
Transcriptomic Insights into GABA Accumulation in Tomato via CRISPR/Cas9-Based Editing of SlGAD2 and SlGAD3




Abstract
1. Introduction
2. Materials and Methods
2.1. Development of CRISPR/Cas9-Edited Tomato Lines and Fruit Tissue Sampling
2.2. Analysis of GABA and Total Amino Acids
2.3. RNA Extraction and Transcriptome Sequencing
2.4. Quantitative Reverse Transcription PCR (qRT-PCR) Analysis
2.5. Statistical Analysis
3. Results
3.1. Generation and Molecular Characterization of SlGAD2 and SlGAD3 CRISPR/Cas9-Edited Lines
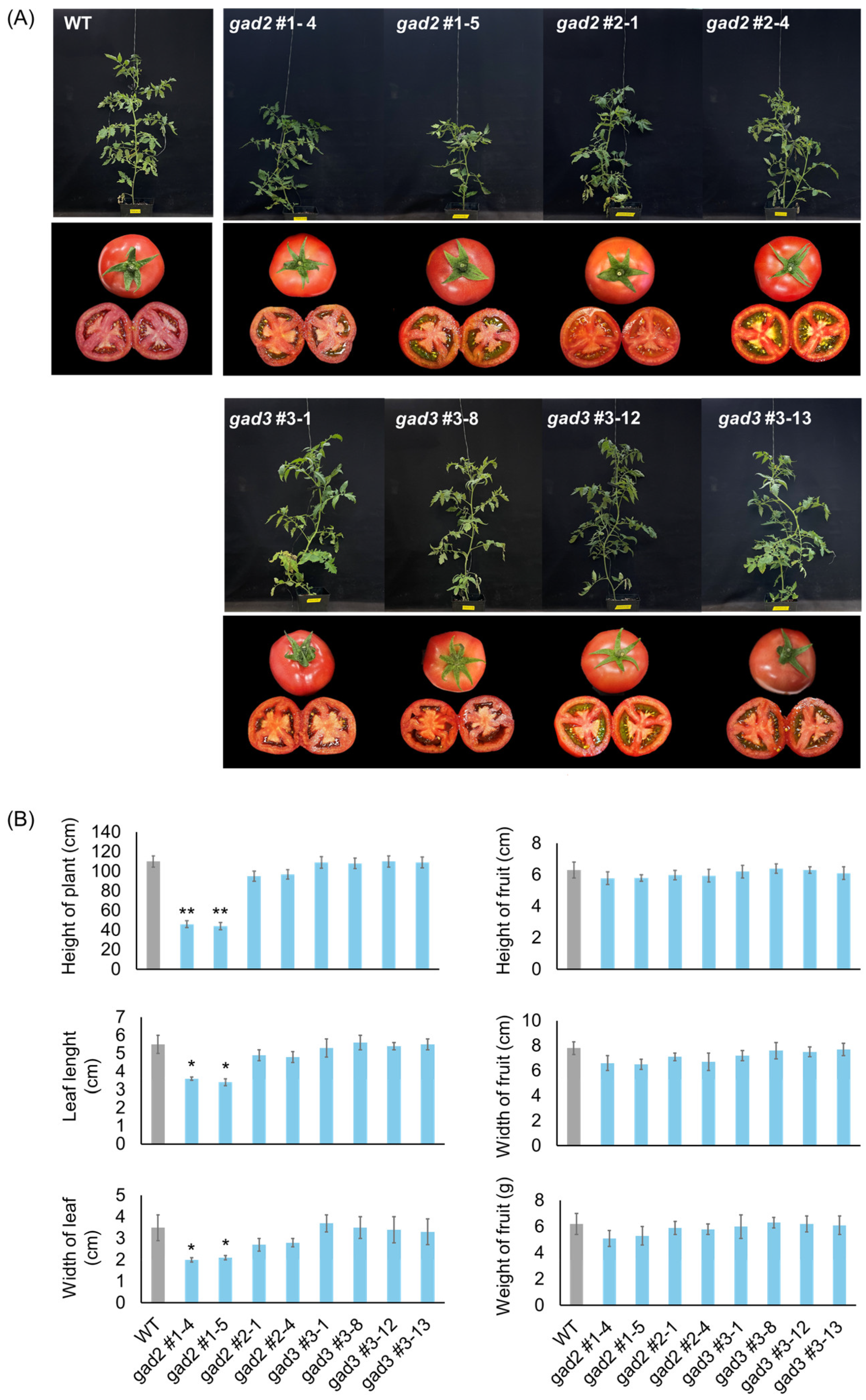
3.2. Phenotypic Characterization of Homozygous SlGAD2 and SlGAD3 Edited Lines
3.3. GABA and Free Amino Acid Profiling in Edited Tomato Fruit
3.4. Transcriptomic Profiling of SlGAD2 and SlGAD3 Mutants Highlights GABA-Associated Pathways
3.5. qRT-PCR Validation of Differentially Expressed Genes Related to GABA Metabolism and Stress Signaling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Le Magueresse, C.; Monyer, H. GABAergic interneurons shape the functional maturation of the cortex. Neuron 2013, 77, 388–405. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Shirai, T.; Ochiai, H.; Kasao, M.; Hayakawa, K.; Kimura, M.; Sansawa, H. Blood-pressure-lowering effect of a novel fermented milk containing γ-aminobutyric acid (GABA) in mild hypertensives. Eur. J. Clin. Nutr. 2003, 57, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Abdou, A.M.; Higashiguchi, S.; Horie, K.; Kim, M.; Hatta, H.; Yokogoshi, H. Relaxation and immunity enhancement effects of γ-aminobutyric acid (GABA) administration in humans. Biofactors 2006, 26, 201–208. [Google Scholar] [CrossRef]
- Wu, Q.Y.; Ma, S.Z.; Zhang, W.W.; Yao, K.B.; Chen, L.; Zhao, F.; Zhuang, Y.Q. Accumulating pathways of γ-aminobutyric acid during anaerobic and aerobic sequential incubations in fresh tea leaves. Food Chem. 2018, 240, 1081–1086. [Google Scholar] [CrossRef]
- Shimada, M.; Hasegawa, T.; Nishimura, C.; Kan, H.; Kanno, T.; Nakamura, T.; Matsubayashi, T. Anti-hypertensive effect of γ-aminobutyric acid (GABA)-rich Chlorella on high-normal blood pressure and borderline hypertension in placebo-controlled double-blind study. Clin. Exp. Hypertens. 2009, 31, 342–354. [Google Scholar] [CrossRef]
- Satya Narayan, V.; Nair, P.M. Metabolism, enzymology and possible roles of gamma-aminobutyric acid. Biochemistry 1989, 8, 21–25. [Google Scholar]
- Bouche, N.; Fromm, H. GABA in plants: Just a metabolite? Trends Plant Sci. 2004, 9, 110–115. [Google Scholar] [CrossRef]
- Takayama, M.; Ezura, H. How and why does tomato accumulate a large amount of GABA in the fruit? Front. Plant Sci. 2015, 6, 612. [Google Scholar] [CrossRef]
- Fait, A.; Fromm, H.; Walter, D.; Galili, G.; Fernie, A.R. Highway or byway: The metabolic role of the GABA shunt in plants. Trends Plant Sci. 2008, 13, 14–19. [Google Scholar] [CrossRef]
- Shelp, B.J.; Bown, A.W.; McLean, M.D. Metabolism and functions of gamma-aminobutyric acid. Trends Plant Sci. 1999, 4, 446–452. [Google Scholar] [CrossRef]
- Baum, G.; Chen, Y.; Arazi, T.; Takatsuji, H.; Fromm, H. A plant glutamate decarboxylase containing a calmodulin binding domain: Cloning, sequence, and functional analysis. J. Biol. Chem. 1993, 268, 19610–19617. [Google Scholar] [CrossRef] [PubMed]
- Shimajiri, Y.; Oonishi, T.; Ozaki, K.; Kainou, K.; Akama, K. Genetic manipulation of the γ-aminobutyric acid (GABA) shunt in rice: Overexpression of truncated glutamate decarboxylase (GAD2) and knockdown of γ-aminobutyric acid transaminase (GABA-T) lead to sustained and high levels of GABA accumulation in rice kernels. Plant Biotechnol. J. 2013, 11, 594–604. [Google Scholar] [PubMed]
- Akihiro, T.; Koike, S.; Tani, R.; Tominaga, T.; Watanabe, S.; Iijima, Y.; Aoki, K.; Shibata, D.; Ezura, H. Biochemical mechanism on GABA accumulation during fruit development in tomato. Plant Cell Physiol. 2008, 49, 1378–1389. [Google Scholar] [CrossRef]
- Nonaka, S.; Arai, C.; Takayama, M.; Matsukura, C.; Ezura, H. Efficient increase of γ-aminobutyric acid (GABA) content in tomato fruits by targeted mutagenesis. Sci. Rep. 2017, 7, 7057. [Google Scholar] [CrossRef]
- Park, J.; Bae, S.; Kim, J.S. Cas-Designer: A web-based tool for choice of CRISPR-Cas9 target sites. Bioinformatics 2015, 31, 4014–4016. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, D.H.; Kim, M.-S.; Jung, Y.J.; Kang, K.K. Physicochemical properties and antioxidant activity of CRISPR/Cas9-edited tomato SGR1 knockout (KO) line. Int. J. Mol. Sci. 2024, 25, 5111. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, J.H.; Jang, Y.H.; Yu, J.; Bae, S.; Kim, M.-S.; Cho, Y.-G.; Jung, Y.J.; Kang, K.K. Transcriptome and metabolite profiling of tomato SGR-knockout null lines using the CRISPR/Cas9 system. Int. J. Mol. Sci. 2023, 24, 109. [Google Scholar] [CrossRef]
