1. Introduction
Saussurea DC. is one of the largest genera in the Asteraceae family, with more than 500 species found in the northern hemisphere [
1].
S. obvallata (DC.) Edgew., a perennial hermaphrodite herb of the Asteraceae family, is endemic to the Qinghai–Tibet Plateau (QTP), thriving at altitudes between 3200 and 4700 masl.
S. obv is commonly found in alpine screes, alpine meadows, rocky slopes, shrubs, and near streams [
2,
3]. The stones and shrubs can shield
S. obv from strong wind, allowing it to achieve an average height of 50–80 cm.
S. obv blooms from July to September. It is a typical “greenhouse plant”. The light-yellow bracts envelop flowers, thus protecting them from UV light and retaining heat at night (
Figure 1A), attracting insects for cross-pollination. By September and October, the fruits will gradually mature, and the bracts will open (
Figure 1C). The fruits with pappi (
Figure 1D) are dispersed by wind and grow in suitable habitats [
4]. During winter, the above-ground parts wither, and the roots will regenerate in the following spring [
5].
Over the past years, global climate change and human alterations have causedhabitat fragmentation and a dramatic decline in distribution. In 2009, the Science and Technology Department of Xizang Autonomous Region classified it as a Class III endangered medicinal plant [
6]. In 2017, Fei et al. investigated the medicinal plant resources of Sejira Mountain, and
S. obv was suggested as a Class II endangered species [
7]. In India, because of its wide application in medicine, decoration, and social economy [
8], coupled with the weak ecological awareness of residents and tourists,
S. obv is facing over-harvesting and, therefore, has been listed as an endangered species and one of the highest priorities for conservation programs by the Conservation Assessment Management Plan (CAMP) and Convention of International Trade in Endangered Species of Wild Fauna and Flora (CITES).
More than 40
S. species have been used as traditional medicinal plants in China [
9,
10,
11], and their phytochemicals and pharmacological activities havealso been investigated for many years [
12,
13]. The main compounds identified from
S. species are terpenoids, flavonoids, lignans phenylpropanoids, and polysaccharides, and they were mainly used for anti-tumor, antibacterial, anti-inflammatory, and anti-ulcer properties.
S. medusa, containing neoechinulin A, arctigenin, epigenin, and luteolin, has been used to protect patients from inflammation, ultraviolet radiation, cold, hypoxia, and skin photo-aging [
14,
15,
16].
S. costus is an endangered species, traditionally used for a variety of diseases, showing potent effects on carminative, expectorant, anti-arthritic, and antiseptic [
17].
S. involucrata, mainly distributed in Xinjiang, China, was found to have neuroprotection effect, ischemia/reperfusion injury protection, adipogenesis suppression, anti-tumor effect, anti-inflammation, cardio-protective effect, and other pharmaceutical values [
18,
19].
