
Satellite DNA Mapping in Suliformes (Aves): Insights into the Evolution of the Multiple Sex Chromosome System in Sula spp.

 , , , ,
, , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Chromosomal Obtainment
2.2. DNA Extraction, Genome Sequencing, and Bioinformatic Analysis
2.3. SatDNA Probes and Fluorescence In Situ Hybridization (FISH)
2.4. Image Processing
3. Results
3.1. Abundance and Characteristics of the satDNAs
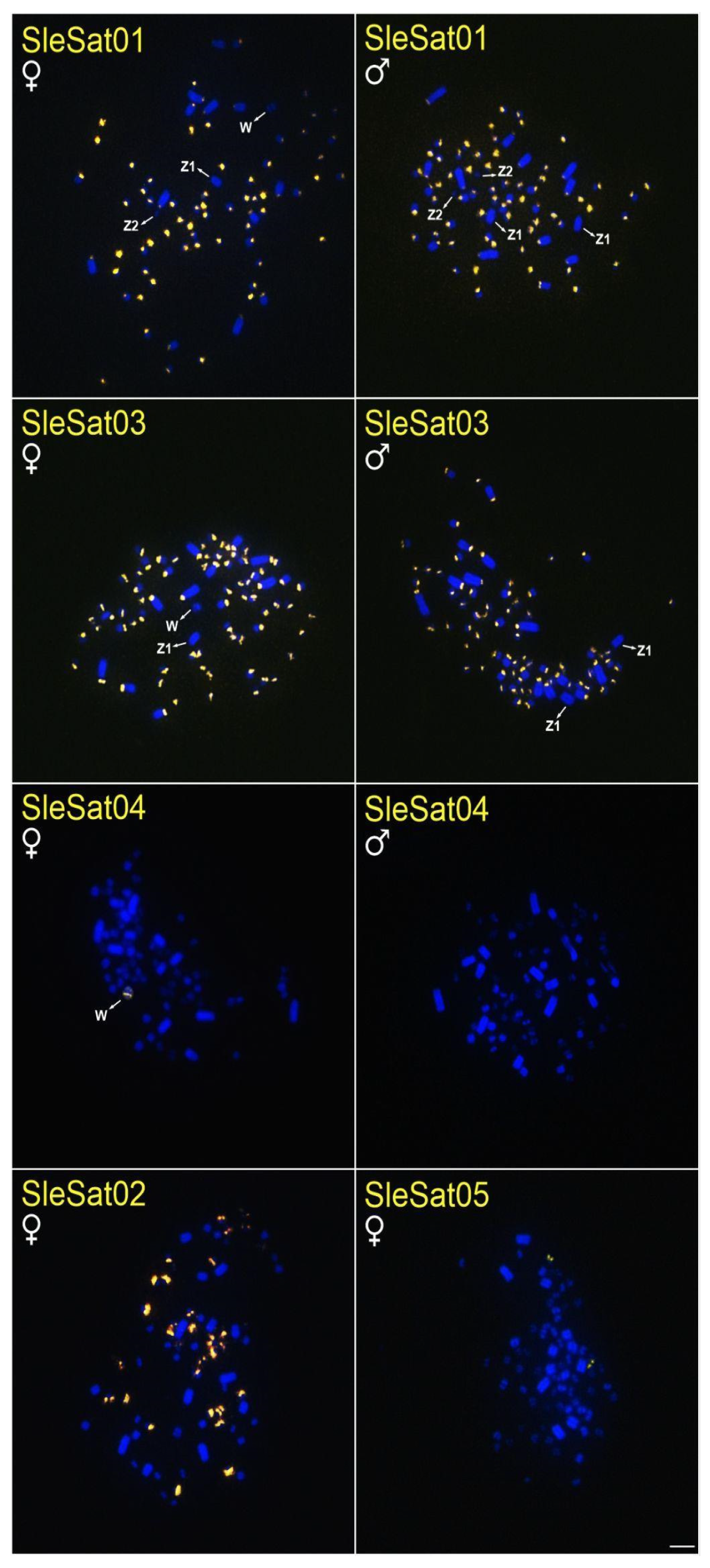
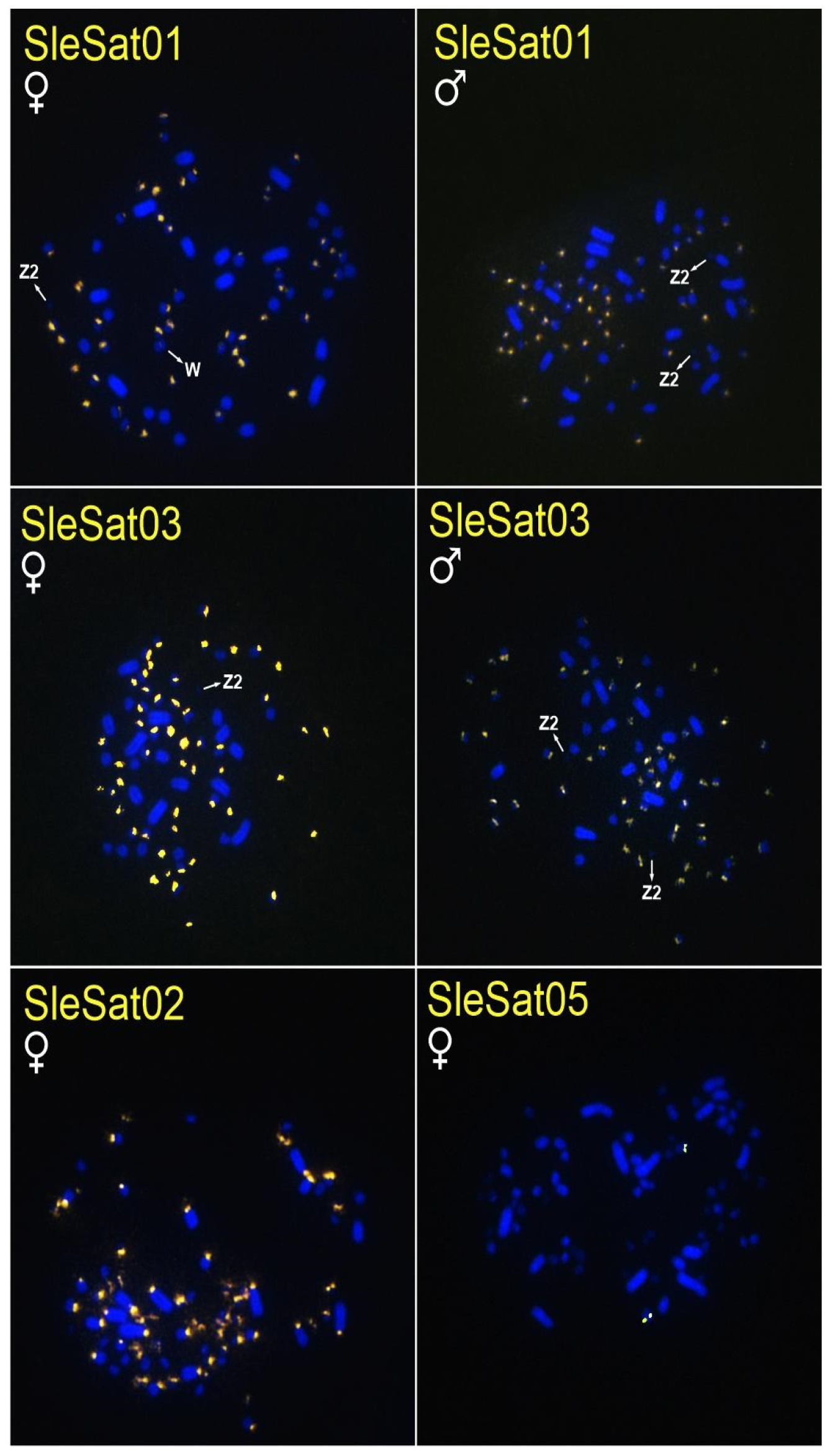
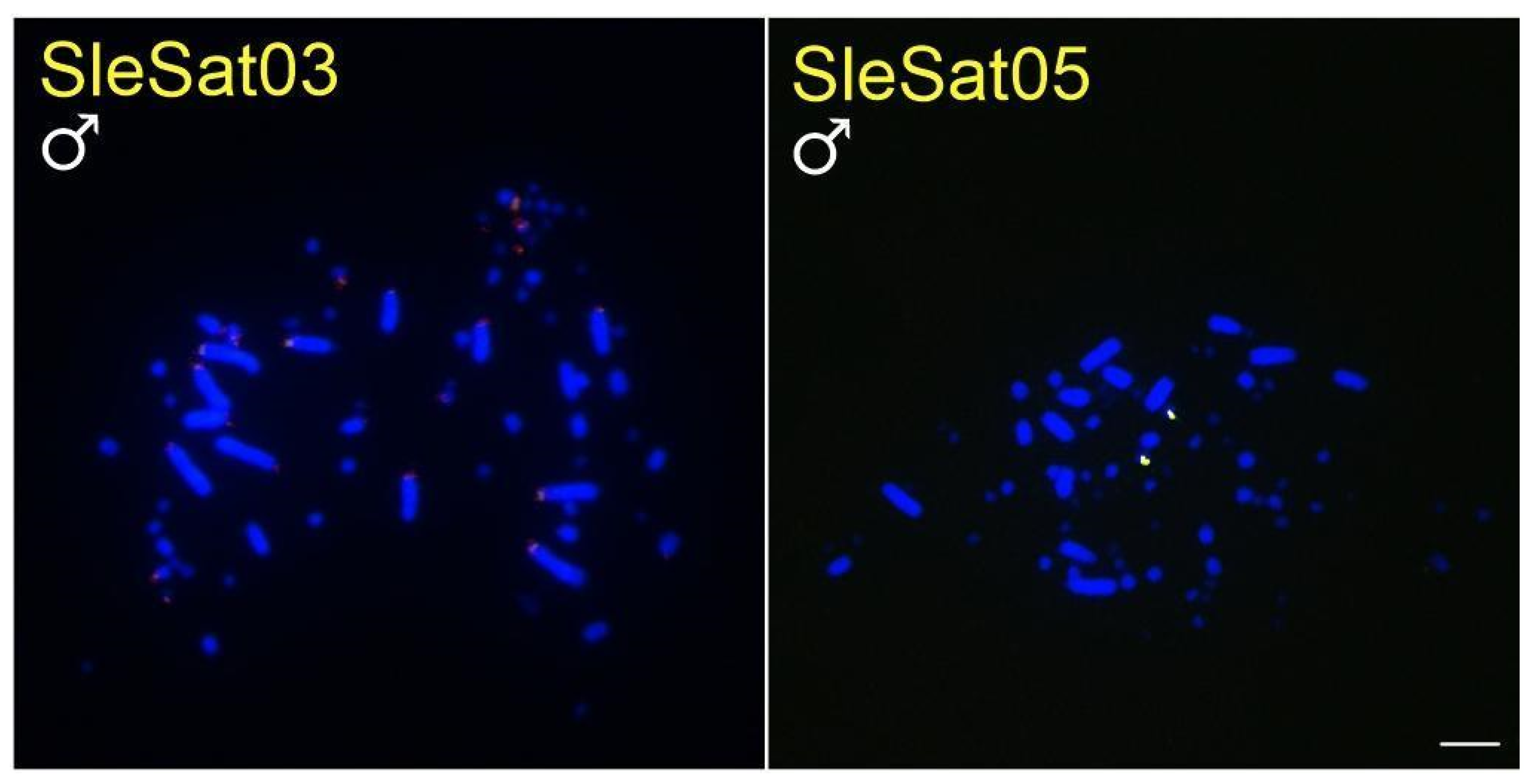
3.2. Chromosomal Location of S. leucogaster satDNAs (SleSatDNAs)
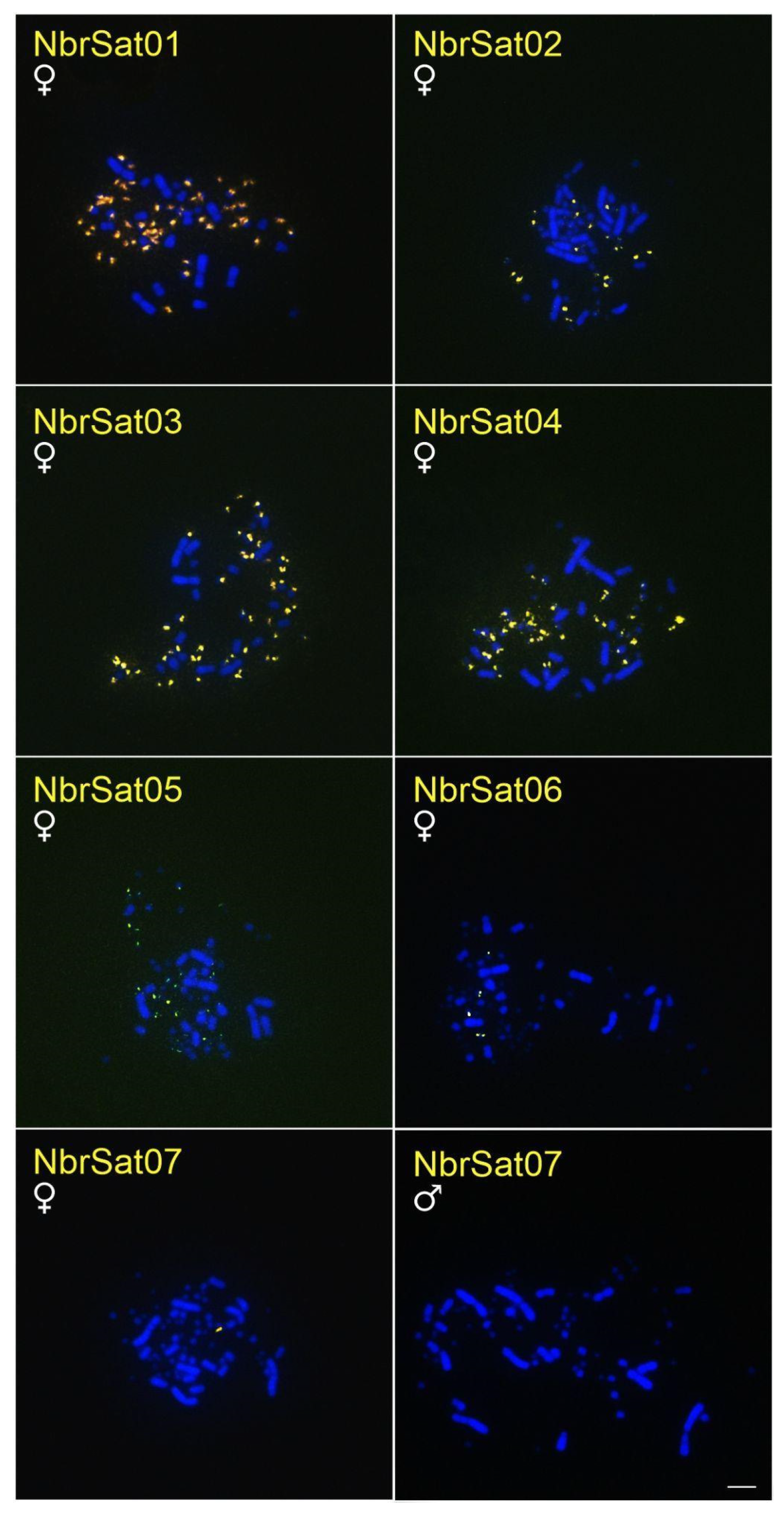
3.3. Chromosomal Location of N. brasilianum satDNAs (NbrSatDNAs)
4. Discussion
4.1. Overview of Satellite DNA in S. leucogaster and N. brasilianum
4.2. Comparative Chromosomal Mapping of Satellite DNAs in Suliformes
4.3. Satellite DNA Mapping in Suliformes Sheds Light on Sex Chromosome Evolution
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Charlesworth, B.; Sniegowski, P.; Stephan, W. The evolutionary dynamics of repetitive DNA in eukaryotes. Nature 1994, 371, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T. Genomes, genes and junk: The large-scale organization of plant chromosomes. Trends Plant Sci. 1998, 3, 195–199. [Google Scholar] [CrossRef]
- Richard, G.-F.; Kerrest, A.; Dujon, B. Comparative Genomics and Molecular Dynamics of DNA Repeats in Eukaryotes. Microbiol. Mol. Biol. Rev. 2008, 72, 686–727. [Google Scholar] [CrossRef] [PubMed]
- Plohl, M.; Meštrović, N.; Mravinac, B. Satellite DNA Evolution. In Repetitive DNA; Garrido-Ramos, M.A., Ed.; S. Karger AG: Basel, Switzerland, 2012; Volume 7, pp. 126–152. ISBN 978-3-318-02149-3. [Google Scholar]
- Garrido-Ramos, M. Satellite DNA: An Evolving Topic. Genes 2017, 8, 230. [Google Scholar] [CrossRef]
- Ruiz-Ruano, F.J.; López-León, M.D.; Cabrero, J.; Camacho, J.P.M. High-throughput analysis of the satellitome illuminates satellite DNA evolution. Sci. Rep. 2016, 6, 28333. [Google Scholar] [CrossRef]
