
CRISPR/Cas13-Based Anti-RNA Viral Approaches

 ,
, 
Abstract
1. Introduction
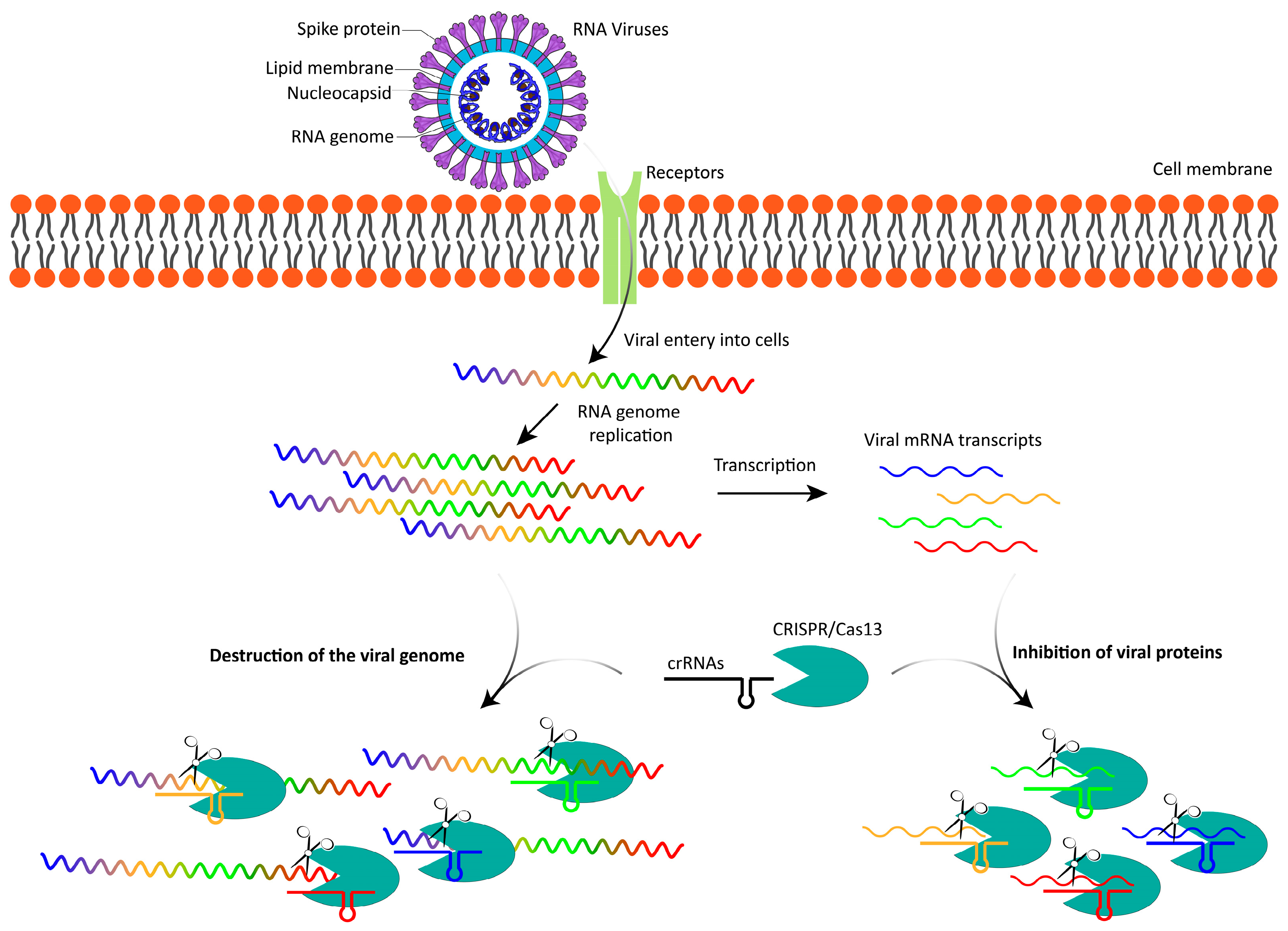
1.1. Overview of RNA Viruses
1.2. Current Antiviral Challenges
1.3. Emergence of Cas13
2. Cas13: Mechanism and Potential in Targeting RNA Viruses
2.1. Cas13 Discovery and Classification
2.2. Mechanism of Action
2.3. Advantages of Cas13 over Traditional Antiviral Strategies
3. Applications of Cas13 in Targeting RNA Viruses
3.1. SARS-CoV-2
3.2. HIV
3.3. Dengue, Influenza, and Other RNA Viruses
4. Challenges and Limitations of Cas13-Based Antiviral Therapies
4.1. Collateral Cleavage and Off-Target Effects
4.2. Delivery Challenges
4.3. Immunogenicity
4.4. Limited In Vivo Data
4.5. Regulatory and Clinical Approval Hurdles
4.6. Economic Considerations and Scalability
5. Future Directions and Ethical Considerations
5.1. Research Advancements
5.2. Ethical Considerations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| UTR | Untranslated Region |
| ORFs | Open Reading Frames |
| RdRPs | RNA-Dependent RNA Polymerases |
| PRRSV | Porcine Reproductive and Respiratory Syndrome Virus |
| AIVs | Avian Influenza Viruses |
| ART | Antiretroviral Therapy |
| CRISPR | Clustered Regularly Interspaced Short Palindromic Repeats |
| Cas | CRISPR/Associated Proteins |
| HEPN | Higher Eukaryotes and Prokaryotes Nucleotide-Binding |
| WHO | World Health Organization |
| HIV | Human Immunodeficiency Virus |
| HAART | Highly Active Antiretroviral Therapy |
| NRTI | Nucleoside Reverse Transcriptase Inhibitor |
| NNRTI | Non-nucleoside Reverse Transcriptase Inhibitor |
| INSTI | Integrase Strand Transfer Inhibitor |
| PI | Protease Inhibitor |
| LNPs | Lipid Nanoparticles |
| DENV | Dengue Virus |
| crRNA | CRISPR RNA |
| CPE | Cytopathic Effect |
| IAV | Influenza A Virus |
| ssRNA | Single-Stranded RNA |
| HCV | Hepatitis C Virus |
| IRES | Internal Ribosome Entry Site |
| HDV | Hepatitis Delta Virus |
| BoDV-1 | Borna Disease Virus |
| AAVs | Adeno-Associated Viruses |
| LNAs | Locked Nucleic Acids |
| FDA | Food and Drug Administration |
| EMA | European Medicines Agency |
| ICH | International Council for Harmonisation of Technical Requirements for Pharmaceuticals for Human Use |
| CDRH | Center for Devices and Radiological Health |
| EUA | Emergency Use Authorization |
| nAMD | Neovascular Age-related Macular Degeneration |
| CNV | Choroidal Neovascularization |
| aa | Amino acid |
| nt | Nucleotide |
References
- Bolduc, B.; Shaughnessy, D.P.; Wolf, Y.I.; Koonin, E.V.; Roberto, F.F.; Young, M. Identification of Novel Positive-Strand RNA Viruses by Metagenomic Analysis of Archaea-Dominated Yellowstone Hot Springs. J. Virol. 2012, 86, 5562–5573. [Google Scholar] [CrossRef] [PubMed]
- Callanan, J.; Stockdale, S.R.; Adriaenssens, E.M.; Kuhn, J.H.; Rumnieks, J.; Pallen, M.J.; Shkoporov, A.N.; Draper, L.A.; Ross, R.P.; Hill, C. Leviviricetes: Expanding and restructuring the taxonomy of bacteria-infecting single-stranded RNA viruses. Microb. Genom. 2021, 7, 000686. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Krupovic, M.; Agol, V.I. The Baltimore Classification of Viruses 50 Years Later: How Does It Stand in the Light of Virus Evolution? Microbiol. Mol. Biol. Rev. 2021, 85, e0005321. [Google Scholar] [CrossRef] [PubMed]
- Dreher, T.W. F UNCTIONS OF THE 3′-U NTRANSLATED R EGIONS OF P OSITIVE S TRAND RNA V IRAL G ENOMES. Annu. Rev. Phytopathol. 1999, 37, 151–174. [Google Scholar] [CrossRef] [PubMed]
- Strauss, E.G.; Strauss, J.H. RNA viruses: Genome structure and evolution. Curr. Opin. Genet. Dev. 1991, 1, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cousiño, N.; Maqueda, M.; Ambrona, J.; Zamora, E.; Esteban, R.; Ramírez, M. A New Wine Saccharomyces cerevisiae Killer Toxin (Klus), Encoded by a Double-Stranded RNA Virus, with Broad Antifungal Activity Is Evolutionarily Related to a Chromosomal Host Gene. Appl. Environ. Microbiol. 2011, 77, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Saberi, A.; Gulyaeva, A.A.; Brubacher, J.L.; Newmark, P.A.; Gorbalenya, A.E. A planarian nidovirus expands the limits of RNA genome size. PLoS Pathog. 2018, 14, e1007314. [Google Scholar] [CrossRef] [PubMed]
- Gorbalenya, A.E.; Enjuanes, L.; Ziebuhr, J.; Snijder, E.J. Nidovirales: Evolving the largest RNA virus genome. Virus Res. 2006, 117, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Nayak, B.; Kumar, S.; Collins, P.L.; Samal, S.K. Molecular characterization and complete genome sequence of avian paramyxovirus type 4 prototype strain duck/Hong Kong/D3/75. Virol. J. 2008, 5, 124. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, S.; Prasad, B.; Selvarajan, R. RNA Dependent RNA Polymerases: Insights from Structure, Function and Evolution. Viruses 2018, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Mattenberger, F.; Vila-Nistal, M.; Geller, R. Increased RNA virus population diversity improves adaptability. Sci. Rep. 2021, 11, 6824. [Google Scholar] [CrossRef] [PubMed]
- Sanjuán, R.; Domingo-Calap, P. Mechanisms of viral mutation. Cell Mol. Life Sci. 2016, 73, 4433–4448. [Google Scholar] [CrossRef] [PubMed]
- Duffy, S.; Shackelton, L.A.; Holmes, E.C. Rates of evolutionary change in viruses: Patterns and determinants. Nat. Rev. Genet. 2008, 9, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Graham, R.L.; Baric, R.S. Recombination, Reservoirs, and the Modular Spike: Mechanisms of Coronavirus Cross-Species Transmission. J. Virol. 2010, 84, 3134–3146. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-T.; Linster, M.; Mendenhall, I.H.; Su, Y.C.F.; Smith, G.J.D. Avian influenza viruses in humans: Lessons from past outbreaks. Br. Med. Bull. 2019, 132, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Gray, G.C.; Robie, E.R.; Studstill, C.J.; Nunn, C.L. Mitigating Future Respiratory Virus Pandemics: New Threats and Approaches to Consider. Viruses 2021, 13, 637. [Google Scholar] [CrossRef] [PubMed]
- Ishino, Y.; Krupovic, M.; Forterre, P. History of CRISPR-Cas from Encounter with a Mysterious Repeated Sequence to Genome Editing Technology. J. Bacteriol. 2018, 200, e00580-17. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.; Embden, J.D.A.v.; Gaastra, W.; Schouls, L.M. Identification of genes that are associated with DNA repeats in prokaryotes. Mol. Microbiol. 2002, 43, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Gasiunas, G.; Barrangou, R.; Horvath, P.; Siksnys, V. Cas9–crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proc. Natl. Acad. Sci. USA 2012, 109, E2579–E2586. [Google Scholar] [CrossRef] [PubMed]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-guided human genome engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Ledford, H. CRISPR treatment inserted directly into the body for first time. Nature 2020, 579, 185. [Google Scholar] [CrossRef] [PubMed]