Due to its antipyretic, antidotal, anti-inflammatory, and analgesic effects,
S. obv has been used as a traditional medical plant in China, India, and Pakistan [
20,
21,
22,
23,
24]. It is recorded in ancient Tibetan medicine books “The Four Medical Tantras”, “Crystal Materia Medica”, “The Stainless Crystal Mirror: A Tibetan Materia Medica”, and so on. Over 130 phytochemicals have been reported in
S. obv, including flavonoids, terpenoids, fatty acids, and alkaloids [
25,
26,
27,
28,
29,
30,
31,
32,
33]. These metabolites not only play an important role in plant response to environmental stressbut also are the pharmacological basis of medicinal plants.
Metabolomics studies all metabolites in the biological body. Compared with genomics and proteomics, it reflects the immediate biochemical state in the biological body. In addition, metabolomics can provide comprehensive information about the types and contents of various metabolites and quickly obtain the changes of metabolites and their change rules in different environments, different growth periods, different parts, or under certain specific stimuli [
34,
35]. Metabolomics technology has been widely used because of its relatively low price and labor-saving [
36].
The primary metabolites of plants, such as proteins, lipids, and polysaccharides, are essential substances to maintain basic life activities. Secondary metabolites, including flavonoids, terpenes, alkaloids, coumarins, and other small-molecule compounds, not only play a key role in plant stress responses, such as UV protection, antibacterial, and resistance to pests and diseases, but also have important value for human health [
37].
UPLC-Q-TOF-MS/MS is a liquid chromatography-mass spectrometry combined technology, which has high sensitivity, good separation effect, fast detection speed, and relatively accurate mass charge ratio for detected ions.Therefore, it is widely used in natural product analysis and metabolomics research [
38,
39].
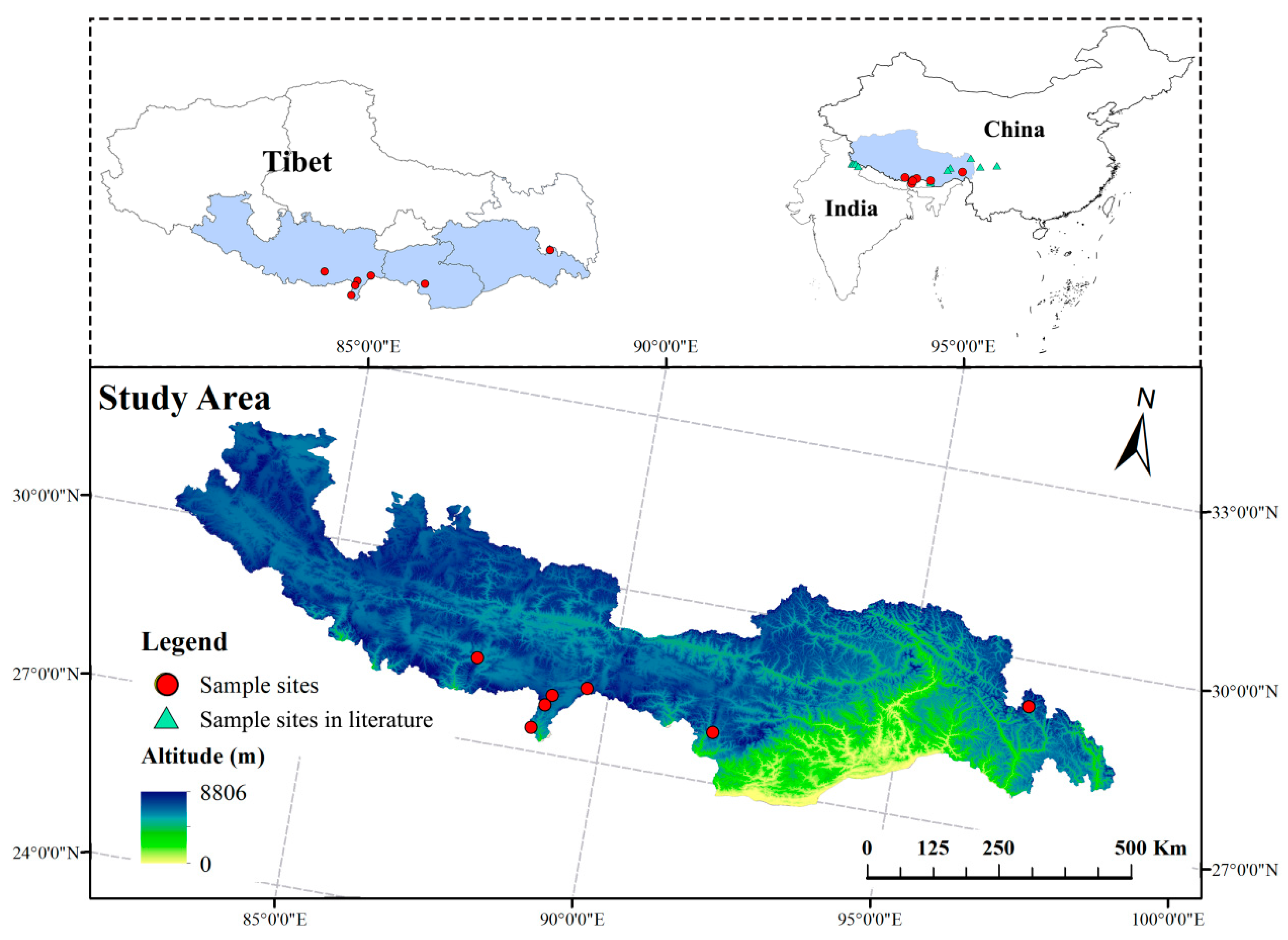
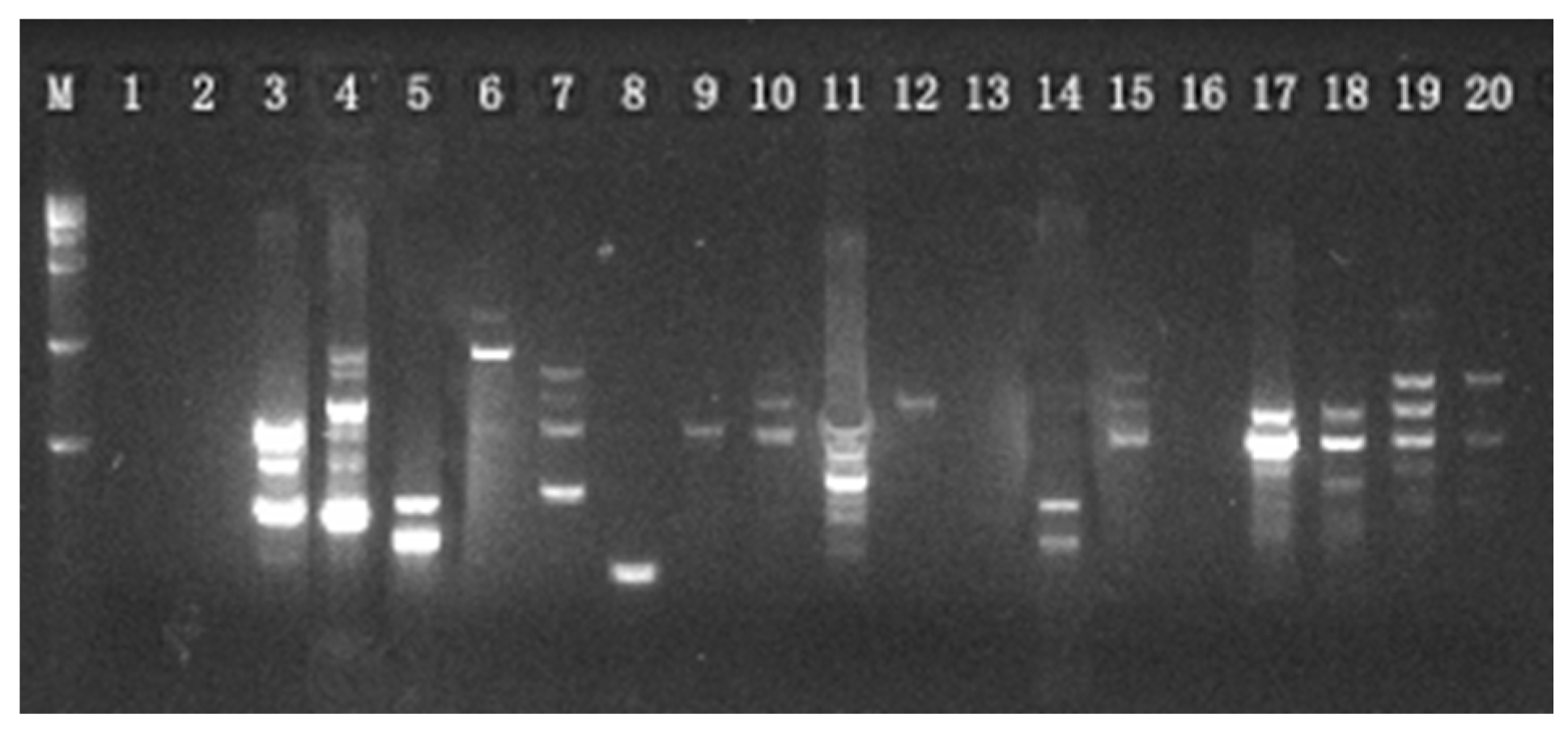
In this study, the ISSR marker was used to assesspopulation genetic variation, and UPLC-Q-TOF-MS/MS was used to determinethe metabolism. Analyzing the genetic diversity and differential metabolites of different populationsprovidesbaseline data for the conservation, development, and utilization of S. obv.
4. Discussion
Genetic diversity is a critical indicator of a species’ ability to survive, reproduce, and evolve. Species with higher genetic diversity would better adapt to environmental changes [
47,
48,
49,
50,
51]. Wild species of the QTP are threatened by extreme environments, global warming, and inadequate conservation [
52,
53]. Moreover, the habitats of
S. obv are relatively moist and densely vegetated, which makes it easy to attract herbivores. Understanding the genetic diversity of
S. obv is essential for its survival, reproduction, development, and utilization, and it can also play a good role in umbrella protection for other species within the habitats.
Genetic diversity assessments are crucial for designing effective conservation strategies. Nei’s gene diversity (H) among seven populations was 0.3027, and Shannon’s information index (I) was 0.4535. At the population level, these values were lower (H: 0.0834–0.2698; I: 0.1454–0.4145). The genetic differentiation index (Fst) of seven populations was 0.114. Comparatively, studies on
S. species using different molecular markers revealed divergent patterns. Hu et al. [
54] investigated the genetic diversity of
S. involucrata using simple sequence repeat (SSR) molecular markers, revealing high genetic diversity (H = 0.470 and I = 0.837) among 11 populations in Xinjiang, China. Another genetic diversity analysis of
S. involucrata using ITS and chloroplast fragments sequences found higher total nucleotide diversity in ITS1-4 region (π = 6.13) than that of cpDNA (π = 1.26), while the total haplotype diversity in the ITS1-4 region (Hd = 0.813) was similar to that of cpDNA (Hd = 0.895) [
55]. Twenty
S. medusa populations in the Qilian Mountains, China, also showed similar genetic diversity (H = 0.2757 and I = 0.4237) yet strong genetic differentiation (Gst = 0.4926), as sequence-related amplified polymorphism (SRAP) markerswere used [
56]. The ISSR marker was used to study the genetic diversity of
S. dorogostaiskii, a globally threatened species, and the results show low genetic diversity (H = 0.17, I = 0.25) but high genetic differentiation (Gst = 0.49) [
57]. Semwal et al. [
40] studied the genetic diversity of five populations of
S. obv using an ISSR marker in India, revealing low genetic diversity (H = 0.27 and I = 0.38) due to habitat fragmentation. A broad review [
58] comprising 108 entries revealed mean within-population H = 0.214 ± 0.117 and mean Gst = 0.29 ± 0.21. Generally, the genetic diversity and differentiation of
S. obv were relatively low.
There was no significant correlation between genetic diversity and environmental factors, but this does not necessarily mean that the growth and reproduction of
S. obv are not restricted by the environment. Instead, it may reflect its adaptation to specific microhabitats. The habitats of different populations are similar, with altitudes between 4000–4500 m, the presence of streams, diurnal temperature variation from June to September, and bushes or boulders to withstand strong winds. Therefore, this species may have strict environmental requirements, which further limit distribution. Moreover, genetic diversity is also associated with life history traits (life form, geographical range, breeding system, seed dispersal, and successional status) and sampling strategies [
58]. The genetic diversity of
S. obv may be shaped by the combined effects of several factors: short-lived perennial life form, endemic to the QTP, mixed breeding system, and seed dispersal by wind. Therefore, further study should incorporate a broader range of sampling sites and life history traits, which would provide valuable insights into the conservation strategies of
S. obv.
However, the ISSR marker relies on PCR products to analyze genetic diversity. Thus, it cannot obtain information such as gene mutation, and it is difficult to distinguish different bands with very small base number differences. Therefore, the genetic differentiation between different populations remains to be further explored. Chloroplast genome sequencing has been widely applied in phylogenetic studies of the Asteraceae family. According to GenBank (
https://www.ncbi.nlm.nih.gov/, accessed on 18 March 2025), the chloroplast genomes of more than 100
S. species have been sequenced, and several highly variable regions have been identified [
59,
60]. This provides new insights into investigating the genetic diversity of
S. obv.
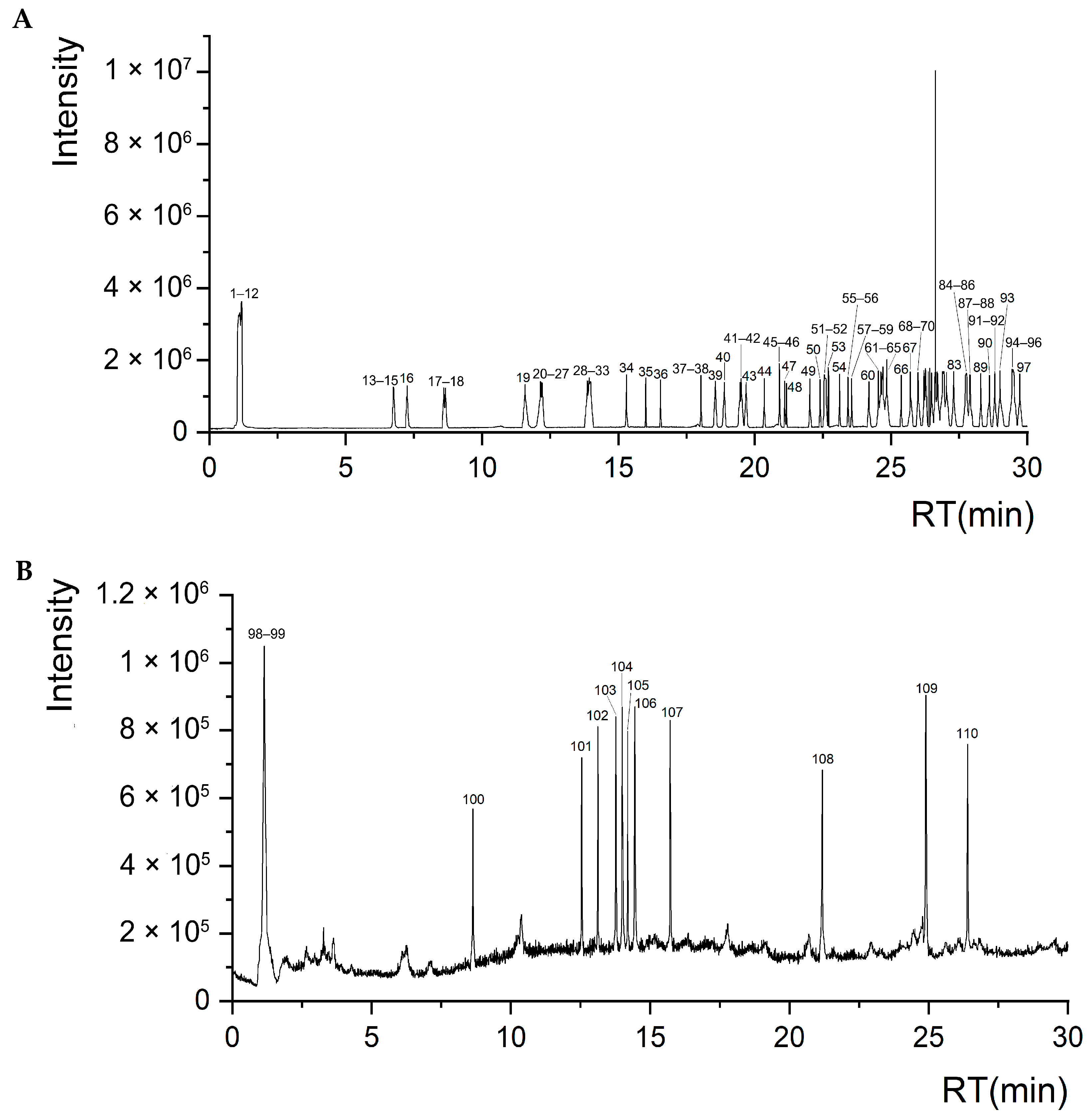
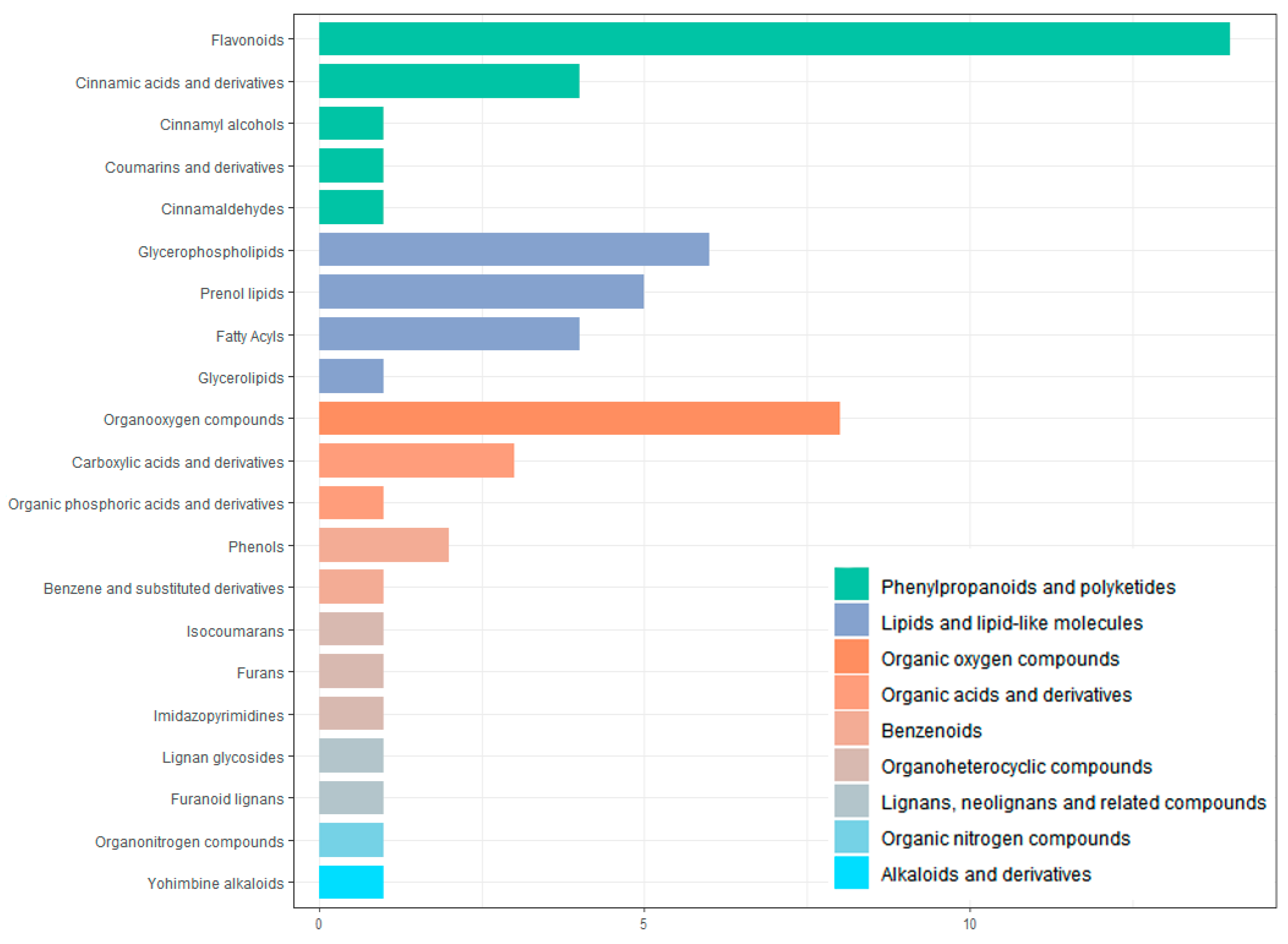
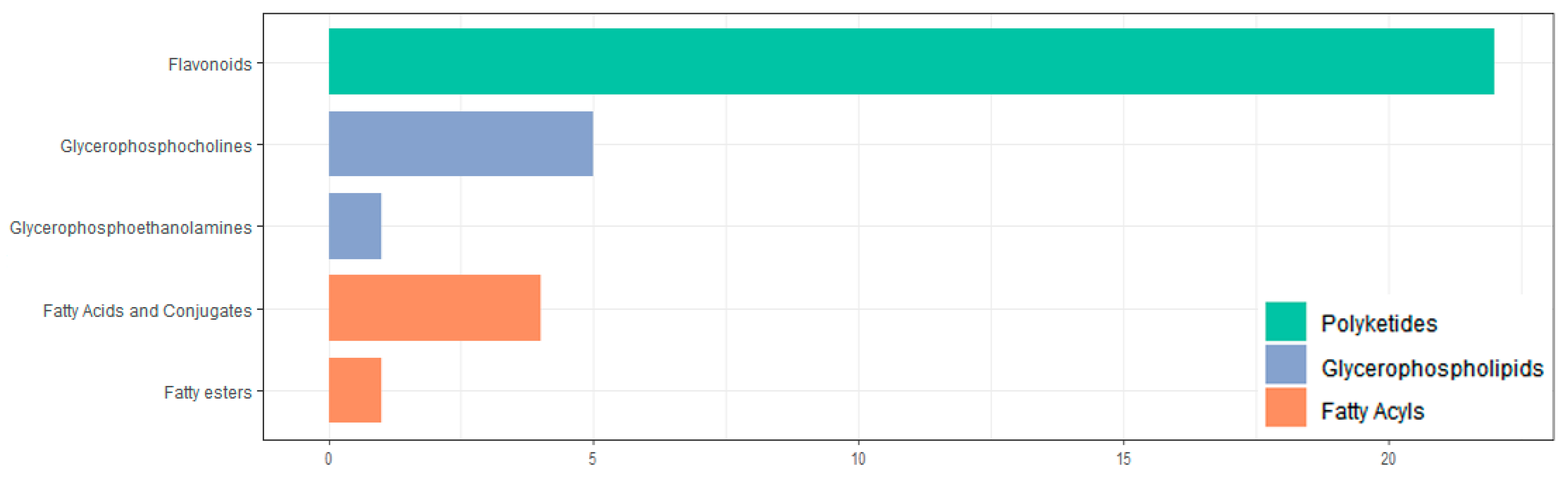
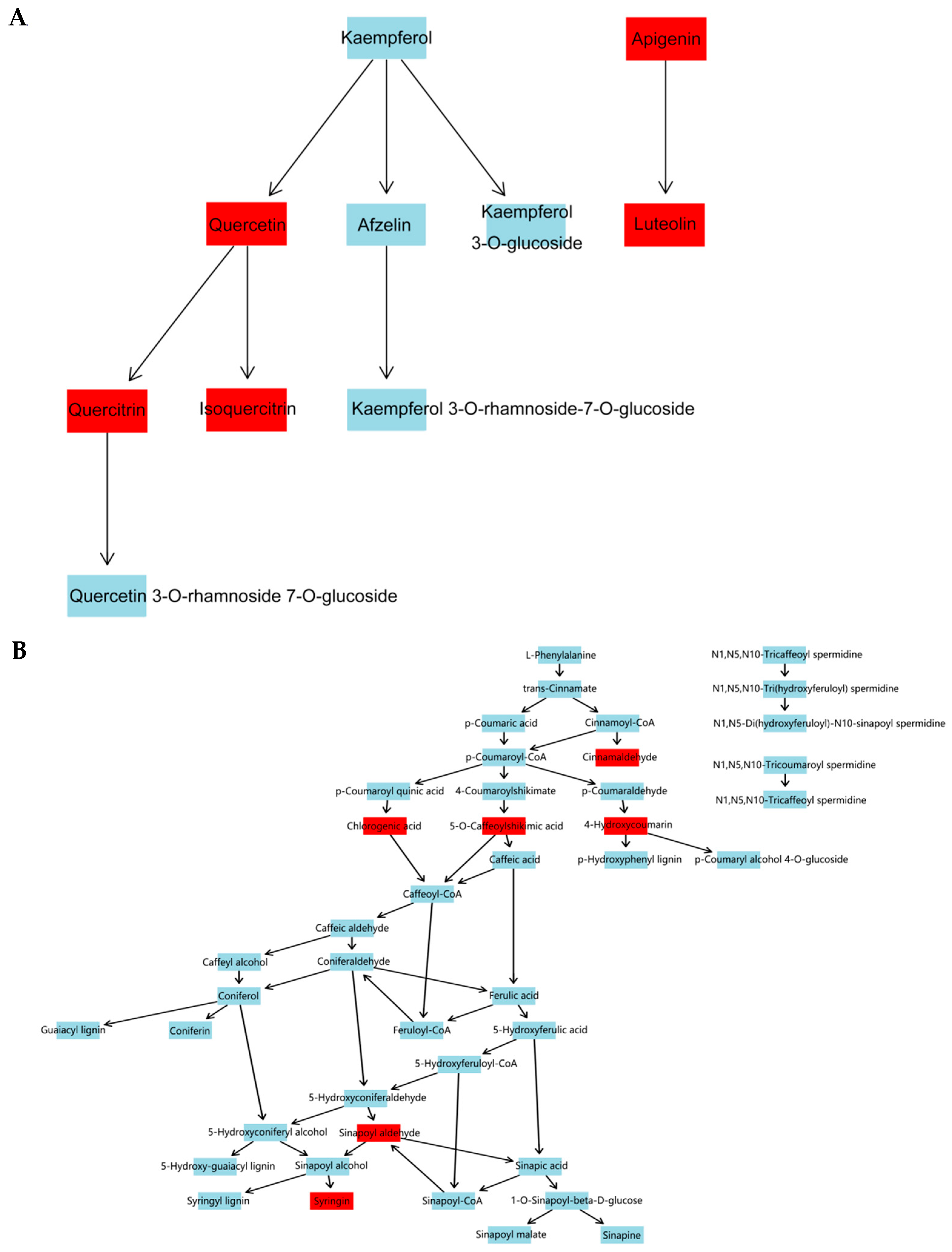
In this study, UPLC-Q-TOF-MS/MS was used, and 110 components were identified from the ethanol extract of S. obv, among which flavonoids and phenylpropanoids were the two most abundant categories.