- Zik, M.; Arazi, T.; Snedden, W.A.; Fromm, H. Two isoforms of glutamate decarboxylase in Arabidopsis are regulated by calcium/calmodulin and differ in organ distribution. Plant Mol. Biol. 1998, 37, 967–975. [Google Scholar] [CrossRef]
- Akama, K.; Akihiro, T.; Kitagawa, M.; Takaiwa, F. Rice (Oryza sativa) contains a novel isoform of glutamate decarboxylase that lacks an authentic calmodulin-binding domain at the C-terminus. Biochim. Biophys. Acta Gene Struct. Expr. 2001, 1522, 143–150. [Google Scholar] [CrossRef]
- Mishra, V.; Gahlowt, P.; Singh, S.; Dubey, N.K.; Singh, S.P.; Tripathi, D.K.; Singh, V.P. GABA: A key player of abiotic stress regulation. Plant Signal. Behav. 2023, 18, 2163343. [Google Scholar] [CrossRef]
- Kinnersley, A.M.; Turano, F.J. Gamma aminobutyric acid (GABA) and plant responses to stress. Crit. Rev. Plant Sci. 2000, 19, 479–509. [Google Scholar] [CrossRef]
- Ramesh, S.A.; Tyerman, S.D.; Gilliham, M.; Xu, B. γ-Aminobutyric acid (GABA) signalling in plants. Cell. Mol. Life Sci. 2017, 74, 1577–1603. [Google Scholar] [CrossRef]
- Takayama, M.; Matsukura, C.; Ariizumi, T.; Ezura, H. Activating glutamate decarboxylase activity by removing the autoinhibitory domain leads to hyper γ-aminobutyric acid (GABA) accumulation in tomato fruit. Plant Cell Rep. 2017, 36, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Renault, H.; Roussel, V.; El Amrani, A.; Arzel, M.; Renault, D.; Bouchereau, A.; Deleu, C. The Arabidopsispop2-1 mutant reveals the involvement of GABA transaminase in salt stress tolerance. BMC Plant Biol. 2010, 10, 20. [Google Scholar] [CrossRef] [PubMed]
- Bouché, N.; Fait, A.; Bouchez, D.; Møller, S.G.; Fromm, H. Mitochondrial succinic-semialdehyde dehydrogenase of the γ-aminobutyrate shunt is required to restrict levels of reactive oxygen intermediates in plants. Proc. Natl. Acad. Sci. USA 2003, 100, 6843–6848. [Google Scholar] [CrossRef]
- Wu, Q.; Su, N.; Huang, X.; Cui, J.; Shabala, L.; Zhou, M.; Chen, Z.-H.; Shabala, S. Hypoxia-induced increase in GABA content is essential for restoration of membrane potential and preventing ROS-induced disturbance to ion homeostasis. Plant Commun. 2021, 2, 100178. [Google Scholar] [CrossRef]
- Mekonnen, D.W.; Flügge, U.I.; Ludewig, F. Gamma-aminobutyric acid depletion affects stomata closure and drought tolerance of Arabidopsis thaliana. Plant Sci. 2016, 245, 25–34. [Google Scholar] [CrossRef]
- Sita, K.; Kumar, V. Role of gamma-aminobutyric acid (GABA) against abiotic stress tolerance in legumes: A review. Plant Physiol. Rep. 2020, 25, 654–663. [Google Scholar] [CrossRef]
- Hayat, F.; Khan, U.; Li, J.; Ahmed, N.; Khanum, F.; Iqbal, S.; Shahid, M.A. γ-Aminobutyric acid (GABA): A key player in alleviating abiotic stress resistance in horticultural crops: Current insights and future directions. Horticulturae 2023, 9, 647. [Google Scholar] [CrossRef]
- Wang, X.; Li, X.; Dong, S. Biochemical characterization and metabolic reprogramming of amino acids in soybean roots under drought stress. Physiol Plant 2024, 176, e14319. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Gene ID | Gene Description | WT vs. gad2 #1–5 | WT vs. gad3 #3–8 | ||
|---|---|---|---|---|---|---|
| log2FC | padj | log2FC | padj | |||
| GABA Shunt | Solyc01g005000.3.1 | Glutamate decarboxylase (GAD) | 1.31 | 9 × 10−4 | 1.73 | 6 × 10−6 |
| Solyc01g080280.3.1 | Chloroplast glutamine synthetase | 2.92 | 2 × 10−40 | 1.35 | 1 × 10−8 | |
| Solyc03g078150.3.1 | Amino acid transporter family protein | 2.52 | 2 × 10−27 | 1.22 | 2 × 10−6 | |
| Solyc03g113980.3.1 | Calmodulin binding protein-like | −2.69 | 5 × 10−5 | −1.93 | 6 × 10−3 | |
| Solyc03g120090.1.1 | Pyridoxal 5′-phosphate synthase pdxS subunit | −1.94 | 6 × 10−103 | −1.47 | 3 × 10−62 | |
| Cell membrane transport and Ca2+-K+ ion transportation | Solyc10g006660.3.1 | Calcium-binding protein PBP1 | −1.16 | 3 × 10−8 | −2.3 | 4 × 10−27 |
| Solyc09g007860.4.1 | Calcium-dependent lipid-binding (CaLB domain) protein | −1.23 | 1 × 10−16 | −1.23 | 3 × 10−16 | |
| Solyc02g091500.1.1 | Calcium-dependent protein kinase (CDPK) | −1.58 | 7 × 10−4 | −1.43 | 4 × 10−3 | |
| Solyc05g051220.3.1 | Potassium outward rectifying channel GORK | −1.02 | 3 × 10−9 | −1.45 | 9 × 10−17 | |
| Solyc08g016500.3.1 | Potassium inward channel KAT1 | 1.06 | 2 × 10−3 | 1.13 | 1 × 10−3 | |
| Solyc10g006800.4.1 | Cyclic nucleotide-gated ion channel 4 | −1.93 | 2 × 10−4 | −2.42 | 4 × 10−6 | |
