
Identification and Expression Analysis of CCCH Zinc Finger Family Genes in Oryza sativa

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of CCCH Proteins in Rice
2.2. Phylogeny and Protein Structure Analysis of the OsC3H Family Members
2.3. OsC3H Family Genomic Localization, Gene Duplication, and Synteny Analysis
2.4. Promoter Analysis of CCCH Genes in Rice
2.5. Analysis of OsC3H Gene Expression
2.6. Haplotype Analysis of OsC3Hs
3. Results
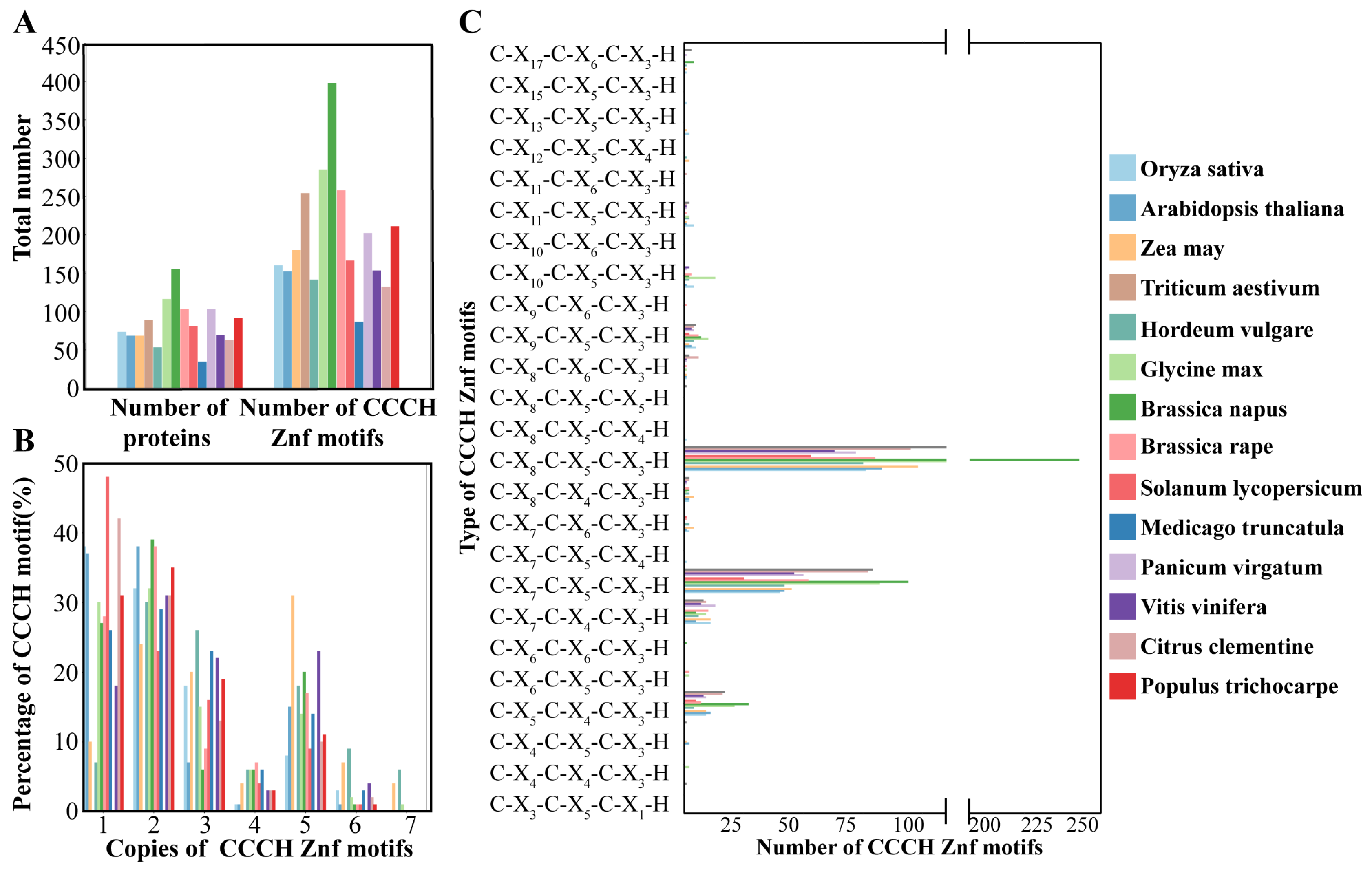
3.1. Identification of CCCH Zinc Finger Proteins in Rice
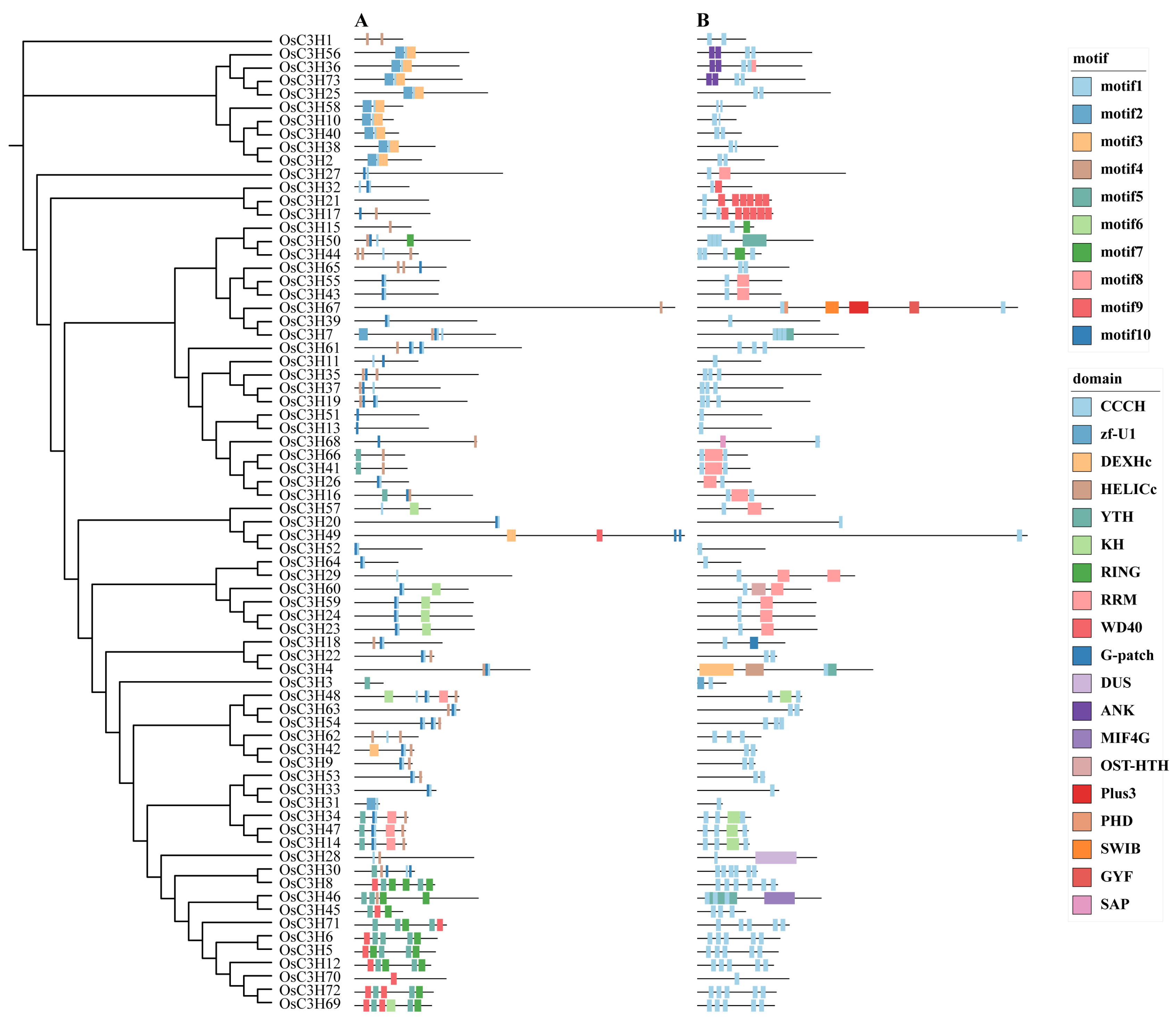
3.2. Phylogenetic Relationships, Protein Structures, and Conserved Domains of OsC3H Genes
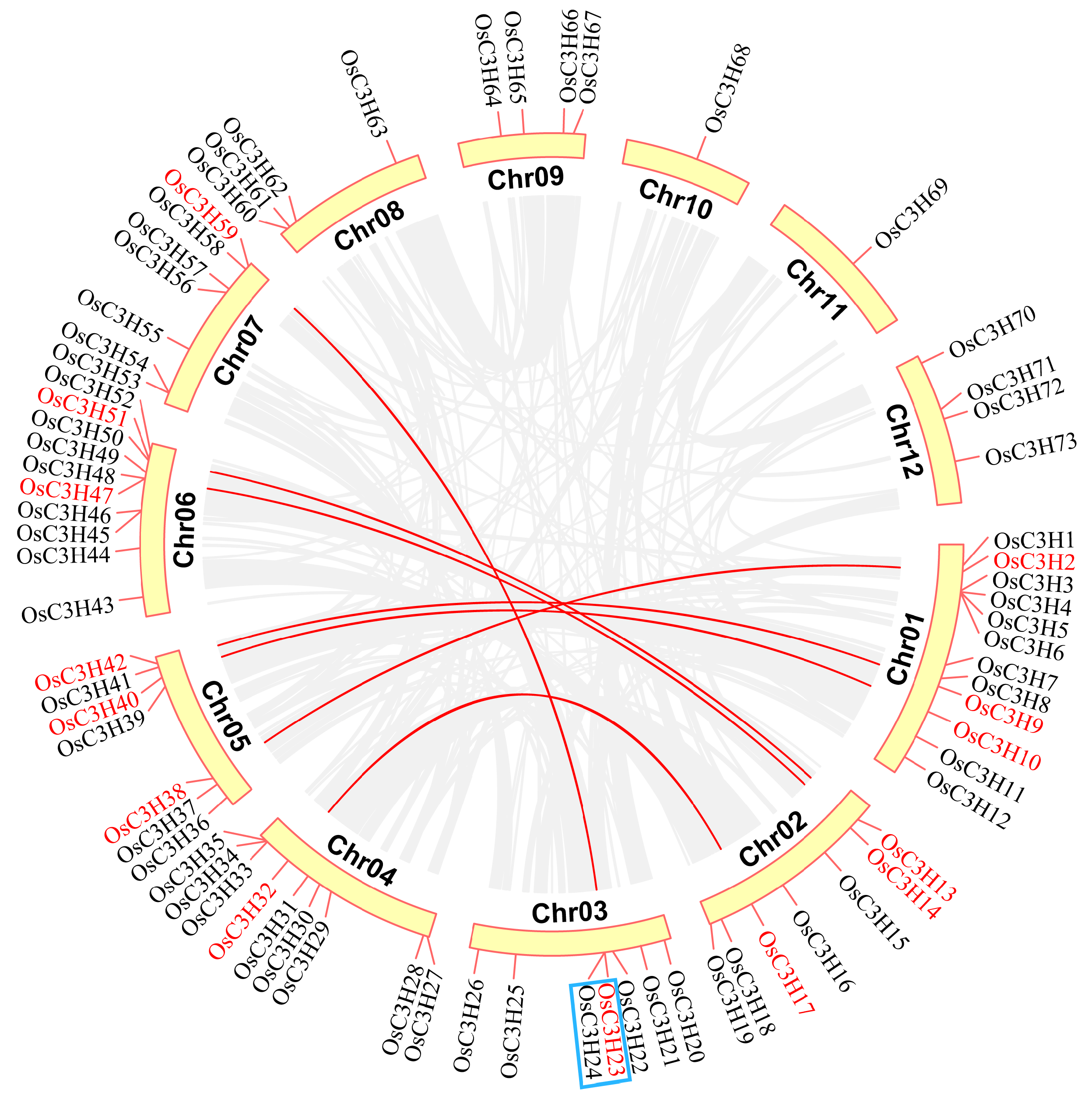
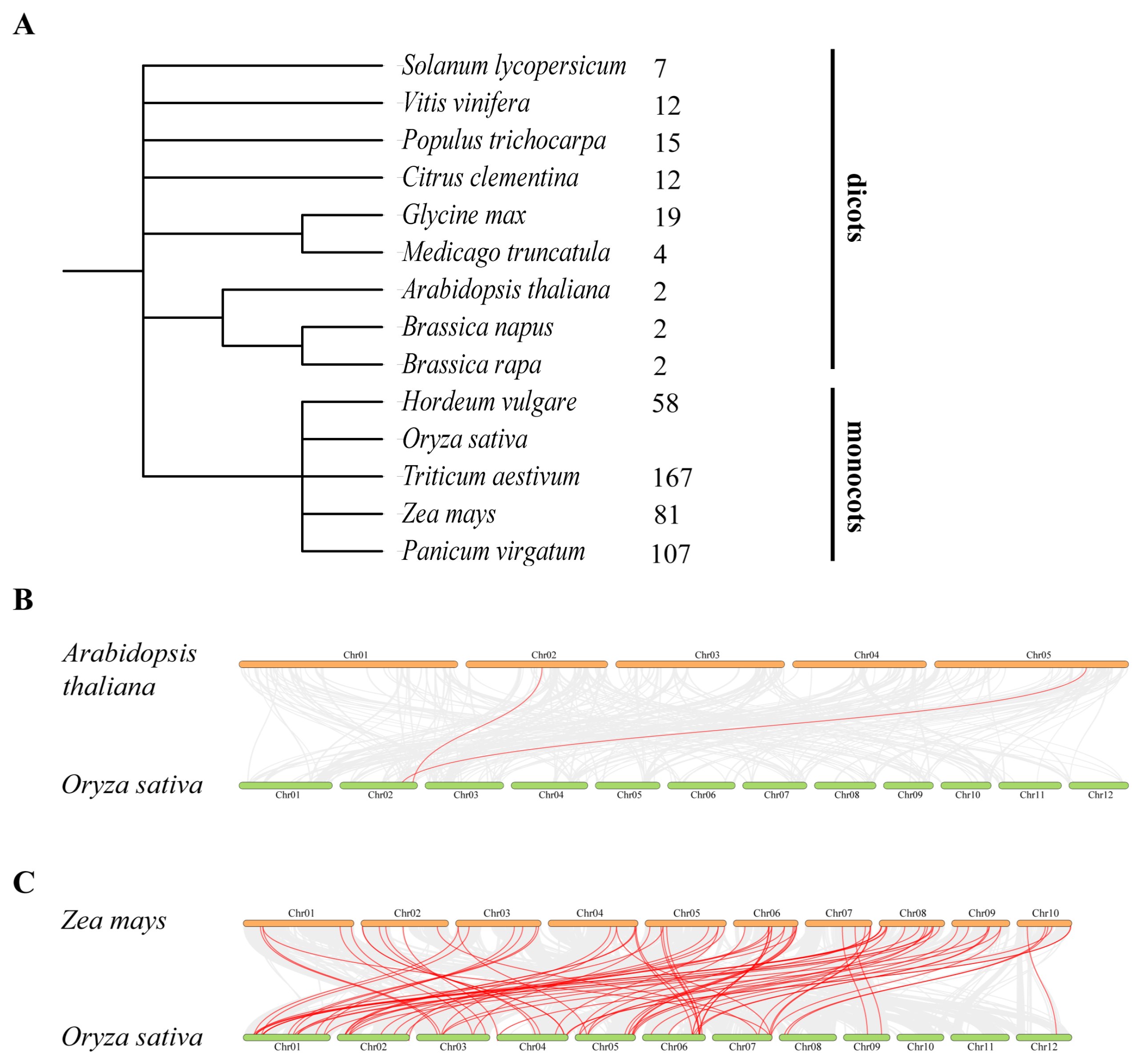
3.3. Chromosomal Distribution and Gene Duplication of OsC3H Genes
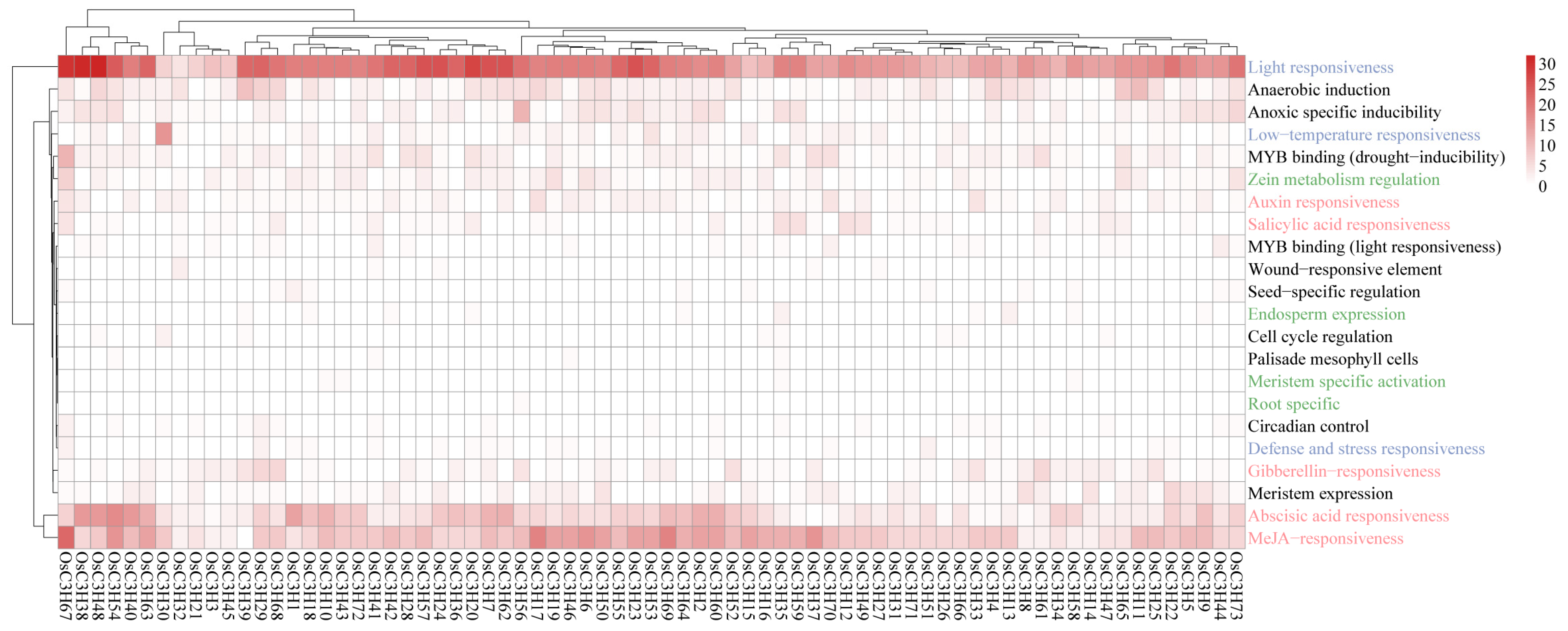
3.4. Cis-Regulatory Elements Analysis in OsC3H Genes
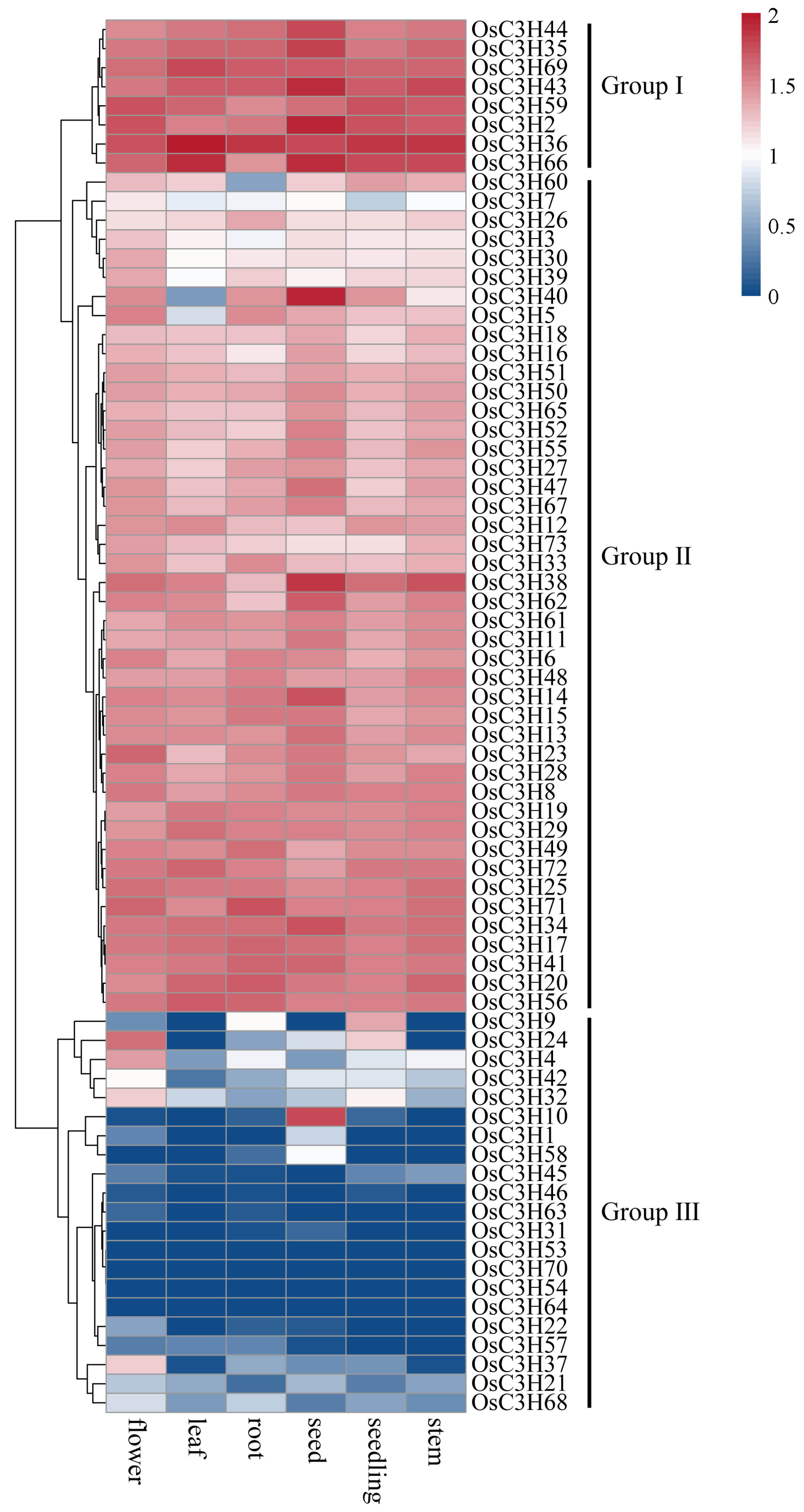
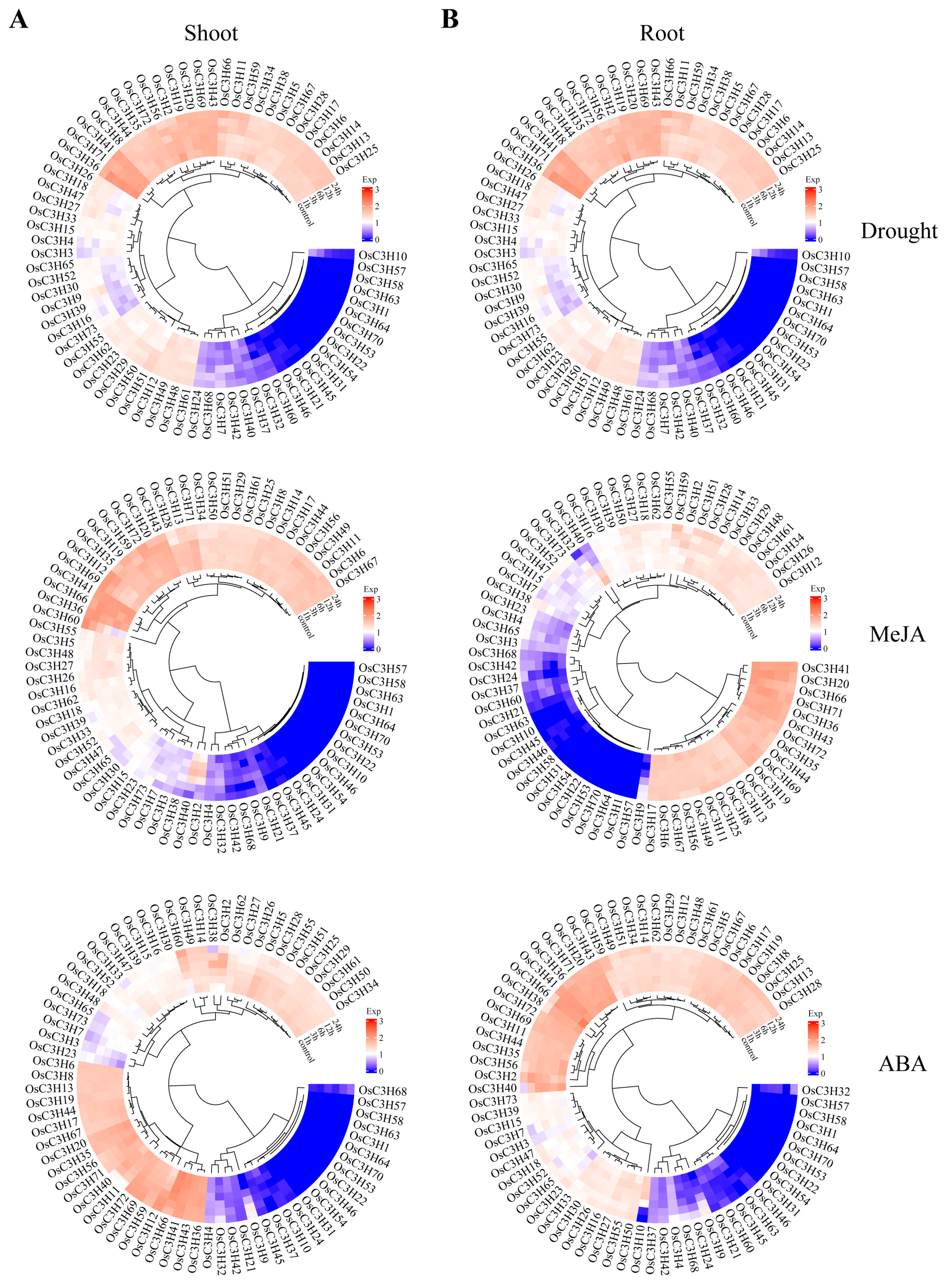
3.5. Temporal–Spatial and Stress-Induced Expression Pattern Analysis of OsC3Hs
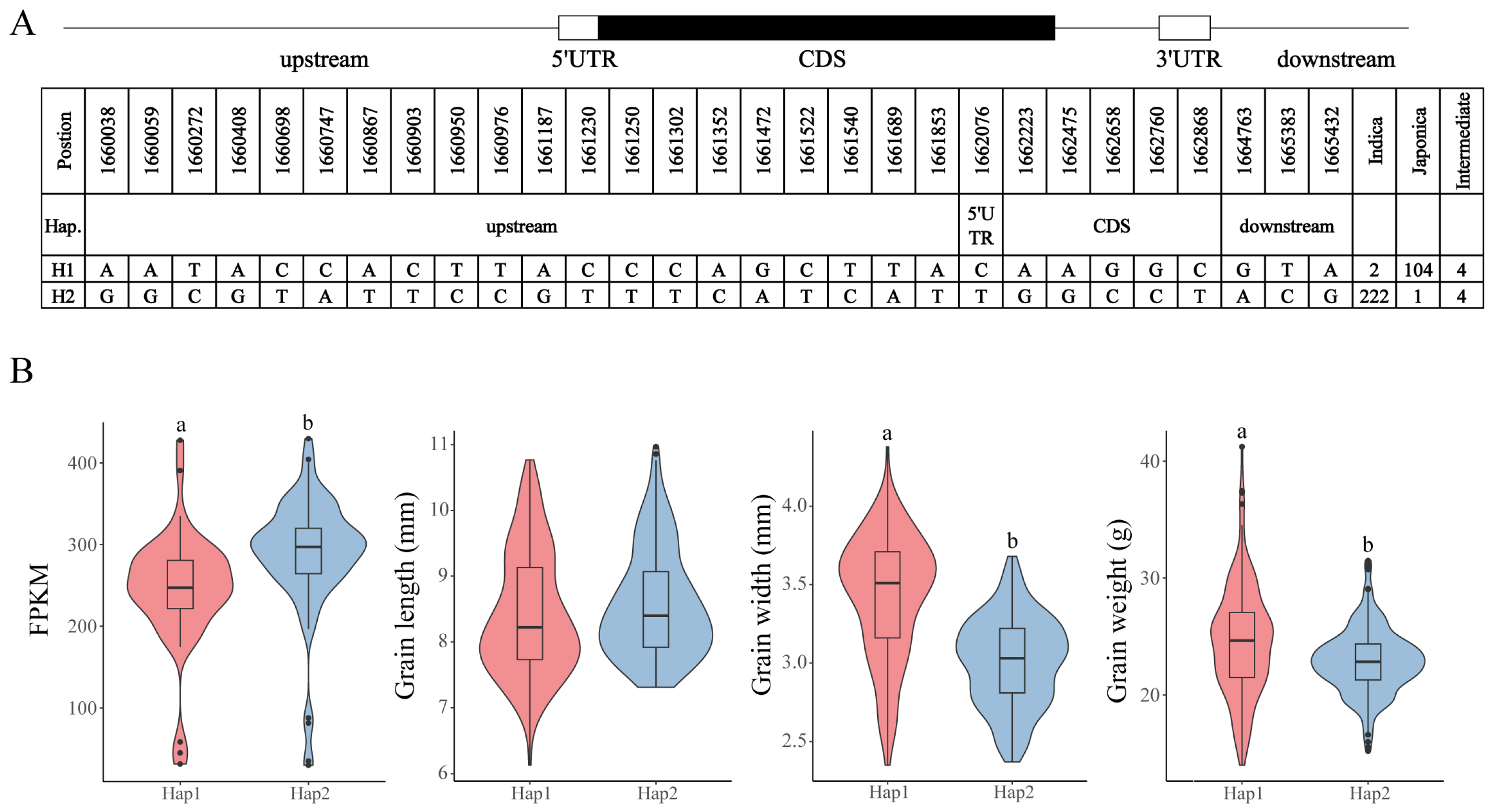
3.6. Analysis of the CCCH Haplotype
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berg, J.M.; Shi, Y. The Galvanization of Biology: A Growing Appreciation for the Roles of Zinc. Science 1996, 271, 1081–1085. [Google Scholar] [CrossRef] [PubMed]
- Laity, J.H.; Lee, B.M.; Wright, P.E. Zinc Finger Proteins: New Insights into Structural and Functional Diversity. Curr. Opin. Struct. Biol. 2001, 11, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, R.; Xie, Y.; Zhao, S.; Yan, M.; Sun, N.; Zhan, Y.; Li, F.; Yu, S.; Feng, Z.; et al. Association of GhGeBP Genes with Fiber Quality and Early Maturity Related Traits in Upland Cotton. BMC Genom. 2024, 25, 1058. [Google Scholar] [CrossRef] [PubMed]
- Ciftci-Yilmaz, S.; Mittler, R. The Zinc Finger Network of Plants. Cell. Mol. Life Sci. 2008, 65, 1150–1160. [Google Scholar] [CrossRef]
- Cheng, X.; Cao, J.; Gao, C.; Gao, W.; Yan, S.; Yao, H.; Xu, K.; Liu, X.; Xu, D.; Pan, X.; et al. Identification of the Wheat C3H Gene Family and Expression Analysis of Candidates Associated with Seed Dormancy and Germination. Plant Physiol. Biochem. 2020, 156, 524–537. [Google Scholar] [CrossRef]
- Wang, Q.; Song, S.; Lu, X.; Wang, Y.; Chen, Y.; Wu, X.; Tan, L.; Chai, G. Hormone Regulation of CCCH Zinc Finger Proteins in Plants. Int. J. Mol. Sci. 2022, 23, 14288. [Google Scholar] [CrossRef]
- Li, L.; Zhang, C.; Huang, J.; Liu, Q.; Wei, H.; Wang, H.; Liu, G.; Gu, L.; Yu, S. Genomic Analyses Reveal the Genetic Basis of Early Maturity and Identification of Loci and Candidate Genes in Upland Cotton (Gossypium hirsutum L.). Plant Biotechnol. J. 2021, 19, 109–123. [Google Scholar] [CrossRef]
- Li, L.; Chang, H.; Zhao, S.; Liu, R.; Yan, M.; Li, F.; El-Sheery, N.I.; Feng, Z.; Yu, S. Combining High-Throughput Deep Learning Phenotyping and GWAS to Reveal Genetic Variants of Fruit Branch Angle in Upland Cotton. Ind. Crops Prod. 2024, 220, 119180. [Google Scholar] [CrossRef]
- Han, G.; Qiao, Z.; Li, Y.; Wang, C.; Wang, B. The Roles of CCCH Zinc-Finger Proteins in Plant Abiotic Stress Tolerance. Int. J. Mol. Sci. 2021, 22, 8327. [Google Scholar] [CrossRef]
- Seok, H.-Y.; Nguyen, L.V.; Park, H.-Y.; Tarte, V.N.; Ha, J.; Lee, S.-Y.; Moon, Y.-H. Arabidopsis Non-TZF Gene AtC3H17 Functions as a Positive Regulator in Salt Stress Response. Biochem. Biophys. Res. Commun. 2018, 498, 954–959. [Google Scholar] [CrossRef]
- Li, J.; Jia, D.; Chen, X. HUA1, a Regulator of Stamen and Carpel Identities in Arabidopsis, Codes for a Nuclear RNA Binding Protein. Plant Cell 2001, 13, 2269–2281. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Kato, N.; Wang, W.; Li, J.; Chen, X. Two RNA Binding Proteins, HEN4 and HUA1, Act in the Processing of AGAMOUS Pre-mRNA in Arabidopsis thaliana. Dev. Cell 2003, 4, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Chen, Y.; Yan, M.; Zhao, S.; Li, F.; Yu, S.; Feng, Z.; Li, L. Genome-Wide Identification and Mining Elite Allele Variation of the Monoacylglycerol Lipase (MAGL) Gene Family in Upland Cotton (Gossypium hirsutum L.). BMC Plant Biol. 2024, 24, 587. [Google Scholar] [CrossRef]
- Kim, D.H.; Yamaguchi, S.; Lim, S.; Oh, E.; Park, J.; Hanada, A.; Kamiya, Y.; Choi, G. SOMNUS, a CCCH-Type Zinc Finger Protein in Arabidopsis, Negatively Regulates Light-Dependent Seed Germination Downstream of PIL5. Plant Cell 2008, 20, 1260–1277. [Google Scholar] [CrossRef]
- Delaney, K.J.; Xu, R.; Zhang, J.; Li, Q.Q.; Yun, K.-Y.; Falcone, D.L.; Hunt, A.G. Calmodulin Interacts with and Regulates the RNA-Binding Activity of an Arabidopsis Polyadenylation Factor Subunit. Plant Physiol. 2006, 140, 1507–1521. [Google Scholar] [CrossRef]
- Guo, Y.-H.; Yu, Y.-P.; Wang, D.; Wu, C.-A.; Yang, G.-D.; Huang, J.-G.; Zheng, C.-C. GhZFP1, a Novel CCCH-Type Zinc Finger Protein from Cotton, Enhances Salt Stress Tolerance and Fungal Disease Resistance in Transgenic Tobacco by Interacting with GZIRD21A and GZIPR5. New Phytol. 2009, 183, 62–75. [Google Scholar] [CrossRef]
- Sun, J.; Jiang, H.; Xu, Y.; Li, H.; Wu, X.; Xie, Q.; Li, C. The CCCH-Type Zinc Finger Proteins AtSZF1 and AtSZF2 Regulate Salt Stress Responses in Arabidopsis. Plant Cell Physiol. 2007, 48, 1148–1158. [Google Scholar] [CrossRef]
- Lee, S.; Jung, H.J.; Kang, H.; Kim, S.Y. Arabidopsis Zinc Finger Proteins AtC3H49/AtTZF3 and AtC3H20/AtTZF2 Are Involved in ABA and JA Responses. Plant Cell Physiol. 2012, 53, 673–686. [Google Scholar] [CrossRef]
- Bogamuwa, S.; Jang, J.-C. The Arabidopsis Tandem CCCH Zinc Finger Proteins AtTZF4, 5 and 6 Are Involved in Light-, Abscisic Acid- and Gibberellic Acid-Mediated Regulation of Seed Germination. Plant Cell Environ. 2013, 36, 1507–1519. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A Comparative Platform for Green Plant Genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The Protein Families Database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent Updates, New Developments and Status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef] [PubMed]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s Conserved Domain Database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The Proteomics Server for in-Depth Protein Knowledge and Analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein Localization Predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y.; et al. TBtools-II: A “One for All, All for One” Bioinformatics Platform for Biological Big-Data Mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef]
- Zhang, Z. KaKs_Calculator 3.0: Calculating Selective Pressure on Coding and Non-Coding Sequences. Genom. Proteom. Bioinform. 2022, 20, 536–540. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhang, H.; Long, Y.; Shu, Y.; Zhai, J. Plant Public RNA-Seq Database: A Comprehensive Online Database for Expression Analysis of ~45 000 Plant Public RNA-Seq Libraries. Plant Biotechnol. J. 2022, 20, 806–808. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Lu, X.; Shen, L.; Guo, L.; Zhang, G.; Gao, Z.; Zhu, L.; Hu, J.; Dong, G.; