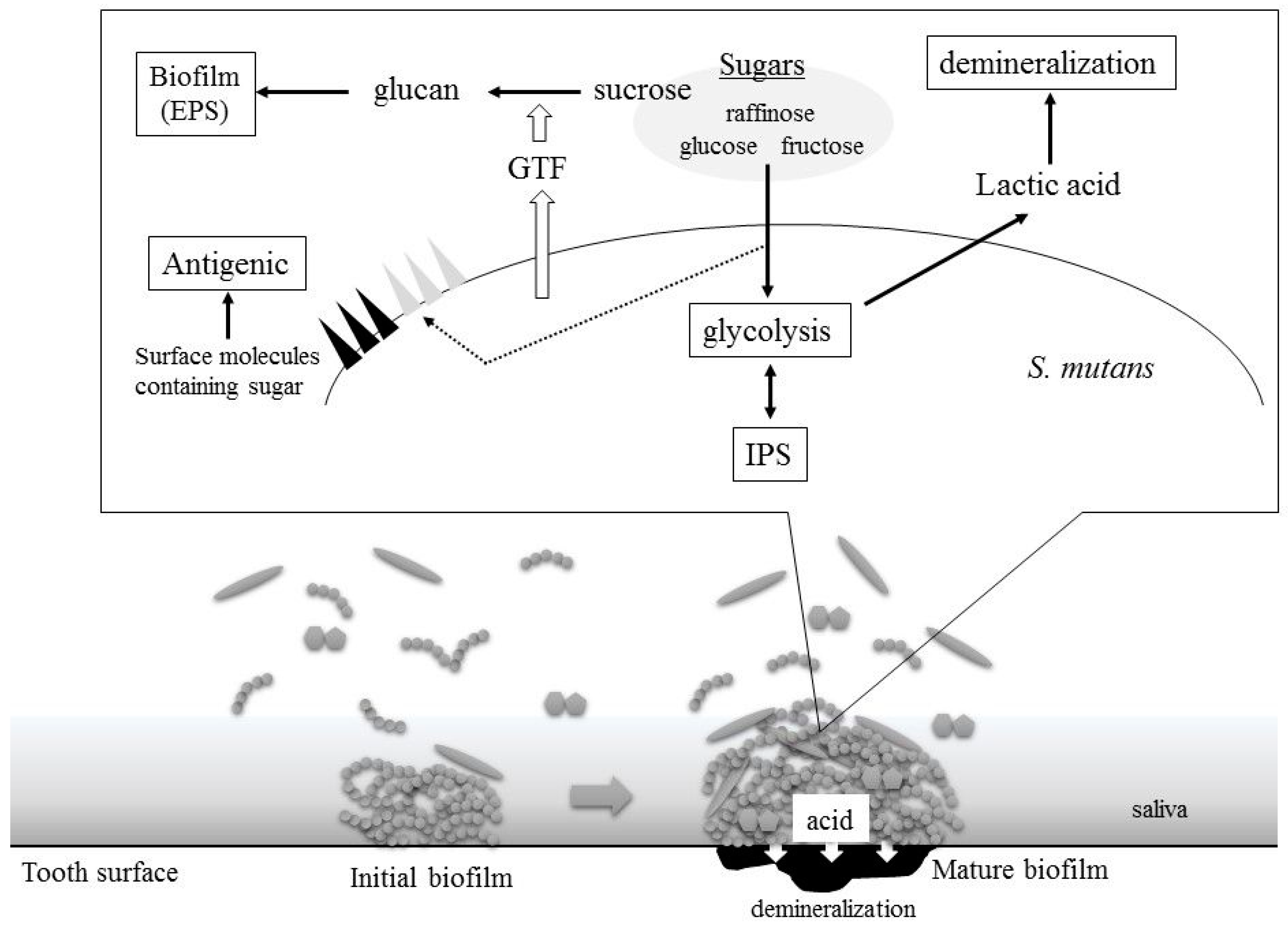
2. S. mutans Virulence Is Related to Sugar Metabolism
2.1. Acid Production
2.2. Biofilm Formation
2.3. Intercellular Polysaccharides Synthesis
2.4. Cell Surface Antigens: Serotype-Specific Carbohydrate Antigens and Lipoteichoic Acids
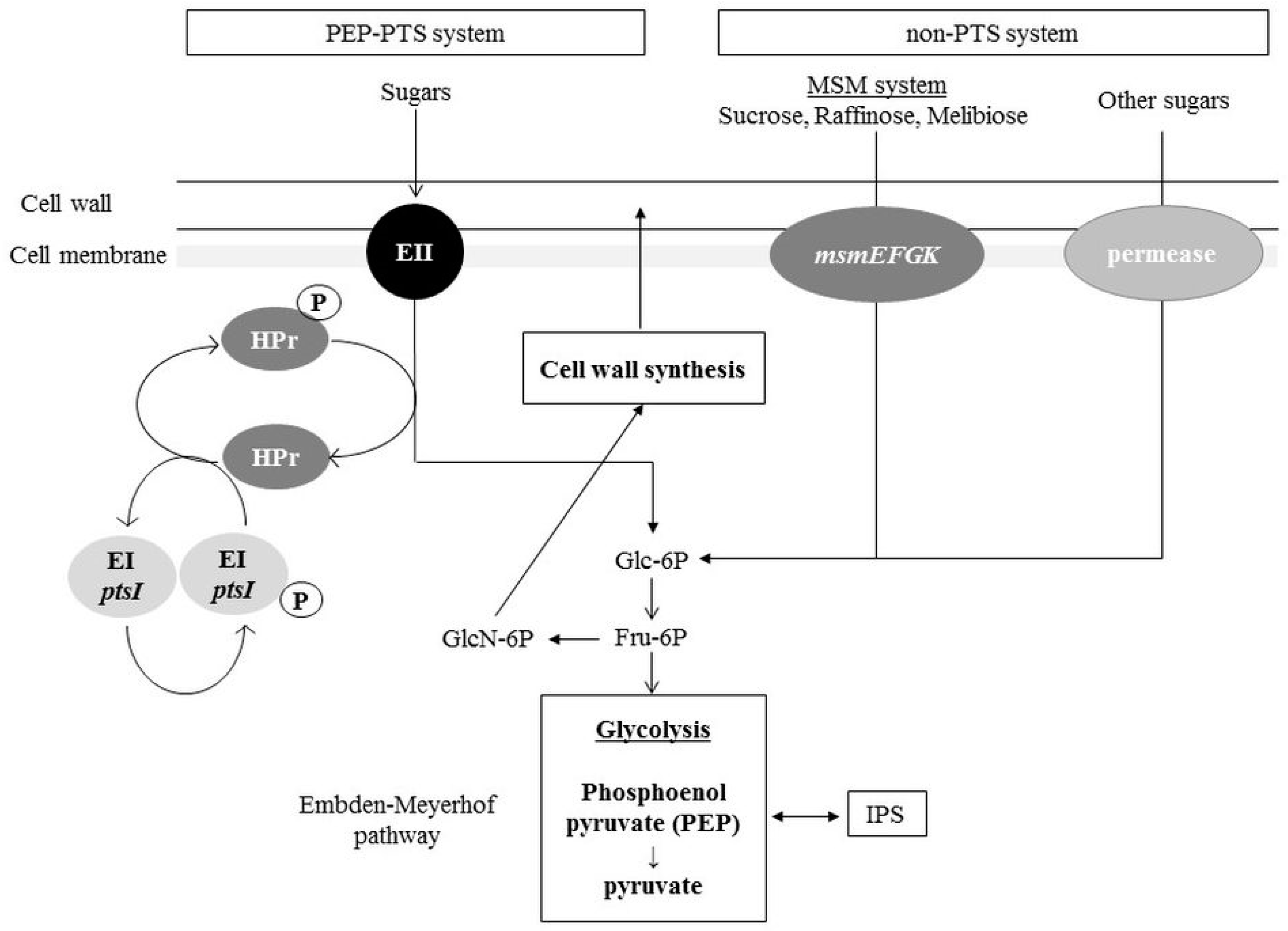
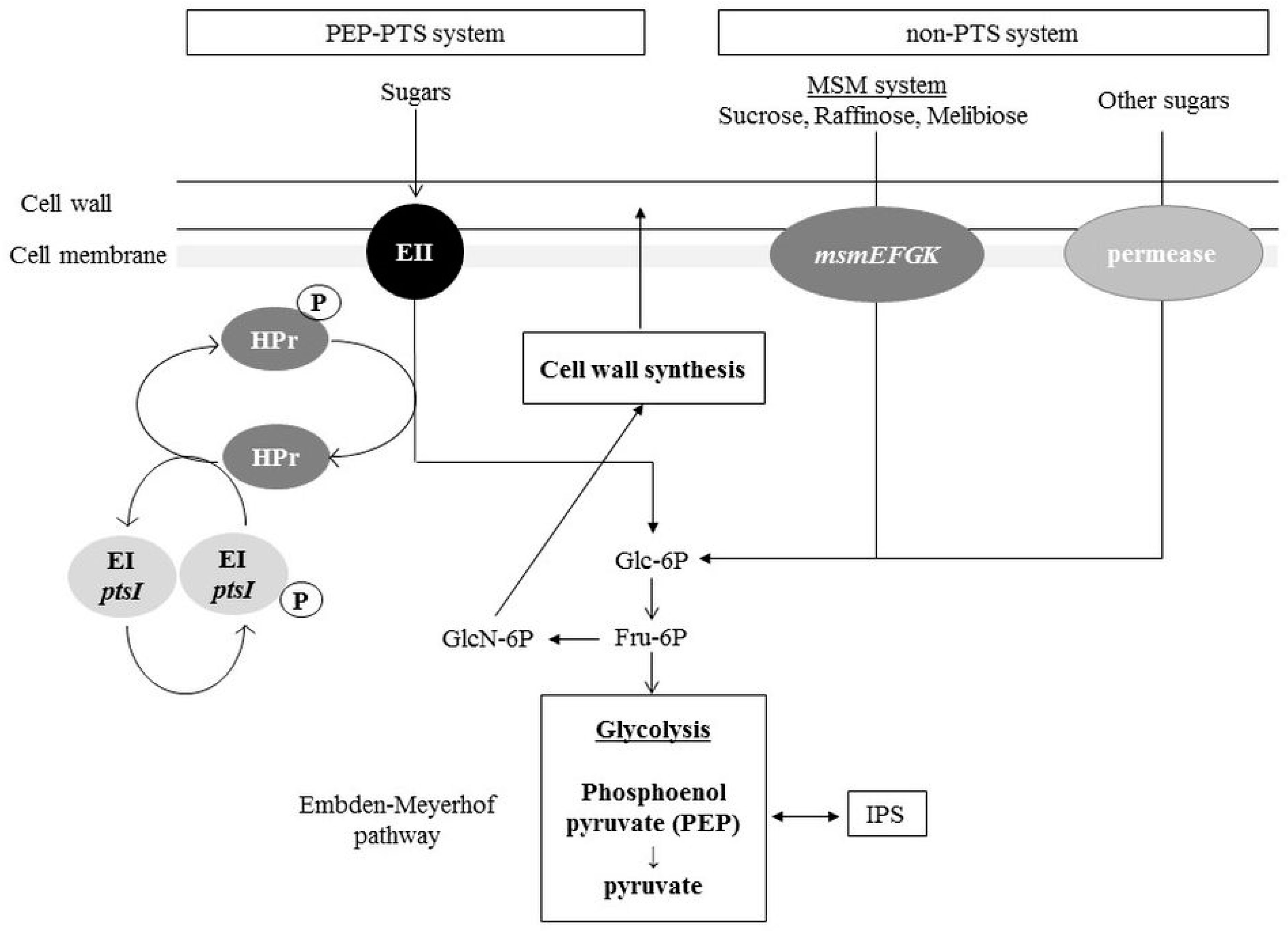
3. Distribution Pathways for Sugars and Amino Sugars into the Glycolysis Pathway and Cell Wall Synthesis Pathways in S. mutans
3.1. Sugar Incorporation: PTS and Non-PTS Systems
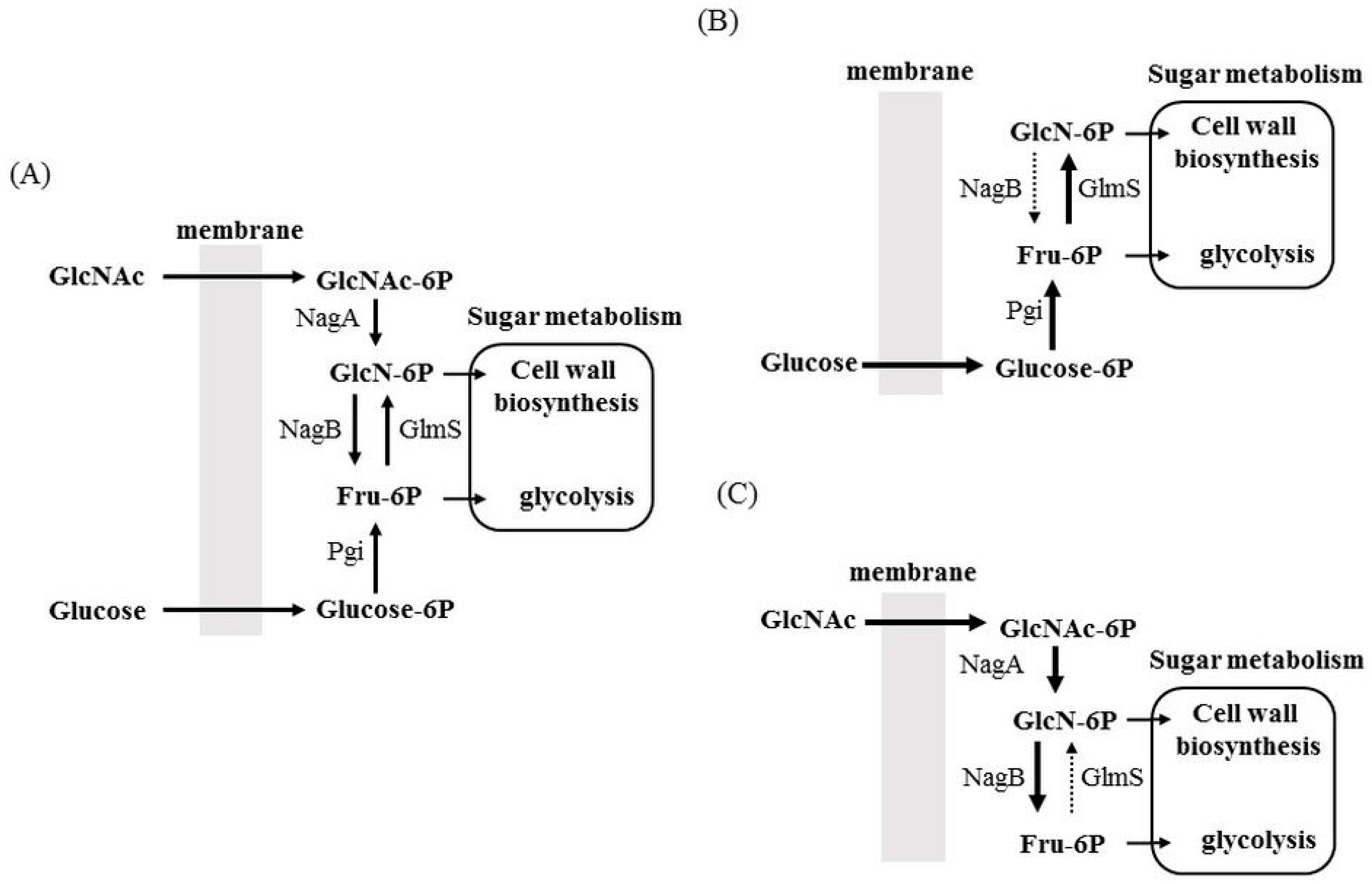
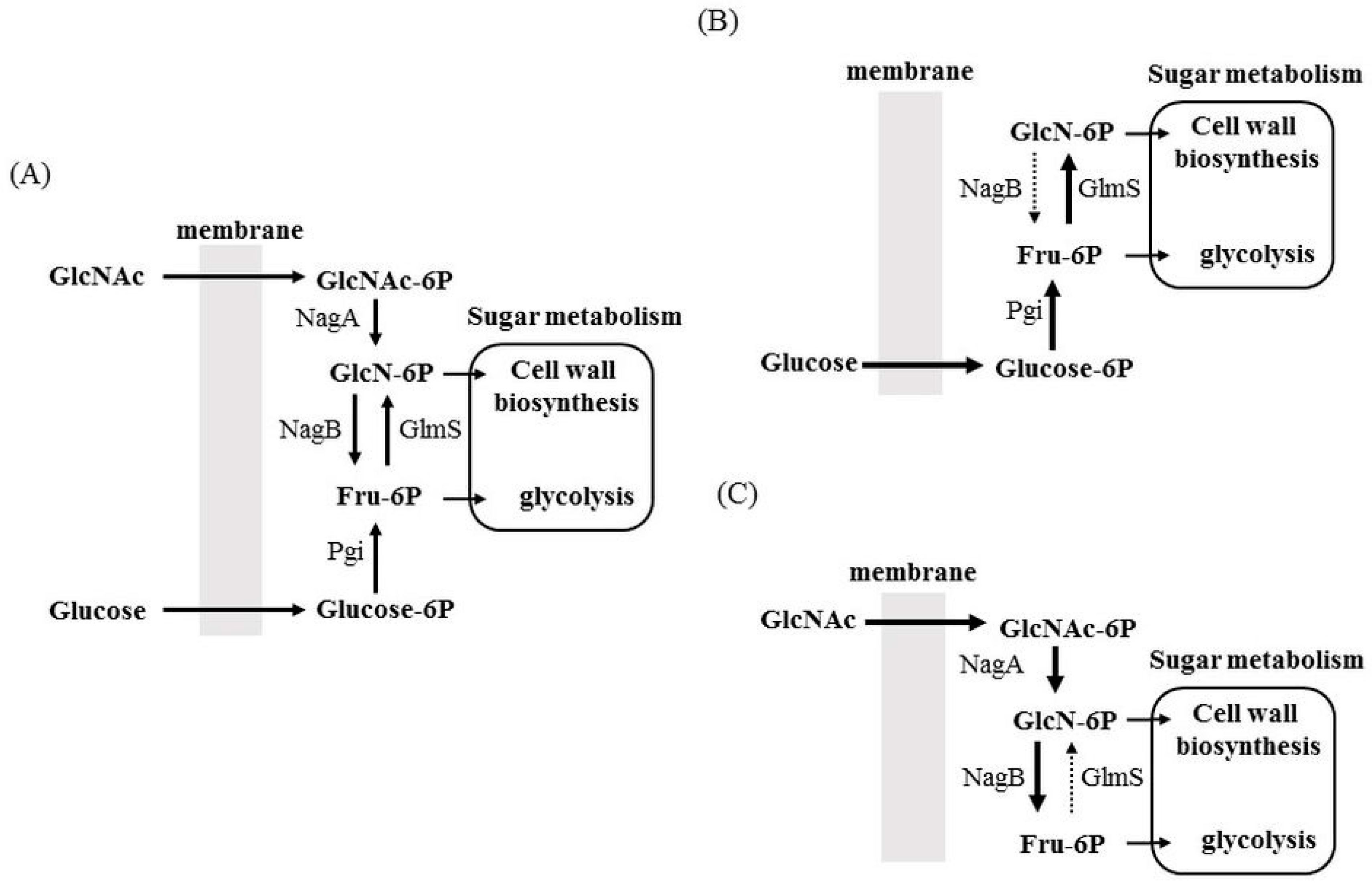
3.2. Allocation of Sugars and Amino Sugars to Glycolysis and Cell Wall Biosynthesis
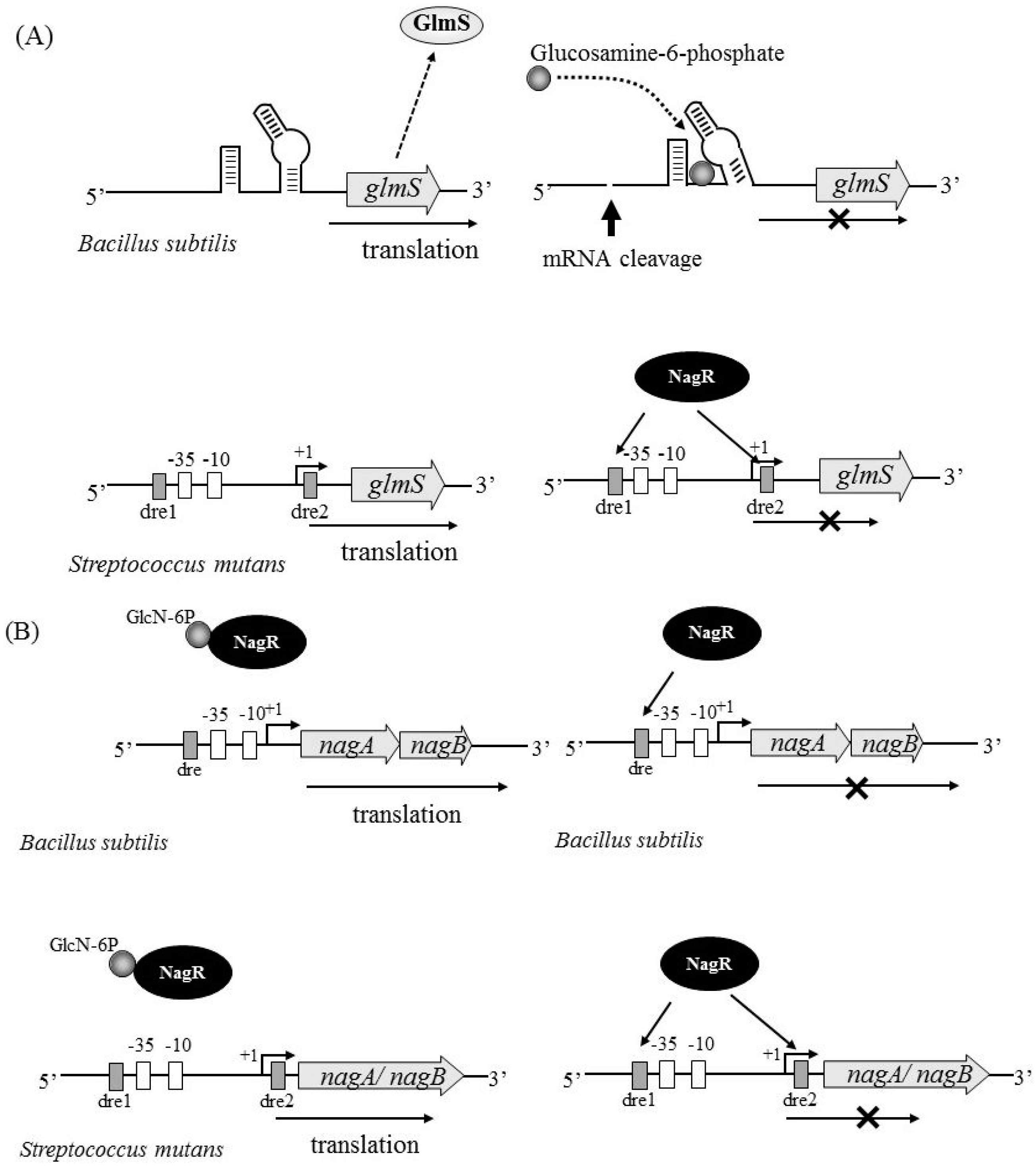
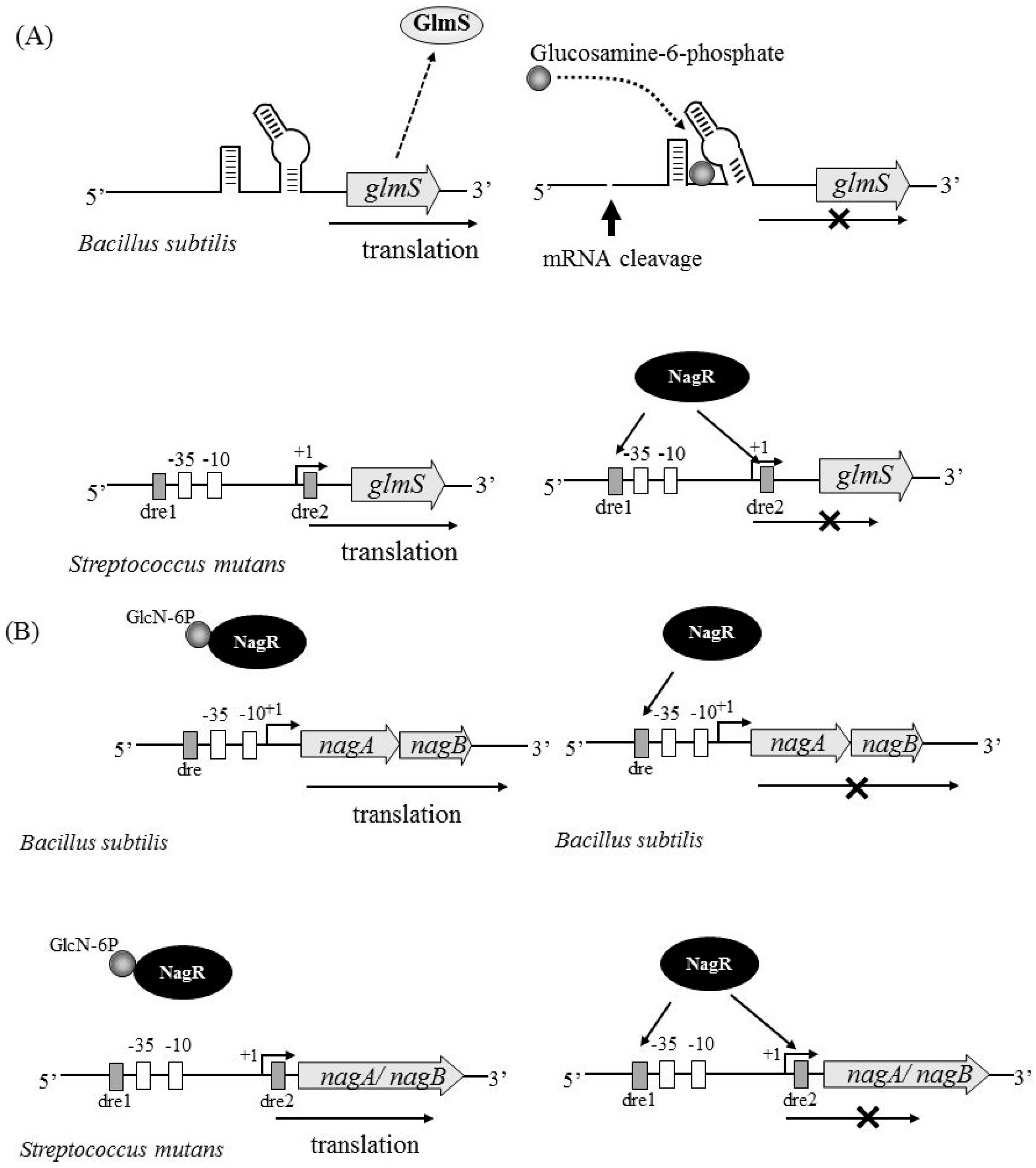
4. Factors Regulating nagB and glmS Expression
4.1. NagR
4.2. Carbon Catabolite Repression
5. Association of Carbohydrates with Virulence
6. Conclusions
Conflicts of Interest
References
- Loesche, W.J. Role of Streptococcus mutans in human dental decay. Microbiol. Rev. 1986, 50, 353–380. [Google Scholar] [PubMed]
- Hamada, S.; Slade, H.D. Biology, immunology, and cariogenicity of Streptococcus mutans. Microbiol. Rev. 1980, 44, 331–384. [Google Scholar] [PubMed]
- Van Houte, J. Role of micro-organisms in caries etiology. J. Dent. Res. 1994, 73, 672–681. [Google Scholar] [PubMed]
- Abbe, K.; Carlsson, J.; Takahashi-Abbe, S.; Yamada, T. Oxygen and the sugar metabolism in oral streptococci. Proc. Finn. Dent. Soc. 1991, 87, 477–487. [Google Scholar] [PubMed]
- Komatsuzawa, H.; Fujiwara, T.; Nishi, H.; Yamada, S.; Ohara, M.; McCallum, N.; Berger-Bächi, B.; Sugai, M. The gate controlling cell wall synthesis in Staphylococcus aureus. Mol. Microbiol. 2004, 53, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Neves, A.R.; Pool, W.A.; Kok, J.; Kuipers, O.P.; Santos, H. Overview on sugar metabolism and its control in Lactococcus lactis—The input from in vivo NMR. FEMS Microbiol. Rev. 2005, 29, 531–554. [Google Scholar] [PubMed]