| Solyc02g094000.1.1 | EF-hand domain protein (Ca2+ binding) | −2.19 | 5 × 10−5 | −3.37 | 3 × 10−9 | |
| Solyc03g026280.3.1 | C-repeat binding factor 1 (possibly involved in cold/ion stress) | −3.03 | 2 × 10−6 | −5.03 | 5 × 10−12 | |
| Solyc09g005260.4.1 | Vacuolar cation/proton exchanger (ion homeostasis under stress) | −2.14 | 5 × 10−32 | −2.15 | 5 × 10−32 | |
| Stress Response | Solyc01g098270.1.1 | Chaperone protein DnaJ | −6.16 | 3 × 10−21 | −7.35 | 1 × 10−22 |
| Solyc09g092260.4.1 | Chaperone protein DnaJ | 1.74 | 8 × 10−5 | 2.33 | 5 × 10−8 | |
| Solyc09g005120.3.1 | DnaJ protein ERDJ3A | −2.25 | 7 × 10−138 | −1.32 | 8 × 10−54 | |
| Solyc02g077670.3.1 | DnaJ-like protein | −1.85 | 1 × 10−232 | −1.26 | 5 × 10−117 | |
| Solyc04g007470.3.1 | Drought responsive Zinc finger protein | 1.28 | 2 × 10−4 | 1.34 | 1 × 10−4 | |
| Solyc02g077610.3.1 | NAC domain protein | −2.07 | 3 × 10−44 | −1.06 | 3 × 10−13 | |
| Solyc02g093420.4.1 | NAC domain-containing protein 10 | −1.37 | 1 × 10−6 | −1.13 | 1 × 10−4 | |
| Solyc09g015770.3.1 | WRKY transcription factor 81 | −1.12 | 2 × 10−8 | −1.91 | 8 × 10−21 | |
| Solyc04g077980.1.1 | C2H2-type zinc finger protein | −1.58 | 4 × 10−10 | −2.98 | 3 × 10−29 | |
| Solyc10g006130.1.1 | EAR motif-containing protein SlERF36 | −1.33 | 1 × 10−11 | −1.94 | 2 × 10−22 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-Y.; Jung, Y.-J.; Kim, D.H.; Kang, K.-K. Transcriptomic Insights into GABA Accumulation in Tomato via CRISPR/Cas9-Based Editing of SlGAD2 and SlGAD3. Genes 2025, 16, 744. https://doi.org/10.3390/genes16070744
Kim J-Y, Jung Y-J, Kim DH, Kang K-K. Transcriptomic Insights into GABA Accumulation in Tomato via CRISPR/Cas9-Based Editing of SlGAD2 and SlGAD3. Genes. 2025; 16(7):744. https://doi.org/10.3390/genes16070744
Chicago/Turabian StyleKim, Jin-Young, Yu-Jin Jung, Dong Hyun Kim, and Kwon-Kyoo Kang. 2025. "Transcriptomic Insights into GABA Accumulation in Tomato via CRISPR/Cas9-Based Editing of SlGAD2 and SlGAD3" Genes 16, no. 7: 744. https://doi.org/10.3390/genes16070744
APA StyleKim, J.-Y., Jung, Y.-J., Kim, D. H., & Kang, K.-K. (2025). Transcriptomic Insights into GABA Accumulation in Tomato via CRISPR/Cas9-Based Editing of SlGAD2 and SlGAD3. Genes, 16(7), 744. https://doi.org/10.3390/genes16070744