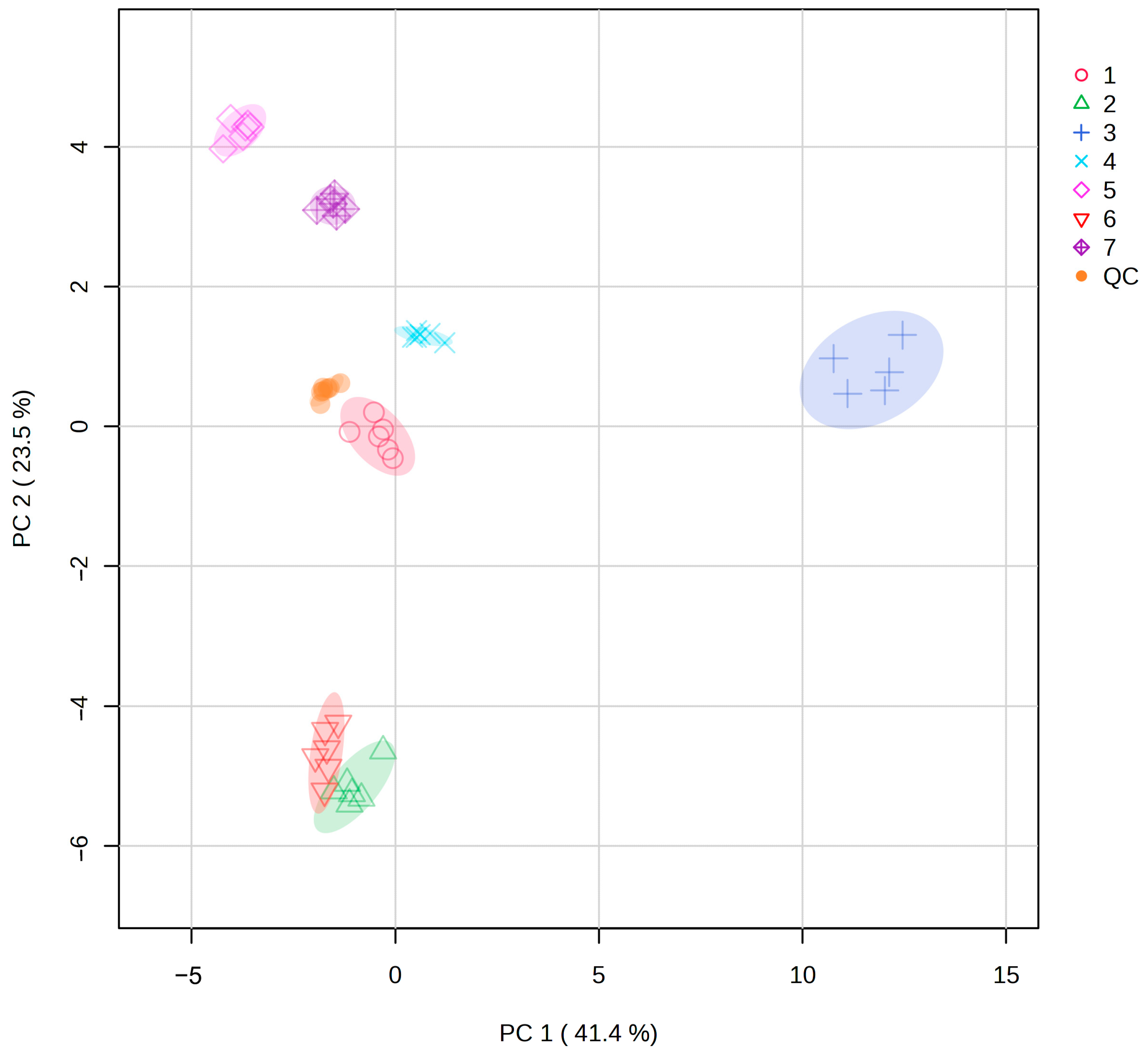
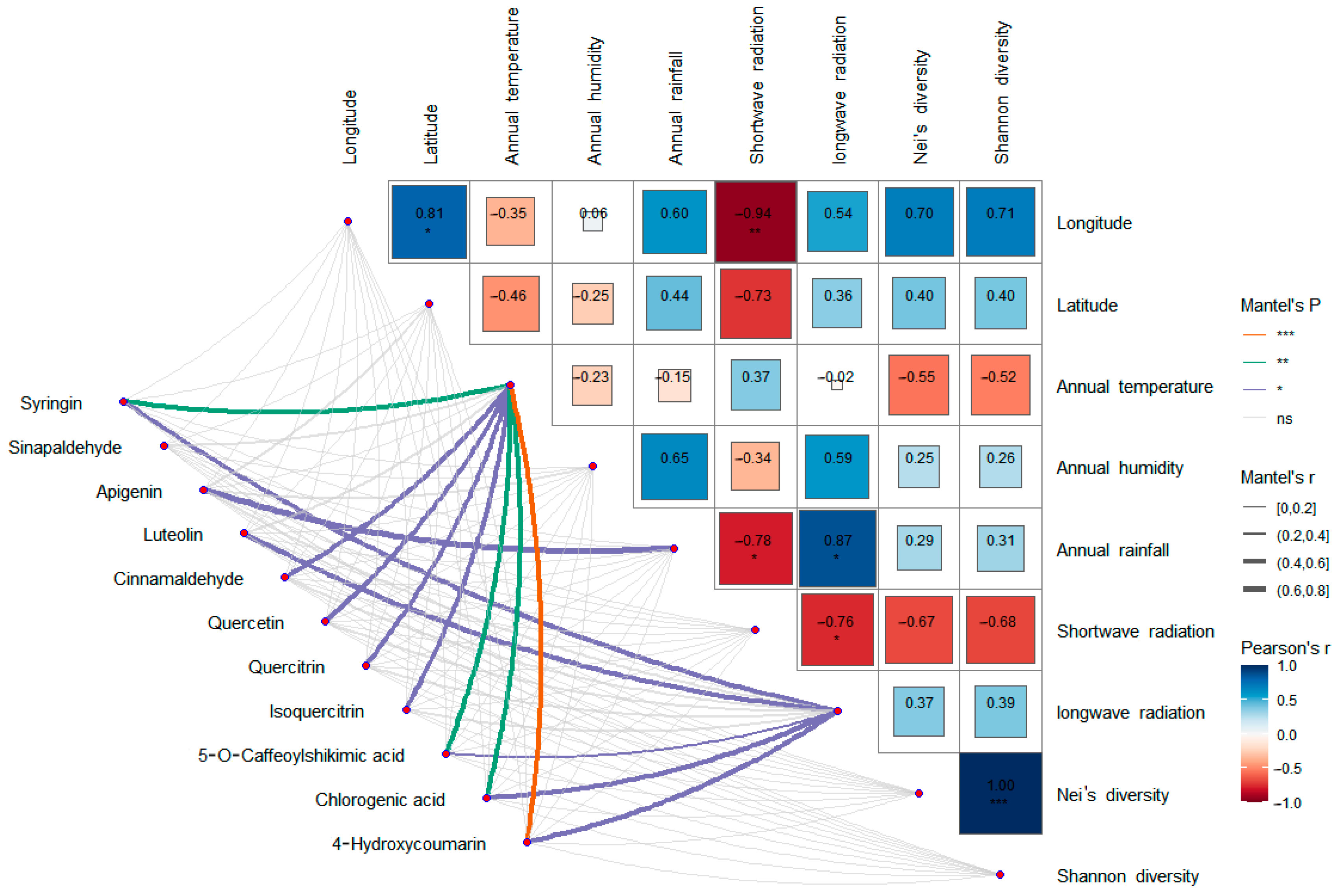
According to the PCA analysis, there were significant differences in metabolites among different populations. Correlation analysis with environmental factors and genetic diversity showed that the differences in metabolites among populations were not significantly related to geographical distance or genetic diversity. The metabolites of Population 2 and Population 6 were the most similar. In terms of geographical distance, Sample 2 was close to Sample 1, Sample 3, and Sample 4 but far from Sample 6, which further proved that there was no significant correlation between metabolic differences among populations and geographical distance.
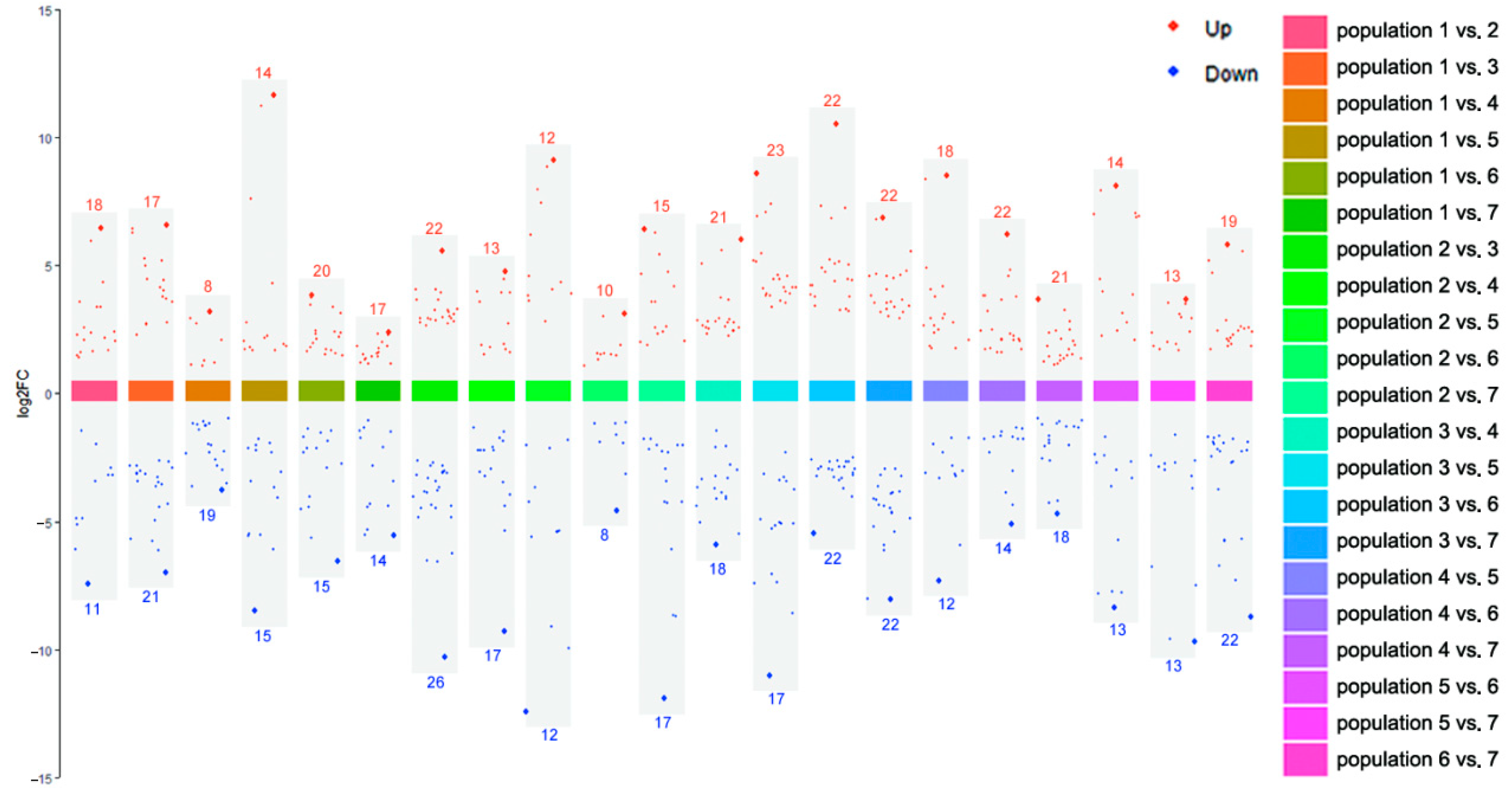
Pairwise comparison of metabolites showed that the number of upregulated differential metabolites ranged from 8 to 23, and downregulated differential metabolites ranged from 8 to 26. There was no significant difference in the number of differentiated metabolites between upregulated and downregulated groups, but the magnitude of variation was different. Interestingly, Population 5, which exhibited the lowest genetic diversity, also showed the most pronounced metabolic differentiation compared to other populations. Among all the differential metabolites, the upregulation of arctiin was the highest (Population 1 vs. Population 5, log2FC = 11.559), and the downregulation of Curdionolide B was the highest (Population 2 vs. Population 5, log2FC = −12.486). Among the environmental factors, Sample 5 had the highest annual mean temperature, lower annual mean precipitation and annual mean moisture content, and stronger short-wave radiation and weaker long-wave radiation, indicating that Sample 5 had greater degrees of high temperature, drought, and temperature fluctuation than other sample sites, which likely contributed to its distinct metabolic profile. In follow-up studies, we should pay more attention to Population 5.