- Peona, V.; Palacios-Gimenez, O.M.; Blommaert, J.; Liu, J.; Haryoko, T.; Jønsson, K.A.; Irestedt, M.; Zhou, Q.; Jern, P.; Suh, A. The avian W chromosome is a refugium for endogenous retroviruses with likely effects on female-biased mutational load and genetic incompatibilities. Phil. Trans. R. Soc. B 2021, 376, 20200186. [Google Scholar] [CrossRef]
- Wei, L.; Liu, B.; Zhang, C.; Yu, Y.; Yang, X.; Dou, Q.; Dong, Q. Identification and characterization of satellite DNAs in Poa L. Mol. Cytogenet. 2020, 13, 47. [Google Scholar] [CrossRef]
- De Lima, L.G.; Ruiz-Ruano, F.J. In-Depth Satellitome Analyses of 37 Drosophila Species Illuminate Repetitive DNA Evolution in the Drosophila Genus. Genome Biol. Evol. 2022, 14, evac064. [Google Scholar] [CrossRef]
- Fry, K.; Salser, W. Nucleotide sequences of HS-α satellite DNA from kangaroo rat dipodomys ordii and characterization of similar sequences in other rodents. Cell 1977, 12, 1069–1084. [Google Scholar] [CrossRef]
- Yurchenko, A.; Pšenička, T.; Mora, P.; Ortega, J.A.M.; Baca, A.S.; Rovatsos, M. Cytogenetic Analysis of Satellitome of Madagascar Leaf-Tailed Geckos. Genes 2024, 15, 429. [Google Scholar] [CrossRef]
- Despot-Slade, E.; Širca, S.; Mravinac, B.; Castagnone-Sereno, P.; Plohl, M.; Meštrović, N. Satellitome analyses in nematodes illuminate complex species history and show conserved features in satellite DNAs. BMC Biol. 2022, 20, 259. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, M.J.; Fogarin Destro, R.; Gazoni, T.; Narimatsu, H.; Pereira Dos Santos, P.S.; Haddad, C.F.B.; Parise-Maltempi, P.P. Great Abundance of Satellite DNA in Proceratophrys (Anura, Odontophrynidae) Revealed by Genome Sequencing. Cytogenet. Genome Res. 2020, 160, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Cabral-de-Mello, D.C.; Zrzavá, M.; Kubíčková, S.; Rendón, P.; Marec, F. The Role of Satellite DNAs in Genome Architecture and Sex Chromosome Evolution in Crambidae Moths. Front. Genet. 2021, 12, 661417. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.A.; Milani, D.; Ferretti, A.B.S.M.; Bardella, V.B.; Cabral-de-Mello, D.C.; Lopes, D.M. The extensive amplification of heterochromatin in Melipona bees revealed by high throughput genomic and chromosomal analysis. Chromosoma 2021, 130, 251–262. [Google Scholar] [CrossRef]
- Voleníková, A.; Lukšíková, K.; Mora, P.; Pavlica, T.; Altmanová, M.; Štundlová, J.; Pelikánová, Š.; Simanovsky, S.A.; Jankásek, M.; Reichard, M.; et al. Fast satellite DNA evolution in Nothobranchius annual killifishes. Chromosome Res. 2023, 31, 33. [Google Scholar] [CrossRef]
- Crepaldi, C.; Martí, E.; Gonçalves, É.M.; Martí, D.A.; Parise-Maltempi, P.P. Genomic Differences Between the Sexes in a Fish Species Seen Through Satellite DNAs. Front. Genet. 2021, 12, 728670. [Google Scholar] [CrossRef]
- Sena, R.S.; Heringer, P.; Valeri, M.P.; Pereira, V.S.; Kuhn, G.C.S.; Svartman, M. Identification and characterization of satellite DNAs in two-toed sloths of the genus Choloepus (Megalonychidae, Xenarthra). Sci. Rep. 2020, 10, 19202. [Google Scholar] [CrossRef]
- Vozdova, M.; Kubickova, S.; Martínková, N.; Galindo, D.J.; Bernegossi, A.M.; Cernohorska, H.; Kadlcikova, D.; Musilová, P.; Duarte, J.M.; Rubes, J. Satellite DNA in Neotropical Deer Species. Genes 2021, 12, 123. [Google Scholar] [CrossRef]
- Gutiérrez, J.; Aleix-Mata, G.; Montiel, E.E.; Cabral-de-Mello, D.C.; Marchal, J.A.; Sánchez, A. Satellitome Analysis on Talpa aquitania Genome and Inferences about the satDNAs Evolution on Some Talpidae. Genes 2022, 14, 117. [Google Scholar] [CrossRef]
- Melters, D.P.; Bradnam, K.R.; Young, H.A.; Telis, N.; May, M.R.; Ruby, J.G.; Sebra, R.; Peluso, P.; Eid, J.; Rank, D.; et al. Comparative analysis of tandem repeats from hundreds of species reveals unique insights into centromere evolution. Genome Biol. 2013, 14, R10. [Google Scholar] [CrossRef]
- Ohno, S. So much “junk” DNA in our genome. Brookhaven Symp. Biol. 1972, 23, 366–370. [Google Scholar] [PubMed]
- Dernburg, A.F.; Sedat, J.W.; Hawley, R.S. Direct Evidence of a Role for Heterochromatin in Meiotic Chromosome Segregation. Cell 1996, 86, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, G.C.S. Satellite DNA transcripts have diverse biological roles in Drosophila. Heredity 2015, 115, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Shatskikh, A.S.; Kotov, A.A.; Adashev, V.E.; Bazylev, S.S.; Olenina, L.V. Functional Significance of Satellite DNAs: Insights From Drosophila. Front. Cell. Dev. Biol. 2020, 8, 312. [Google Scholar] [CrossRef]
- Cattani, M.V.; Presgraves, D.C. Incompatibility between X chromosome factor and pericentric heterochromatic region causes lethality in hybrids between Drosophila melanogaster and its sibling species. Genetics 2012, 191, 549–559. [Google Scholar] [CrossRef]
- Rošić, S.; Köhler, F.; Erhardt, S. Repetitive centromeric satellite RNA is essential for kinetochore formation and cell division. J. Cell. Biol. 2014, 207, 335–349. [Google Scholar] [CrossRef]
- Bayes, J.J.; Malik, H.S. Altered Heterochromatin Binding by a Hybrid Sterility Protein in Drosophila Sibling Species. Science 2009, 326, 1538–1541. [Google Scholar] [CrossRef]
- Jagannathan, M.; Yamashita, Y.M. Defective Satellite DNA Clustering into Chromocenters Underlies Hybrid Incompatibility in Drosophila. Mol. Biol. Evol. 2021, 38, 4977–4986. [Google Scholar] [CrossRef]
- Ferretti, A.B.S.M.; Milani, D.; Palacios-Gimenez, O.M.; Ruiz-Ruano, F.J.; Cabral-de-Mello, D.C. High dynamism for neo-sex chromosomes: Satellite DNAs reveal complex evolution in a grasshopper. Heredity 2020, 125, 124–137. [Google Scholar] [CrossRef]
- Gatto, K.P.; Busin, C.S.; Lourenço, L.B. Unraveling the Sex Chromosome Heteromorphism of the Paradoxical Frog Pseudis tocantins. PLoS ONE 2016, 11, e0156176. [Google Scholar] [CrossRef]
- Smeds, L.; Warmuth, V.; Bolivar, P.; Uebbing, S.; Burri, R.; Suh, A.; Nater, A.; Bureš, S.; Garamszegi, L.Z.; Hogner, S.; et al. Evolutionary analysis of the female-specific avian W chromosome. Nat. Commun. 2015, 6, 7330. [Google Scholar] [CrossRef] [PubMed]
- Peona, V.; Blom, M.P.K.; Xu, L.; Burri, R.; Sullivan, S.; Bunikis, I.; Liachko, I.; Haryoko, T.; Jønsson, K.A.; Zhou, Q.; et al. Identifying the causes and consequences of assembly gaps using a multiplatform genome assembly of a bird-of-paradise. Mol. Ecol. Resour. 2021, 21, 263–286. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, A.M.; Souza, G.M.; Toma, G.A.; Dos Santos, N.; Dos Santos, R.Z.; Goes, C.A.G.; Deon, G.A.; Setti, P.G.; Porto-Foresti, F.; Utsunomia, R.; et al. Satellite DNAs, heterochromatin, and sex chromosomes of the wattled jacana (Charadriiformes; Jacanidae): A species with highly rearranged karyotype. Genome 2024, 67, 109–118. [Google Scholar] [CrossRef]
- Kretschmer, R.; Toma, G.A.; Deon, G.A.; Dos Santos, N.; Dos Santos, R.Z.; Utsunomia, R.; Porto-Foresti, F.; Gunski, R.J.; Garnero, A.D.V.; Liehr, T.; et al. Satellitome Analysis in the Southern Lapwing (Vanellus chilensis) Genome: Implications for SatDNA Evolution in Charadriiform Birds. Genes 2024, 15, 258. [Google Scholar] [CrossRef] [PubMed]
- Setti, P.G.; Deon, G.A.; Zeni Dos Santos, R.; Goes, C.A.G.; Garnero, A.D.V.; Gunski, R.J.; De Oliveira, E.H.C.; Porto-Foresti, F.; De Freitas, T.R.O.; Silva, F.A.O.; et al. Evolution of bird sex chromosomes: A cytogenomic approach in Palaeognathae species. BMC Ecol. Evo. 2024, 24, 51. [Google Scholar] [CrossRef]
- Souza, G.M.; Kretschmer, R.; Toma, G.A.; De Oliveira, A.M.; Deon, G.A.; Setti, P.G.; Zeni Dos Santos, R.; Goes, C.A.G.; Del Valle Garnero, A.; Gunski, R.J.; et al. Satellitome analysis on the pale-breasted thrush Turdus leucomelas (Passeriformes; Turdidae) uncovers the putative co-evolution of sex chromosomes and satellite DNAs. Sci. Rep. 2024, 14, 20656. [Google Scholar] [CrossRef]
- Souza, G.M.; Vidal, J.A.D.; Utsunomia, R.; Deon, G.A.; De Oliveira, E.H.C.; Franca, R.T.; Porto-Foresti, F.; Liehr, T.; De Souza, F.H.S.; Kretschmer, R.; et al. Cytogenomic analysis in Seriemas (Cariamidae): Insights into an Atypical Avian Karyotype. J. Hered. 2025, esaf012. [Google Scholar] [CrossRef]
- Theodorescu, R.C. The Karyotypic Evolution in Two Pelecaniformes Species (Aves). Caryologia 1975, 28, 459–466. [Google Scholar] [CrossRef]
- Patnaik, A.K.; Samanta, M.; Prasad, R. Chromosome complement and banding patterns in a pelecaniform bird, Phalacrocorax niger. J. Hered. 1981, 72, 447–449. [Google Scholar] [CrossRef]
- Belterman, R.H.R.; De Boer, L.E.M. A karyological study of 55 species of birds, including karyotypes of 39 species new to cytology. Genetica 1984, 65, 39–82. [Google Scholar] [CrossRef]