- Ramos, A.; Boels, I.C.; de Vos, W.M.; Santos, H. Relationship between glycolysis and exopolysaccharide biosynthesis in Lactococcus lactis. Appl. Environ. Microbiol. 2001, 67, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Postma, P.W.; Lengeler, J.W.; Jacobson, G.R. Phosphoenolpyruvate: Carbohydrate phosphotransferase systems of bacteria. Microbiol. Rev. 1993, 57, 543–594. [Google Scholar] [PubMed]
- Vadeboncoeur, C.; Pelletier, M. The phosphoenolpyruvate: Sugar phosphotransferase system of oral streptococci and its role in the control of sugar metabolism. FEMS Microbiol. Rev. 1997, 19, 187–207. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, R.E.; Ferretti, J.J. The multiple-sugar metabolism (msm) gene cluster of Streptococcus mutans is transcribed as a single operon. FEMS Microbiol. Lett. 1996, 140, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Cvitkovitch, D.G.; Boyd, D.A.; Hamilton, I.R. Regulation of sugar transport via the multiple sugar metabolism operon of Streptococcus mutans by the phosphoenolpyruvate phosphotransferase system. J. Bacteriol. 1995, 177, 5704–5706. [Google Scholar] [CrossRef] [PubMed]
- Kawada-Matsuo, M.; Mazda, Y.; Oogai, Y.; Kajiya, M.; Kawai, T.; Yamada, S.; Miyawaki, S.; Oho, T.; Komatsuzawa, H. GlmS and NagB regulate amino sugar metabolism in opposing directions and affect Streptococcus mutans virulence. PLoS ONE 2012, 7, e33382. [Google Scholar] [CrossRef] [PubMed]
- Abranches, J.; Nascimento, M.M.; Zeng, L.; Browngardt, C.M.; Wen, Z.T.; Rivera, M.F.; Burne, R.A. CcpA regulates central metabolism and virulence gene expression in Streptococcus mutans. J. Bacteriol. 2008, 190, 2340–2349. [Google Scholar] [CrossRef] [PubMed]
- Seidl, K.; Goerke, C.; Wolz, C.; Mack, D.; Berger-Bächi, B.; Bischoff, M. Staphylococcus aureus CcpA affects biofilm formation. Infect. Immun. 2008, 76, 2044–2050. [Google Scholar] [CrossRef] [PubMed]