- Abudayyeh, O.O.; Gootenberg, J.S.; Konermann, S.; Joung, J.; Slaymaker, I.M.; Cox, D.B.T.; Shmakov, S.; Makarova, K.S.; Semenova, E.; Minakhin, L.; et al. C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector. Science 2016, 353, aaf5573. [Google Scholar] [CrossRef] [PubMed]
- Shmakov, S.; Abudayyeh, O.O.; Makarova, K.S.; Wolf, Y.I.; Gootenberg, J.S.; Semenova, E.; Minakhin, L.; Joung, J.; Konermann, S.; Severinov, K.; et al. Discovery and Functional Characterization of Diverse Class 2 CRISPR-Cas Systems. Mol. Cell 2015, 60, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A.; et al. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Konermann, S.; Lotfy, P.; Brideau, N.J.; Oki, J.; Shokhirev, M.N.; Hsu, P.D. Transcriptome Engineering with RNA-Targeting Type VI-D CRISPR Effectors. Cell 2018, 173, 665–676.e614. [Google Scholar] [CrossRef] [PubMed]
- Smargon, A.A.; Cox, D.B.T.; Pyzocha, N.K.; Zheng, K.; Slaymaker, I.M.; Gootenberg, J.S.; Abudayyeh, O.A.; Essletzbichler, P.; Shmakov, S.; Makarova, K.S.; et al. Cas13b Is a Type VI-B CRISPR-Associated RNA-Guided RNase Differentially Regulated by Accessory Proteins Csx27 and Csx28. Mol. Cell 2017, 65, 618–630.e617. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.X.; Chong, S.; Zhang, H.; Makarova, K.S.; Koonin, E.V.; Cheng, D.R.; Scott, D.A. Cas13d Is a Compact RNA-Targeting Type VI CRISPR Effector Positively Modulated by a WYL-Domain-Containing Accessory Protein. Mol. Cell 2018, 70, 327–339.e325. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhou, Y.; Xiao, Q.; He, B.; Geng, G.; Wang, Z.; Cao, B.; Dong, X.; Bai, W.; Wang, Y.; et al. Programmable RNA editing with compact CRISPR-Cas13 systems from uncultivated microbes. Nat. Methods 2021, 18, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.; Huang, J.; Xiao, Q.; He, B.; Dong, X.; Liu, Y.; Yang, X.; Han, D.; Wang, Z.; Ying, W.; et al. High-fidelity Cas13 variants for targeted RNA degradation with minimal collateral effect. Nat. Biotechnol. 2021, 41, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Ghosh, A.; Chakravarti, R.; Singh, R.; Ravichandiran, V.; Swarnakar, S.; Ghosh, D. Cas13d: A New Molecular Scissor for Transcriptome Engineering. Front. Cell Dev. Biol. 2022, 10, 866800. [Google Scholar] [CrossRef] [PubMed]
- Abbott, T.R.; Dhamdhere, G.; Liu, Y.; Lin, X.; Goudy, L.; Zeng, L.; Chemparathy, A.; Chmura, S.; Heaton, N.S.; Debs, R.; et al. Development of CRISPR as an Antiviral Strategy to Combat SARS-CoV-2 and Influenza. Cell 2020, 181, 865–876.e812. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Liu, Y.; Nguyenla, X.H.; Abbott, T.R.; Han, M.; Zhu, Y.; Chemparathy, A.; Lin, X.; Chen, X.; Wang, H.; et al. Broad-spectrum CRISPR-mediated inhibition of SARS-CoV-2 variants and endemic coronaviruses in vitro. Nat. Commun. 2022, 13, 2766. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, E.L.; Vanover, D.; Bawage, S.S.; Tiwari, P.M.; Rotolo, L.; Beyersdorf, J.; Peck, H.E.; Bruno, N.C.; Hincapie, R.; Michel, F.; et al. Treatment of influenza and SARS-CoV-2 infections via mRNA-encoded Cas13a in rodents. Nat. Biotechnol. 2021, 39, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhou, J.; Wang, Q.; Wang, Y.; Kang, C. Rapid design and development of CRISPR-Cas13a targeting SARS-CoV-2 spike protein. Theranostics 2021, 11, 649–664. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Han, H.J.; Yu, J.; Kim, J.; Lee, G.H.; Yang, J.H.; Song, B.M.; Tark, D.; Choi, B.S.; Kang, S.M.; et al. Pseudoknot-targeting Cas13b combats SARS-CoV-2 infection by suppressing viral replication. Mol. Ther. 2023, 31, 1675–1687. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.; Andrade Dos Ramos, Z.; Vink, M.A.; Kroon, P.; Yu, Z.; Enjuanes, L.; Zuniga, S.; Berkhout, B.; Herrera-Carrillo, E. Efficient CRISPR-Cas13d-Based Antiviral Strategy to Combat SARS-CoV-2. Viruses 2023, 15, 686. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gao, X.; Kan, C.; Li, L.; Zhang, Y.; Gao, Y.; Zhang, S.; Zhou, L.; Zhao, H.; Li, M.; et al. CRISPR-Cas13d effectively targets SARS-CoV-2 variants, including Delta and Omicron, and inhibits viral infection. MedComm 2023, 4, e208. [Google Scholar] [CrossRef] [PubMed]
- Keng, C.T.; Yogarajah, T.; Lee, R.C.H.; Muhammad, I.B.H.; Chia, B.S.; Vasandani, S.R.; Lim, D.S.; Guo, K.; Wong, Y.H.; Mok, C.K.; et al. AAV-CRISPR-Cas13 eliminates human enterovirus and prevents death of infected mice. EBioMedicine 2023, 93, 104682. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, S.; Dong, X.; Li, Q.; Li, M.; Li, J.; Guo, Y.; Jin, X.; Zhou, Y.; Song, H.; et al. CRISPR-Cas13a Cleavage of Dengue Virus NS3 Gene Efficiently Inhibits Viral Replication. Mol. Ther. Nucleic Acids 2020, 19, 1460–1469. [Google Scholar] [CrossRef] [PubMed]
- Singsuksawat, E.; Onnome, S.; Posiri, P.; Suphatrakul, A.; Srisuk, N.; Nantachokchawapan, R.; Praneechit, H.; Sae-Kow, C.; Chidpratum, P.; Sa-Ngiamsuntorn, K.; et al. Potent programmable antiviral against dengue virus in primary human cells by Cas13b RNP with short spacer and delivery by VLP. Mol. Ther. Methods Clin. Dev. 2021, 21, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Basu, M.; Zurla, C.; Auroni, T.T.; Vanover, D.; Chaves, L.C.S.; Sadhwani, H.; Pathak, H.; Basu, R.; Beyersdorf, J.P.; Amuda, O.O.; et al. mRNA-encoded Cas13 can be used to treat dengue infections in mice. Nat. Microbiol. 2024, 9, 2160–2172. [Google Scholar] [CrossRef] [PubMed]
- Challagulla, A.; Schat, K.A.; Doran, T.J. In Vitro Inhibition of Influenza Virus Using CRISPR/Cas13a in Chicken Cells. Methods Protoc. 2021, 4, 40. [Google Scholar] [CrossRef] [PubMed]