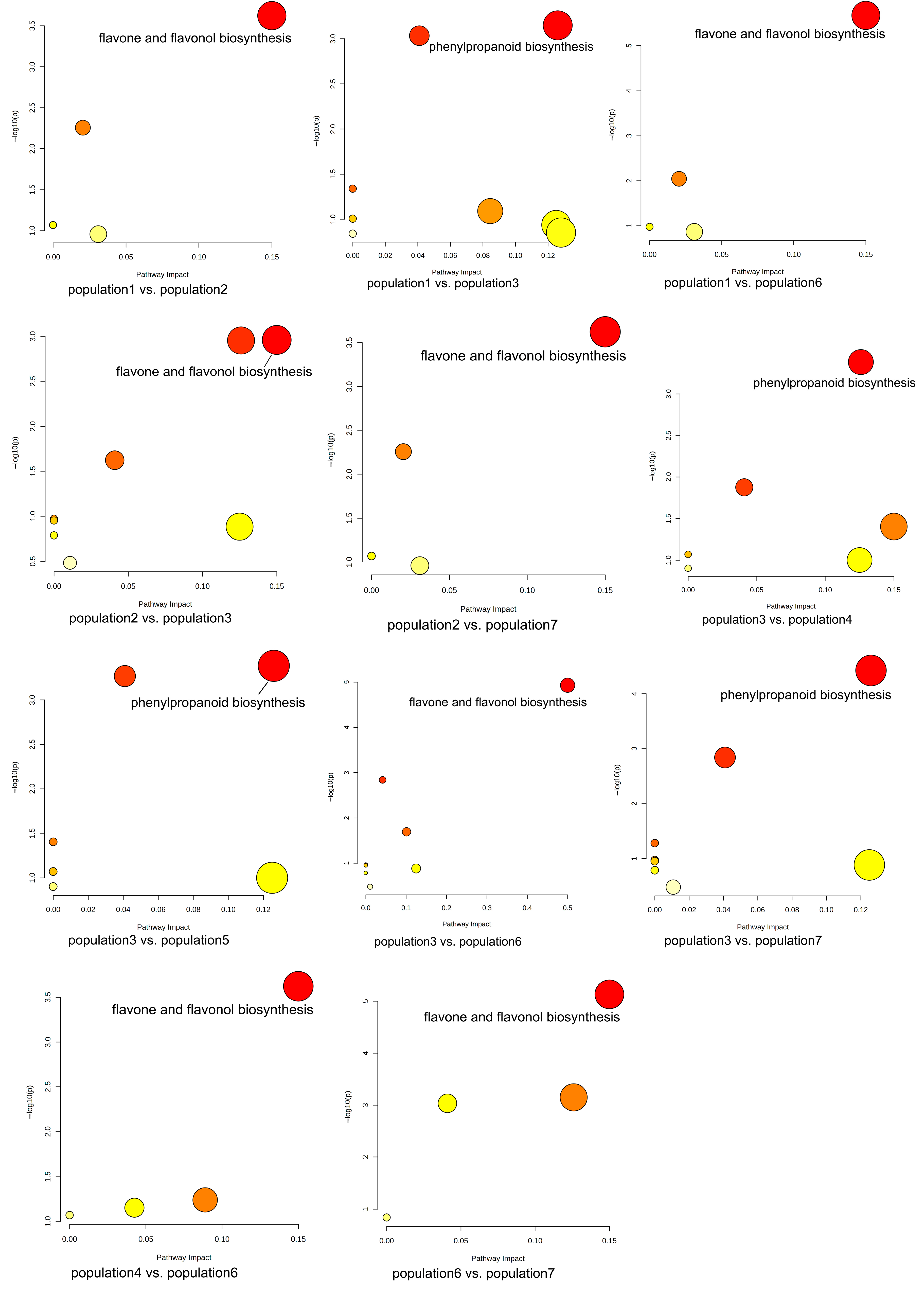
During the growth of plants, a metabolite does not exist independently but interacts with a variety of other metabolites and is regulated by the environment, genes, transcription, and other factors, thus forming a complex metabolic network. Annotation and KEGG pathway enrichment showed that the differential metabolites were mostly flavonoids and related to two pathways: flavone and flavonol biosynthesis and phenylpropanoid biosynthesis. Flavonoids are secondary metabolites commonly foundin plants, and they are also important medicinal substances in medicinal plants, usually with anti-inflammatory, antibacterial, antioxidant, anti-tumor, cardiovascular protection, and other effects [
61,
62,
63,
64]. Flavonoids are also important substances for plants to cope with environmental stress, and low temperature and ultraviolet radiation can promote the accumulation of flavonoids [
65,
66]. Phenylpropanoids are a class of natural products with one or more C6-C3 groups, which have good potential in anti-oxidation, anti-tumor, anti-virus, regulation of blood sugar, and protection of ultraviolet radiation damage [
67,
68]. Phenylpropanoids can enhance plant resistance to insect pests, cold resistance, oxidation resistance, and mechanical damage [
69,
70]. Some volatile phenylpropanoids, such as benzyl alcohol, benzyl acetate, methyl benzoate, and phenylacetaldehyde, can attract pollinating insects and play an important role in plant reproduction [
71]. The biosynthesis of phenylpropanoids is regulated by biological and abiotic factors. Studies have shown that low temperature can promote the accumulation of lignin and anthocyanins in the phenylpropanoids biosynthesis pathway [
72,
73,
74], and lignin can enhance the toughness of the plant cell wall to cope with the mechanical damage caused by low temperature. Anthocyanins have good antioxidant capacity, which can reduce ROS accumulation and alleviate oxidative damage caused by low-temperature stress on plants. In addition, syringin and other phenylpropanoids were also affected by drought stress [
75]. Metal ion concentration (including copper, iron, zinc, manganese, and sodium) [
76] and salt stress [
77] can also affect the pathways related to flavonoids and phenylpropanoids and then promote or inhibit the expression of flavonoids and phenylpropanoids.
Among the differential metabolites of different populations, 11 differential metabolites related to these two pathways were screened out, including syringin, sinapaldehyde, apigenin, luteolin, cinnamaldehyde, quercetin, quercitrin, isoquercetin, 5-O-caffeoylshikimic acid, chlorogenic acid, and 4-hydroxycoumarin. Correlation analysis of these 11 differential metabolites with environmental factors and genetic diversity showed that there was no significant correlation between the expression levels of these 11 differential metabolites and genetic diversity, latitude, and longitude, indicating that the metabolites of
S. obv were mainly affected by environmental factors rather than genetic factors. Also, there is no obvious spatial structure on a large scale. Among the environmental factors, the annual mean temperature had the greatest influence on the differential metabolites. There were eight differential metabolites (syringin, cinnamaldehyde, quercetin, quercetin, isoquercetin, 5-O-cafeoylshikimic acid, chlorogenic acid, and 4-hydroxycoumarin) that were significantly correlated with the annual mean temperature, followed by the annual mean long-wave radiation. Five different metabolites (syringin, luteolin, 5-O-caffeoylshikimic acid, chlorogenic acid, and 4-hydroxycoumarin) were significantly correlated with the annual mean long-wave radiation amount, suggesting that the different metabolites were mainly related to temperature and diurnal temperature variation. The responses of plant secondary metabolite accumulation to temperature have been proven [
78], while the mechanisms, which are how temperature and diurnal temperature variation affect the above metabolites, remain to be studied.
Although the mean annual precipitation and mean annual moisture content of each sample site were significantly different, there was almost no significant correlation between the two factors and the expression levels of different metabolites, except for apigenin. This may be due to the fact that S. obv grows near waterfalls or streams. Even when precipitation is low, the water flow can also make up for the lack of precipitation. S. obv mainly grows in summer and autumn. During this period, the QTP has higher temperature and precipitation, leading to abundant river water, so S. obv may not be susceptible to drought stress.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}