- Bhunya, S.P.; Mohanty, M.K. Localization of constitutive heterochromatin (C-band) and nucleolus organizers (NORs) in the somatic chromosomes of a pelecaniform bird Phalacrocorax niger (Viellot). Chrom. Inform. Serv. 1985, 39, 17–19. [Google Scholar]
- Ebied, A.M.; Hassan, H.A.; Abu Almaaty, A.H.; Yaseen, A.E. Karyotypic Characterization of Ten Species of Birds. Cytologia 2005, 70, 181–194. [Google Scholar] [CrossRef]
- Ledesma, M.A.; Cardozo, D.E.; Montalti, D.; Leotta, G.A.; Gunski, R. Estudios citogenéticos en tres especies de Aves Antárticas. Rev. De Cienc. Y Tecnol. 2005, 7. [Google Scholar]
- Griffin, D.K.; Robertson, L.B.W.; Tempest, H.G.; Skinner, B.M. The evolution of the avian genome as revealed by comparative molecular cytogenetics. Cytogenet. Genome Res. 2007, 117, 64–77. [Google Scholar] [CrossRef]
- Kretschmer, R.; De Souza, M.S.; Furo, I.D.O.; Romanov, M.N.; Gunski, R.J.; Garnero, A.D.V.; De Freitas, T.R.O.; De Oliveira, E.H.C.; O’Connor, R.E.; Griffin, D.K. Interspecies Chromosome Mapping in Caprimulgiformes, Piciformes, Suliformes, and Trogoniformes (Aves): Cytogenomic Insight into Microchromosome Organization and Karyotype Evolution in Birds. Cells 2021, 10, 826. [Google Scholar] [CrossRef] [PubMed]
- Pozzobon, L.C.; Toma, G.A.; Cioffi, M.D.B.; De Oliveira, E.H.C.; Kretschmer, R.; De Freitas, T.R.O. Karyotype evolution of suliformes and description of a ♂Z1Z1Z2Z2/♀Z1Z2W multiple sex chromosome system in boobies (Sula spp.). Genome 2025, 68, 1–11. [Google Scholar] [CrossRef]
- Sasaki, M.; Hitotsumachi, S.; Makino, S.; Terao, T. A Comparative Study of the Chromosomes in the Chum Salmon, the Kokanee Salmon and their Hybrids. Caryologia 1968, 21, 389–394. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Novák, P.; Neumann, P.; Macas, J. Global analysis of repetitive DNA from unassembled sequence reads using RepeatExplorer2. Nat. Protoc. 2020, 15, 3745–3776. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Fast Identification and Removal of Sequence Contamination from Genomic and Metagenomic Datasets. PLoS ONE 2011, 6, e17288. [Google Scholar] [CrossRef]
- Smit, A.F.A.; Hubley, R.; Green, P. RepeatMasker Open-4.1.5. 2013–2015. Available online: http://www.repeatmasker.org. (accessed on 30 October 2023).
- Kretschmer, R.; dos Santos, M.S.; Furo, I.D.O.; Cioffi, M.D.B. FISH—In Birds. In Cytogenetics and Molecular Cytogenetics; Liehr, T., Ed.; CRC Press: Boca Raton, FL, USA, 2022; Volume 1, pp. 263–280. ISBN 978-1-003-22365-8. [Google Scholar]
- Peona, V.; Kutschera, V.E.; Blom, M.P.K.; Irestedt, M.; Suh, A. Satellite DNA evolution in Corvoidea inferred from short and long reads. Mol. Ecol. 2023, 32, 1288–1305. [Google Scholar] [CrossRef] [PubMed]
- Boštjančić, L.L.; Bonassin, L.; Anušić, L.; Lovrenčić, L.; Besendorfer, V.; Maguire, I.; Grandjean, F.; Austin, C.M.; Greve, C.; Hamadou, A.B.; et al. The Pontastacus leptodactylus (Astacidae) Repeatome Provides Insight Into Genome Evolution and Reveals Remarkable Diversity of Satellite DNA. Front. Genet. 2021, 11, 611745. [Google Scholar] [CrossRef] [PubMed]
- Lorite, P.; Carrillo, J.A.; Aguilar, J.A.; Palomeque, T. Isolation and characterization of two families of satellite DNA with repetitive units of 135 bp and 2.5 kb in the ant Monomorium subopacum (Hymenoptera, Formicidae). Cytogenet. Genome Res. 2004, 105, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Crepaldi, C.; Parise-Maltempi, P.P. Heteromorphic Sex Chromosomes and Their DNA Content in Fish: An Insight through Satellite DNA Accumulation in Megaleporinus elongatus. Cytogenet. Genome Res. 2020, 160, 38–46. [Google Scholar] [CrossRef]
- Rojo, V.; Martínez-Lage, A.; Giovannotti, M.; González-Tizón, A.M.; Cerioni, P.N.; Barucchi, V.C.; Galán, P.; Olmo, E.; Naveira, H. Evolutionary dynamics of two satellite DNA families in rock lizards of the genus Iberolacerta (Squamata, Lacertidae): Different histories but common traits. Chromosome Res. 2015, 23, 441–461. [Google Scholar] [CrossRef]
- Yamada, K.; Yamada, K.; Nishida-Umehara, C.; Matsuda, Y. Characterization and chromosomal distribution of novel satellite DNA sequences of the lesser rhea (Pterocnemia pennata) and the greater rhea (Rhea americana). Chromosome Res. 2002, 10, 513–523. [Google Scholar] [CrossRef]
- Yamada, K.; Nishida-Umehara, C.; Matsuda, Y. A new family of satellite DNA sequences as a major component of centromeric heterochromatin in owls (Strigiformes). Chromosoma 2004, 112, 277–287. [Google Scholar] [CrossRef]
- Axelsson, E.; Webster, M.T.; Smith, N.G.C.; Burt, D.W.; Ellegren, H. Comparison of the chicken and turkey genomes reveals a higher rate of nucleotide divergence on microchromosomes than macrochromosomes. Genome Res. 2005, 15, 120–125. [Google Scholar] [CrossRef]
- Warren, W.C.; Hillier, L.W.; Tomlinson, C.; Minx, P.; Kremitzki, M.; Graves, T.; Markovic, C.; Bouk, N.; Pruitt, K.D.; Thibaud-Nissen, F.; et al. A New Chicken Genome Assembly Provides Insight into Avian Genome Structure. G3 Genes. Genomes Genet. 2017, 7, 109–117. [Google Scholar] [CrossRef]
- Bolívar, P.; Guéguen, L.; Duret, L.; Ellegren, H.; Mugal, C.F. GC-biased gene conversion conceals the prediction of the nearly neutral theory in avian genomes. Genome Biol. 2019, 20, 5. [Google Scholar] [CrossRef]
- Ruiz-Ruano, F.J.; Navarro-Domínguez, B.; Camacho, J.P.M.; Garrido-Ramos, M.A. Characterization of the satellitome in lower vascular plants: The case of the endangered fern Vandenboschia speciosa. Ann. Bot. 2019, 123, 587–599. [Google Scholar] [CrossRef] [PubMed]
- Goes, C.A.G.; Dos Santos, N.; Rodrigues, P.H.D.M.; Stornioli, J.H.F.; Silva, A.B.D.; Dos Santos, R.Z.; Vidal, J.A.D.; Silva, D.M.Z.D.A.; Artoni, R.F.; Foresti, F.; et al. The Satellite DNA Catalogues of Two Serrasalmidae (Teleostei, Characiformes): Conservation of General satDNA Features over 30 Million Years. Genes 2022, 14, 91. [Google Scholar] [CrossRef] [PubMed]
- Rico-Porras, J.M.; Mora, P.; Palomeque, T.; Montiel, E.E.; Cabral-de-Mello, D.C.; Lorite, P. Heterochromatin Is Not the Only Place for satDNAs: The High Diversity of satDNAs in the Euchromatin of the Beetle Chrysolina americana (Coleoptera, Chrysomelidae). Genes 2024, 15, 395. [Google Scholar] [CrossRef] [PubMed]
- Gálvez-Galván, A.; Barea, L.; Garrido-Ramos, M.A.; Prieto, P. Highly divergent satellitomes of two barley species of agronomic importance, Hordeum chilense and H. vulgare. Plant Mol. Biol. 2024, 114, 108. [Google Scholar] [CrossRef]
- Patterson, S.A.; Morris-Pocock, J.A.; Friesen, V.L. A multilocus phylogeny of the Sulidae (Aves: Pelecaniformes). Mol. Phylogenetics Evol. 2011, 58, 181–191. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times. Mol. Biol. Evo. 2017, 34, 1812–1819. [Google Scholar] [CrossRef]
- Talbert, P.B.; Henikoff, S. What makes a centromere? Exp. Cell. Res. 2020, 389, 111895. [Google Scholar] [CrossRef]
- Rutkowska, J.; Lagisz, M.; Nakagawa, S. The long and the short of avian W chromosomes: No evidence for gradual W shortening. Biol. Lett. 2012, 8, 636–638. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhang, J.; Bachtrog, D.; An, N.; Huang, Q.; Jarvis, E.D.; Gilbert, M.T.P.; Zhang, G. Complex evolutionary trajectories of sex chromosomes across bird taxa. Science 2014, 346, 1246338. [Google Scholar] [CrossRef]
- Schmid, M.; Steinlein, C. The Hypermethylated Regions in Avian Chromosomes. Cytogenet. Genome Res. 2017, 151, 216–227. [Google Scholar] [CrossRef]
- Wójcik, E.; Smalec, E. Constitutive heterochromatin in chromosomes of duck hybrids and goose hybrids. Poult. Sci. 2017, 96, 18–26. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | Sex | Locality |
|---|---|---|---|
| N. brasilianum | Phalacrocoracidae | 1♀1♂ | Rio Grande, RS, Brazil |
| S. leucogaster | Sulidae | 1♀1♂ | Abrolhos, PE, Brazil |
| S. dactylatra | Sulidae | 1♀1♂ | Abrolhos, PE, Brazil |
| S. sula | Sulidae | 1♂ | Fernando de Noronha, BA, Brazil |
| F. magnificens | Fregatidae | 1♀ | Abrolhos, PE, Brazil |
| Satellite | Monomer Size | Abundance (F) | Abundance (M) | Ratio F/M | C + G (%) | A + T (%) |
|---|---|---|---|---|---|---|
| SleSat01 | 100 | 0.114238948 | 0.103363981 | 1.105210408 | 63 | 37 |
| SleSat02 | 190 | 0.044193639 | 0.041334463 | 1.069171739 | 54.7 | 45.3 |
| SleSat03 | 24 | 0.016987825 | 0.014847279 | 1.144170977 | 62.5 | 37.5 |
| SleSat04 | 17 | 0.004358942 | 0.0000005813 | 7498.180046 | 47.1 | 52.9 |
| SleSat05 | 145 | 0.000124061 | 0.000156155 | 0.794473001 | 55.2 | 44.8 |
| NbrSat01 | 113 | 0.016387741 | 0.019163133 | 0.855170235 | 66.4 | 33.6 |
| NbrSat02 | 2559 | 0.004029267 | 0.004085564 | 0.986220589 | 52.1 | 47.9 |
| NbrSat03 | 42 | 0.002012025 | 0.002329438 | 0.863738521 | 71.4 | 28.6 |
| NbrSat04 | 62 | 0.000820984 | 0.000918009 | 0.89430964 | 42.2 | 57.8 |
| NbrSat05 | 2118 | 0.000731359 | 0.000866913 | 0.843635922 | 61.7 | 38.3 |
| NbrSat06 | 862 | 0.000214127 | 0.000206287 | 1.03800201 | 38.4 | 61.6 |
| NbrSat07 | 20 | 0.000155798 | - | - | 60 | 40 |
| NbrSat08 | 77 | 0.0000995020 | 0.000116493 | 0.854148187 | 68.8 | 31.2 |
| Satellite | S. leucogaster | S. dactylatra | S. sula | N. brasilianum | F. magnificens |
|---|---|---|---|---|---|
| SleSat01 | centromeres of autosomes | 46 microchromosomes and the centromere of W | no signal | no signal | no signal |
| SleSat02 | 34 microchromosomes | 20 macrochromosomes and 34 microchromosomes | no signal | no signal | no signal |
| SleSat03 | centromeres of autosomes and Z2 | 52 microchromosomes | 4 microchromosomes and the first 8 pairs of macrochromosomes | no signal | no signal |
| SleSat04 | chromosome W | no signal | no signal | no signal | no signal |
| SleSat05 | 2 microchromosomes | 2 microchromosomes | 2 microchromosomes | no signal | no signal |
| NbrSat01 | no signal | no signal | no signal | 52 microchromosomes | no signal |
| NbrSat02 | no signal | no signal | no signal | 16 microchromosomes | no signal |
| NbrSat03 | no signal | no signal | no signal | 52 microchromosomes | no signal |
| NbrSat04 | no signal | no signal | no signal | 24 microchromosomes | no signal |
| NbrSat05 | no signal | no signal | no signal | 16 microchromosomes | no signal |
| NbrSat06 | no signal | no signal | no signal | 4 microchromosomes | no signal |
| NbrSat07 | no signal | no signal | no signal | W chromosome | no signal |
| NbrSat08 | no signal | no signal | no signal | no signal | no signal |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pozzobon, L.C.; dos Santos, N.; Utsunomia, R.; Porto-Foresti, F.; Cioffi, M.d.B.; Kretschmer, R.; de Freitas, T.R.O. Satellite DNA Mapping in Suliformes (Aves): Insights into the Evolution of the Multiple Sex Chromosome System in Sula spp. Genes 2025, 16, 633. https://doi.org/10.3390/genes16060633
Pozzobon LC, dos Santos N, Utsunomia R, Porto-Foresti F, Cioffi MdB, Kretschmer R, de Freitas TRO. Satellite DNA Mapping in Suliformes (Aves): Insights into the Evolution of the Multiple Sex Chromosome System in Sula spp. Genes. 2025; 16(6):633. https://doi.org/10.3390/genes16060633
Chicago/Turabian StylePozzobon, Luciano Cesar, Natália dos Santos, Ricardo Utsunomia, Fábio Porto-Foresti, Marcelo de Bello Cioffi, Rafael Kretschmer, and Thales Renato Ochotorena de Freitas. 2025. "Satellite DNA Mapping in Suliformes (Aves): Insights into the Evolution of the Multiple Sex Chromosome System in Sula spp." Genes 16, no. 6: 633. https://doi.org/10.3390/genes16060633
APA StylePozzobon, L. C., dos Santos, N., Utsunomia, R., Porto-Foresti, F., Cioffi, M. d. B., Kretschmer, R., & de Freitas, T. R. O. (2025). Satellite DNA Mapping in Suliformes (Aves): Insights into the Evolution of the Multiple Sex Chromosome System in Sula spp. Genes, 16(6), 633. https://doi.org/10.3390/genes16060633