- Meier, S.; Goerke, C.; Wolz, C.; Seidl, K.; Homerova, D.; Schulthess, B.; Kormanec, J.; Berger-Bächi, B.; Bischoff, M. σB and the σB-dependent arlRS and yabJ-spoVG loci affect capsule formation in Staphylococcus aureus. Infect. Immun. 2007, 75, 4562–4571. [Google Scholar] [CrossRef] [PubMed]
- Banas, J.A.; Vickerman, M.M. Glucan-binding proteins of the oral streptococci. Crit. Rev. Oral Biol. Med. 2003, 14, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Kuramitsu, H.K. Molecular genetic analysis of the virulence of oral bacterial pathogens: An historical perspective. Crit. Rev. Oral Biol. Med. 2003, 14, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Colby, S.M.; Russell, R.R. Sugar metabolism by mutans streptococci. Soc. Appl. Bacteriol. Symp. Ser. 1997, 26, 80–88. [Google Scholar] [CrossRef]
- Germaine, G.R.; Harlander, S.K.; Leung, W.L.; Schachtele, C.F. Streptococcus mutans dextransucrase: Functioning of primer dextran and endogenous dextranase in water-soluble and water-insoluble glucan synthesis. Infect. Immun. 1977, 16, 637–648. [Google Scholar] [PubMed]
- Dewar, M.D.; Walker, G.J. Metabolism of the polysaccharides of human dental plaque. I. Dextranase activity of streptococci, and the extracellular polysaccharides synthesized from sucrose. Caries Res. 1975, 9, 21–35. [Google Scholar] [CrossRef] [PubMed]
- DiPersio, J.R.; Mattingly, S.J.; Higgins, M.L.; Shockman, G.D. Measurement of intracellular iodophilic polysaccharide in two cariogenic strains of Streptococcus mutans by cytochemical and chemical methods. Infect. Immun. 1974, 10, 597–604. [Google Scholar] [PubMed]
- Linzer, R.; Reddy, M.S.; Levine, M.J. Structural studies of the rhamnose-glucose polysaccharide antigen from Streptococcus sobrinus B13 and 6715-T2. Infect. Immun. 1985, 50, 583–585. [Google Scholar] [PubMed]