- Chaves, L.C.S.; Orr-Burks, N.; Vanover, D.; Mosur, V.V.; Hosking, S.R.; Kumar, E.K.P.; Jeong, H.; Jung, Y.; Assumpcao, J.A.F.; Peck, H.E.; et al. mRNA-encoded Cas13 treatment of Influenza via site-specific degradation of genomic RNA. PLoS Pathog. 2024, 20, e1012345. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Zhao, F.; Sun, H.; Wang, Z.; Huang, Y.; Zhu, W.; Xu, F.; Mei, S.; Liu, X.; Zhang, D.; et al. CRISPR-Cas13a Inhibits HIV-1 Infection. Mol. Ther. Nucleic Acids 2020, 21, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.; Wilson, H.; Jayakumar, S.; Kulkarni, V.; Kulkarni, S. Efficient Inhibition of HIV Using CRISPR/Cas13d Nuclease System. Viruses 2021, 13, 1850. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Techakriengkrai, N.; Nedumpun, T.; Suradhat, S. Abrogation of PRRSV infectivity by CRISPR-Cas13b-mediated viral RNA cleavage in mammalian cells. Sci. Rep. 2020, 10, 9617. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.U.; Salman, H.M.; Khalid, M.F.; Khan, M.H.F.; Anwar, S.; Afzal, S.; Idrees, M.; Chaudhary, S.U. CRISPR-Cas13a mediated targeting of hepatitis C virus internal-ribosomal entry site (IRES) as an effective antiviral strategy. Biomed. Pharmacother. Biomed. Pharmacother. 2021, 136, 111239. [Google Scholar] [CrossRef] [PubMed]
- Adler, B.A.; Hessler, T.; Cress, B.F.; Lahiri, A.; Mutalik, V.K.; Barrangou, R.; Banfield, J.; Doudna, J.A. Broad-spectrum CRISPR-Cas13a enables efficient phage genome editing. Nat. Microbiol. 2022, 7, 1967–1979. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, S.; Ogawa, H.; Katoh, H.; Honda, T. Suppression of Borna Disease Virus Replication during Its Persistent Infection Using the CRISPR/Cas13b System. Int. J. Mol. Sci. 2024, 25, 3523. [Google Scholar] [CrossRef] [PubMed]
- Fareh, M.; Zhao, W.; Hu, W.; Casan, J.M.L.; Kumar, A.; Symons, J.; Zerbato, J.M.; Fong, D.; Voskoboinik, I.; Ekert, P.G.; et al. Reprogrammed CRISPR-Cas13b suppresses SARS-CoV-2 replication and circumvents its mutational escape through mismatch tolerance. Nat. Commun. 2021, 12, 4270. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.C.; Meng, H.Y.; Liu, H.H.; Ye, Q. Advances in laboratory detection methods and technology application of SARS-CoV-2. J. Med. Virol. 2022, 94, 1357–1365. [Google Scholar] [CrossRef] [PubMed]
- Schurr, F.; Tison, A.; Militano, L.; Cheviron, N.; Sircoulomb, F.; Rivière, M.-P.; Ribière-Chabert, M.; Thiéry, R.; Dubois, E. Validation of quantitative real-time RT-PCR assays for the detection of six honeybee viruses. J. Virol. Methods 2019, 270, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Joung, J.; Ladha, A.; Saito, M.; Segel, M.; Bruneau, R.; Huang, M.-l.W.; Kim, N.-G.; Yu, X.; Li, J.; Walker, B.D.; et al. Point-of-care testing for COVID-19 using SHERLOCK diagnostics. medRxiv 2020. [Google Scholar] [CrossRef]
- Rauch, J.N.; Valois, E.; Solley, S.C.; Braig, F.; Lach, R.S.; Audouard, M.; Ponce-Rojas, J.C.; Costello, M.S.; Baxter, N.J.; Kosik, K.S.; et al. A Scalable, Easy-to-Deploy Protocol for Cas13-Based Detection of SARS-CoV-2 Genetic Material. J. Clin. Microbiol. 2021, 59, e02402–e02420. [Google Scholar] [CrossRef] [PubMed]
- Arizti-Sanz, J.; Freije, C.A.; Stanton, A.C.; Petros, B.A.; Boehm, C.K.; Siddiqui, S.; Shaw, B.M.; Adams, G.; Kosoko-Thoroddsen, T.-S.F.; Kemball, M.E.; et al. Streamlined inactivation, amplification, and Cas13-based detection of SARS-CoV-2. Nat. Commun. 2020, 11, 5921. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.Y.; Worku, D. HIV vaccination: Navigating the path to a transformative breakthrough—A review of current evidence. Health Sci. Rep. 2024, 7, e70089. [Google Scholar] [CrossRef] [PubMed]
- Chun, T.W.; Stuyver, L.; Mizell, S.B.; Ehler, L.A.; Mican, J.A.; Baseler, M.; Lloyd, A.L.; Nowak, M.A.; Fauci, A.S. Presence of an inducible HIV-1 latent reservoir during highly active antiretroviral therapy. Proc. Natl. Acad. Sci. USA 1997, 94, 13193–13197. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhou, T.; Zhang, Y.; Luo, S.; Chen, H.; Chen, D.; Li, C.; Li, W. The reservoir of latent HIV. Front. Cell Infect. Microbiol. 2022, 12, 945956. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.Y.; Wu, H.Y.; Yarla, N.S.; Xu, B.; Ding, J.; Lu, T.R. HAART in HIV/AIDS Treatments: Future Trends. Infect. Disord. Drug Targets 2018, 18, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Ebina, H.; Misawa, N.; Kanemura, Y.; Koyanagi, Y. Harnessing the CRISPR/Cas9 system to disrupt latent HIV-1 provirus. Sci. Rep. 2013, 3, 2510. [Google Scholar] [CrossRef] [PubMed]
- Letchumanan, P.; Theva Das, K. The role of genetic diversity, epigenetic regulation, and sex-based differences in HIV cure research: A comprehensive review. Epigenetics Chromatin 2025, 18, 1. [Google Scholar] [CrossRef] [PubMed]
- Das, A.T.; Binda, C.S.; Berkhout, B. Elimination of infectious HIV DNA by CRISPR-Cas9. Curr. Opin. Virol. 2019, 38, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Nouri, R.; Jiang, Y.; Politza, A.J.; Liu, T.; Greene, W.H.; Zhu, Y.; Nunez, J.J.; Lian, X.; Guan, W. STAMP-Based Digital CRISPR-Cas13a for Amplification-Free Quantification of HIV-1 Plasma Viral Loads. ACS Nano 2023, 17, 10701–10712. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Su, B.; Yang, L.; Kou, Z.; Wu, H.; Zhang, T.; Liu, L.; Han, Y.; Niu, M.; Sun, Y.; et al. Highly sensitive and rapid point-of-care testing for HIV-1 infection based on CRISPR-Cas13a system. BMC Infect. Dis. 2023, 23, 627. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Chen, M.; Chen, Y.; Jing, X.; Zhang, N.; Zhou, X.; Li, X.; Long, G.; Hao, P. Targeted inhibition of Zika virus infection in human cells by CRISPR-Cas13b. Virus Res. 2022, 312, 198707. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Fan, Z.; Zhang, X.; Xu, L.; Cao, Y.; Pan, Z.; Mo, Y.; Gao, Y.; Zheng, S.; Huang, J.; et al. CRISPR/Cas13a-Assisted accurate and portable hepatitis D virus RNA detection. Emerg. Microbes Infect. 2023, 12, 2276337. [Google Scholar] [CrossRef] [PubMed]
- Madigan, V.; Zhang, F.; Dahlman, J.E. Drug delivery systems for CRISPR-based genome editors. Nat. Rev. Drug Discov. 2023, 22, 875–894. [Google Scholar] [CrossRef] [PubMed]
- Paunovska, K.; Loughrey, D.; Dahlman, J.E. Drug delivery systems for RNA therapeutics. Nat. Rev. Genet. 2022, 23, 265–280. [Google Scholar] [CrossRef] [PubMed]
- CRISPR/cas13-Mediated RNA TarGeting THerapy for the Treatment of Neovascular Age-Related Macular Degeneration Investigator-Initiated Trial (SIGHT-I). Available online: https://clinicaltrials.gov/study/NCT06031727?cond=Cas06031713&rank=06031721 (accessed on 4 September 2023).
- Open-Label Dose-Escalation Study for CRISPR/cas13-Rna TargetInG THerapy for the Treatment of Neovascular Age-Related Macular Degeneration in Phase I Trial (BRIGHT). Available online: https://clinicaltrials.gov/study/NCT06623279?cond=Cas06623213&rank=06623272 (accessed on 1 April 2025).
- Pougnet, R.; Derbez, B.; Troadec, M.B. Mapping the ‘Ethical’ Controversy of Human Heritable Genome Editing: A Multidisciplinary Approach. Asian Bioeth. Rev. 2023, 15, 189–204. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Ghosh, S.; Raghunath, M.; Bhaskar, R.; Sinha, J.K. Balancing potential benefits and ethical considerations of gene editing. Lancet 2023, 401, 2109–2110. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| CRISPR /Cas System | Mechanism of Action | Strengths | Weaknesses |
|---|---|---|---|
| Cas9 | Cas9 contains many domains, among which the HNH and RuvC domains are particularly important. When the crRNA synthesized by crRNA and tracrRNA binds to Cas9, the other domains will work together to assist crRNA in identifying the PAM site in the target gene. After locating the target gene, the HNH and RuvC domains will simultaneously cut the double-stranded strands of the target gene. | crRNA is synthesized from crRNA and tracrRNA, which is more accurate for locating the PAM site of the target gene. | 1. Cas9 has no trans-cleavage activity, and in vitro detection is difficult. 2. Cas protein can cut dsDNA, but the range is relatively limited. |
| Cas12 | The Cas12 system does not require tracrRNA and RNaseIII to process crRNA, but the nuclease in the WED domain of the Cas12 protein directly catalyzes the synthesis of crRNA. The processed crRNA directly binds to the Cas12 protein to locate the target gene and is sheared by the RuvC domain. | 1. The Cas12 system only needs crRNA to bind to the Cas12 protein for detection, and the test is easy. 2. The Cas12 protein has trans-cleavage activity and can be independently detected in vitro. | 1. The PAM site sequence of the Cas12 system is TTTV (V is A, G, or C), and crRNA design is difficult. 2. The Cas12 protein can only cut dsDNA and ssDNA and cannot recognize and detect RNA. |
| Cas13 | The Cas13 system does not require tracrRNA. The pre-crRNA is processed into mature crRNA by the HEPN-2 domain, which then recognizes the target gene after binding to the crRNA and is cut by the HEPN domain in the Cas13 protein. | 1. The Cas13 system has a PFS sequence that is equivalent to the PAM sequence. This sequence is composed of A, U, or C at the 3′ end of the spacer sequence, which increases the fault tolerance of the Cas13 protein. 2. The Cas13 protein can tolerate mismatches between crRNA and bases outside the seed region of the target sequence, which has almost no effect on the cutting efficiency. 3. The Cas13 protein has trans-cutting activity and can be tested in vitro alone. | Cas13 protein can only mechanically cut RNA. If DNA detection is required, DNA must be converted to RNA in vitro for detection, which increases the risk of nucleic acid contamination during the experiment. |
| Subtype | VI-A | VI-B | VI-C | VI-D | Cas13X | hfCas13X | Cas13Y |
|---|---|---|---|---|---|---|---|
| Cas effector | Cas13a | Cas13b | Cas13c | Cas13d (CasRx) | Cas13X | hfCas13X | Cas13Y |
| Accessory proteins | Cas1 and Cas2 | Csx27/28 | Unknown | Cas1 and Cas2 | None | None | None |
| Homologs/complexes | Leptotrichia spp. Lahnospiraceae bacterium | Bergeyella zoohelcum, Prevotella buccae | Fusobacterium perfoetens | Leptotrichia shahii | Metagenomic source | Engineered Cas13X | Metagenomic source |
| Size | ~1250 aa | ~1150 aa | ~1120 aa | ~930 aa | ~775 aa | ~775 aa | ~775 aa |
| Architecture | REC and NUC lobe | Pyramidal | Uncharacterized | REC and NUC lobe | Compact single-lobe | Compact single-lobe | Compact single-lobe |
| Pre-crRNA processing site | Helical-1 and HEPN-2 | RRI-2 (Lid) | Unknown | HEPN-2 | HEPN domain | HEPN domain | HEPN domain |
| Pre-crRNA mechanism | Acid–base | Acid–base | Unknown | Acid–base | Not reported | Not reported | Not reported |
| ssRNA cleavage preferences | U- and A-rich | Pyrimidine (U) | Unknown | U | U and others | Reduced off-target | Likely U-rich |
| Protospacer-flanking sequence | 5′ non-G | 5′ non-C, 3′ NAN | No restriction | No restriction | Unknown | Unknown | Unknown |
| Small accessory proteins | None | Csx27/28 | WYL-domain | None | None | None | None |
| Orientation (repeat→spacer) | 5′→3′ | 3′→5′ | Unknown | 5′→3′ | 5′→3′ | 5′→3′ | 5′→3′ |
| Repeat length (mature) | 27–32 nt | 36–88 nt | Unknown | 30 nt | ~28–30 nt | ~28–30 nt | ~28–30 nt |
| Repeat architecture | Stem-loop | Distorted stem-loop | Unknown | Stem-loop | Simple stem-loop | Simple stem-loop | Simple stem-loop |
| Recognition mechanism | Sequence + structure | Structure | Unknown | Sequence + structure | Likely structural | Enhanced fidelity | Likely structural |
| Spacer mismatch-sensitive | Seed region, HEPN switch | Central | Unknown | Internal, 3′ | Limited data | High specificity | Limited data |