- Pritchard, D.G.; Gregory, R.L.; Michalek, S.M.; McGhee, J.R. Characterization of the serotype e polysaccharide antigen of Streptococcus mutans. Mol. Immunol. 1986, 23, 141–145. [Google Scholar] [CrossRef]
- Nakano, K.; Ooshima, T. Serotype classification of Streptococcus mutans and its detection outside the oral cavity. Future Microbiol. 2009, 4, 891–902. [Google Scholar] [CrossRef] [PubMed]
- Chia, J.S.; Lin, Y.L.; Lien, H.T.; Chen, J.Y. Platelet aggregation induced by serotype polysaccharides from Streptococcus mutans. Infect. Immun. 2004, 72, 2605–2617. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.W.; Baik, J.E.; Kang, S.S.; Yun, C.H.; Seo, D.G.; Han, S.H. Lipoteichoic acid of Streptococcus mutans interacts with Toll-like receptor 2 through the lipid moiety for induction of inflammatory mediators in murine macrophages. Mol. Immunol. 2014, 57, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.L.; Shirasu, S.; Daito, M.; Ohura, K. Streptococcus mutans lipoteichoic acid-induced apoptosis in cultured dental pulp cells from human deciduous teeth. Biochem. Biophys. Res. Commun. 2001, 281, 957–961. [Google Scholar] [CrossRef] [PubMed]
- Deutscher, J.; Aké, F.M.; Derkaoui, M.; Zébré, A.C.; Cao, T.N.; Bouraoui, H.; Kentache, T.; Mokhtari, A.; Milohanic, E.; Joyet, P. The bacterial phosphoenolpyruvate: Carbohydrate phosphotransferase system: Regulation by protein phosphorylation and phosphorylation-dependent protein-protein interactions. Microbiol. Mol. Biol. Rev. 2014, 78, 231–256. [Google Scholar] [CrossRef] [PubMed]
- Ajdić, D.; Pham, V.T. Global transcriptional analysis of Streptococcus mutans sugar transporters using microarrays. J. Bacteriol. 2007, 189, 5049–5059. [Google Scholar] [CrossRef] [PubMed]
- Moye, Z.D.; Burne, R.A.; Zeng, L. Uptake and metabolism of N-acetylglucosamine and glucosamine by Streptococcus mutans. Appl. Environ. Microbiol. 2014, 80, 5053–5067. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Xue, P.; Stanhope, M.J.; Burne, R.A. A galactose-specific sugar: Phosphotransferase permease is prevalent in the non-core genome of Streptococcus mutans. Mol. Oral Microbiol. 2013, 28, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Ajdic, D.; Chen, Z. A novel phosphotransferase system of Streptococcus mutans is responsible for transport of carbohydrates with α-1,3 linkage. Mol. Oral Microbiol. 2013, 28, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Honeyman, A.L.; Curtiss, R., 3rd. The mannitol-specific enzyme II (mtlA) gene and the mtlR gene of the PTS of Streptococcus mutans. Microbiology 2000, 146 Pt 7, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- Cote, C.K.; Cvitkovitch, D.; Bleiweis, A.S.; Honeyman, A.L. A novel β-glucoside-specific PTS locus from Streptococcus mutans that is not inhibited by glucose. Microbiology 2000, 146 Pt 7, 1555–1563. [Google Scholar] [CrossRef] [PubMed]
- Webb, A.J.; Homer, K.A.; Hosie, A.H. Two closely related ABC transporters in Streptococcus mutans are involved in disaccharide and/or oligosaccharide uptake. J. Bacteriol. 2008, 190, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Plumbridge, J.A.; Cochet, O.; Souza, J.M.; Altamirano, M.M.; Calcagno, M.L.; Badet, B. Coordinated regulation of amino sugar-synthesizing and -degrading enzymes in Escherichia coli K-12. J. Bacteriol. 1993, 175, 4951–4956. [Google Scholar] [CrossRef] [PubMed]
- Winkler, W.C.; Nahvi, A.; Roth, A.; Collins, J.A.; Breaker, R.R. Control of gene expression by a natural metabolite-responsive ribozyme. Nature 2004, 428, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Scott, W.G. Ribozymes. Curr. Opin. Struct. Biol. 2007, 17, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Tinsley, R.A.; Furchak, J.R.; Walter, N.G. Trans-acting glmS catalytic riboswitch: Locked and loaded. RNA 2007, 13, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Görke, B.; Vogel, J. Noncoding RNA control of the making and breaking of sugars. Genes Dev. 2008, 22, 2914–2925. [Google Scholar] [CrossRef] [PubMed]
- Bertram, R.; Rigali, S.; Wood, N.; Lulko, A.T.; Kuipers, O.P.; Titgemeyer, F. Regulon of the N-acetylglucosamine utilization regulator NagR in Bacillus subtilis. J. Bacteriol. 2011, 193, 3525–3536. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Burne, R.A. NagR Differentially Regulates the Expression of the glmS and nagAB Genes Required for Amino Sugar Metabolism by Streptococcus mutans. J. Bacteriol. 2015, 197, 3533–3544. [Google Scholar] [CrossRef] [PubMed]
- Görke, B.; Stülke, J. Carbon catabolite repression in bacteria: Many ways to make the most out of nutrients. Nat. Rev. Microbiol. 2008, 6, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Warner, J.B.; Lolkema, J.S. CcpA-dependent carbon catabolite repression in bacteria. Microbiol. Mol. Biol. Rev. 2003, 67, 475–490. [Google Scholar] [CrossRef] [PubMed]
- Deutscher, J. The mechanisms of carbon catabolite repression in bacteria. Curr. Opin. Microbiol. 2008, 11, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Deutscher, J.; Francke, C.; Postma, P.W. How phosphotransferase system-related protein phosphorylation regulates carbohydrate metabolism in bacteria. Microbiol. Mol. Biol. Rev. 2006, 70, 939–1031. [Google Scholar] [CrossRef] [PubMed]
- Abranches, J.; Candella, M.M.; Wen, Z.T.; Baker, H.V.; Burne, R.A. Different roles of EIIABMan and EIIGlc in regulation of energy metabolism, biofilm development, and competence in Streptococcus mutans. J. Bacteriol. 2006, 188, 3748–3756. [Google Scholar] [CrossRef] [PubMed]
- Saier, M.H., Jr. Regulatory interactions controlling carbon metabolism: An overview. Res. Microbiol. 1996, 147, 439–447. [Google Scholar] [CrossRef]
- Wen, Z.T.; Burne, R.A. Analysis of cis- and trans-acting factors involved in regulation of the Streptococcus mutans fructanase gene (fruA). J. Bacteriol. 2002, 184, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Fleming, E.; Camilli, A. ManLMN is a glucose transporter and central metabolic regulator in Streptococcus pneumoniae. Mol. Microbiol. 2016, 102, 467–487. [Google Scholar] [CrossRef] [PubMed]
- Kuramitsu, H.K. Virulence factors of mutans streptococci: Role of molecular genetics. Crit. Rev. Oral Biol. Med. 1993, 4, 159–176. [Google Scholar] [PubMed]
- Tovar, J.A.; Durán, C.; Rodríguez, A.; Jaramillo, L. Adhesion of salivary components to Streptococcus mutans peptides. Acta Odontol. Latinoam. 2006, 19, 53–58. [Google Scholar] [PubMed]
- Mattos-Graner, R.O.; Jin, S.; King, W.F.; Chen, T.; Smith, D.J.; Duncan, M.J. Cloning of the Streptococcus mutans gene encoding glucan binding protein B and analysis of genetic diversity and protein production in clinical isolates. Infect. Immun. 2001, 69, 6931–6941. [Google Scholar] [CrossRef] [PubMed]
- Shibata, Y.; Kawada, M.; Nakano, Y.; Toyoshima, K.; Yamashita, Y. Identification and characterization of an autolysin-encoding gene of Streptococcus mutans. Infect. Immun. 2005, 73, 3512–3520. [Google Scholar] [CrossRef] [PubMed]
- Jakubovics, N.S.; Strömberg, N.; van Dolleweerd, C.J.; Kelly, C.G.; Jenkinson, H.F. Differential binding specificities of oral streptococcal antigen I/II family adhesins for human or bacterial ligands. Mol. Microbiol. 2005, 55, 1591–1605. [Google Scholar] [CrossRef] [PubMed]
- Senadheera, M.D.; Guggenheim, B.; Spatafora, G.A.; Huang, Y.C.; Choi, J.; Hung, D.C.; Treglown, J.S.; Goodman, S.D.; Ellen, R.P.; Cvitkovitch, D.G. A VicRK signal transduction system in Streptococcus mutans affects gtfBCD, gbpB, and ftf expression, biofilm formation, and genetic competence development. J. Bacteriol. 2005, 187, 4064–4076. [Google Scholar] [CrossRef] [PubMed]
- Idone, V.; Brendtro, S.; Gillespie, R.; Kocaj, S.; Peterson, E.; Rendi, M.; Warren, W.; Michalek, S.; Krastel, K.; Cvitkovitch, D.; et al. Effect of an orphan response regulator on Streptococcus mutans sucrose-dependent adherence and cariogenesis. Infect. Immun. 2003, 71, 4351–4360. [Google Scholar] [CrossRef] [PubMed]
- Duque, C.; Stipp, R.N.; Wang, B.; Smith, D.J.; Höfling, J.F.; Kuramitsu, H.K.; Duncan, M.J.; Mattos-Graner, R.O. Downregulation of GbpB, a component of the VicRK regulon, affects biofilm formation and cell surface characteristics of Streptococcus mutans. Infect. Immun. 2011, 79, 786–796. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Expression | Phenotype | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Inactivation Gene | gtfB | gtfC | spaP | gbp | atlA | vicR | Biofilm Formation | Salivary Aggregation | Growth | |
| Glucose | GlcNAc | |||||||||
| glmS | Up a | Up | Up | Up | NC c | Up | Up | Up | No | Yes |
| nagB | Down b | Down | Down | Down | NC | Down | Down | Down | Yes | No |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawada-Matsuo, M.; Oogai, Y.; Komatsuzawa, H. Sugar Allocation to Metabolic Pathways is Tightly Regulated and Affects the Virulence of Streptococcus mutans. Genes 2017, 8, 11. https://doi.org/10.3390/genes8010011
Kawada-Matsuo M, Oogai Y, Komatsuzawa H. Sugar Allocation to Metabolic Pathways is Tightly Regulated and Affects the Virulence of Streptococcus mutans. Genes. 2017; 8(1):11. https://doi.org/10.3390/genes8010011
Chicago/Turabian StyleKawada-Matsuo, Miki, Yuichi Oogai, and Hitoshi Komatsuzawa. 2017. "Sugar Allocation to Metabolic Pathways is Tightly Regulated and Affects the Virulence of Streptococcus mutans" Genes 8, no. 1: 11. https://doi.org/10.3390/genes8010011
APA StyleKawada-Matsuo, M., Oogai, Y., & Komatsuzawa, H. (2017). Sugar Allocation to Metabolic Pathways is Tightly Regulated and Affects the Virulence of Streptococcus mutans. Genes, 8(1), 11. https://doi.org/10.3390/genes8010011