| Reference | Abudayyeh et al. [23] | Smargon et al. [27] | – | Konermann et al. [26] | Xu et al., 2021 [29] | Xu et al., 2021 [29] | Xu et al., 2021 [29] |
| Virus Type | Publication | Strategy | Efficacy |
|---|---|---|---|
| SARS-CoV-2 | [32] | Cas13d with DNA constructs co-transfected into lung epithelial cells (A549). | GFP fluorescence reduced by 86%, mRNA expression by 83%. |
| [33] | Cas13d in Vero E6 cells targeting Alpha, Beta, and Omicron variants using lentivirus delivery. | Viral titer inhibited by ~95% at 24 hpi, up to 97% with combined crRNAs. | |
| [34] | Cas13a mRNA and crRNA delivered to hamsters via nebulizer; tested in vitro and in vivo. | 72% plaque reduction in vitro; 57% lung viral RNA reduction and abrogated weight loss in hamsters. | |
| [35] | Cas13a targeting spike protein in HepG2 and AT2 cells via lentivirus delivery. | Silencing efficiency >99.9% by qPCR. | |
| [36] | Cas13b targeting pseudoknot in hACE2 transgenic mice and in vitro. | Reduced viral replication by 99%; spike protein expression significantly lowered. | |
| [37] | Cas13d with 29 crRNAs targeting conserved regions of SARS-CoV-2 genome; tested in replicon and reporter assays. | Efficient suppression of SARS-CoV-2 replicon; crRNAs also inhibited SARS-CoV, showing broad antiviral potential. | |
| [38] | Cas13d with 50 crRNAs targeting conserved regions of ORF1ab region (NSP13 and NSP14) of SARS-CoV-2 genome; tested coronaviral nucleocapsid protein (NP) levels by Western blotting. | Achieved >99% silencing efficiency on nucleocapsid transcripts in human cells which are infected with coronavirus 2. | |
| [36] | Cas13b with 12 crRNAs targeting the pseudoknot site upstream of ORF1b; tested in replicon and infectivity in Vero E6 and hACE2 transgenic mice. | Reduced expression of the spike protein and attenuated viral replication by 99%. | |
| Human Enterovirus | [39] | AAV-delivered Cas13 system targeting conserved viral RNA sequences designed via bioinformatics pipeline. | Reduced viral titers by >99.99% in vitro; prophylactic and therapeutic inhibition in mice prevented death in lethal challenge. |
| Dengue | [40] | Cas13a/crRNA complex transfected into DENV-2-infected Vero cells. | RNA copy inhibition ~95% and plaque reduction ~84% at day 3. |
| [41] | Cas13b RNP delivered via virus-like particles (VLP) to human primary cells. | Efficient suppression of dengue virus infection. | |
| [42] | LNP-formulated mRNA-encoded Cas13a and crRNA applied in DENV infected mice. | Improved the survival of all infected animals and significantly decreased serum viral titers, with no collateral cleavage observed. | |
| Influenza A (IAV) | [34] | Cas13a mRNA and crRNA were tested in mouse model. | 94% mRNA expression reduction and significant protection in treated animals. |
| [43] | Cas13a in chicken fibroblast cells targeting IAV strains WSN and PR8. | Two- to fourfold reduction in infection by plaque assays. | |
| [44] | mRNA-encoded LbuCas13a, along with two crRNAs targeting H1N1 and H3N2 strains, was tested in A549 cells and hamsters. | RNA degradation was observed when delivered 24 h post-infection in vitro, along with a 1–2 log reduction in viral titers in hamsters | |
| HIV | [45] | Cas13a targets HIV in HEK293T cells. | viral replication of HIV was suppressed and the RNA levels were reduced. |
| [46] | RfxCas13d with HIV-specific crRNAs targeting in primary CD4+ T cells and reactivated latent HIV. | Inhibited HIV-1 replication. | |
| PRRSV | [47] | Cas13b targeting ORF5 and ORF7 in cell culture. | Gene knockdown and significant GFP fluorescence reduction. |
| Hepatitis C Virus | [48] | Cas13a targeting internal ribosomal entry site (IRES) in Huh7.5 cells. | Demonstrated 85% inhibition of luciferase activity. |
| Phage | [49] | Versatile Cas13a for phage genome editing. | Effective phage targeting and genome editing. |
| Borna | [50] | pspCas13b was used in BoDV-1 infected 293T cells. | Suppressed BoDV-1 in both acute and persistent infections. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, X.; Li, J.; Cui, B.; Wu, J.; Toischer, K.; Hasenfuß, G.; Xu, X. CRISPR/Cas13-Based Anti-RNA Viral Approaches. Genes 2025, 16, 875. https://doi.org/10.3390/genes16080875
Tan X, Li J, Cui B, Wu J, Toischer K, Hasenfuß G, Xu X. CRISPR/Cas13-Based Anti-RNA Viral Approaches. Genes. 2025; 16(8):875. https://doi.org/10.3390/genes16080875
Chicago/Turabian StyleTan, Xiaoying, Juncong Li, Baolong Cui, Jingjing Wu, Karl Toischer, Gerd Hasenfuß, and Xingbo Xu. 2025. "CRISPR/Cas13-Based Anti-RNA Viral Approaches" Genes 16, no. 8: 875. https://doi.org/10.3390/genes16080875
APA StyleTan, X., Li, J., Cui, B., Wu, J., Toischer, K., Hasenfuß, G., & Xu, X. (2025). CRISPR/Cas13-Based Anti-RNA Viral Approaches. Genes, 16(8), 875. https://doi.org/10.3390/genes16080875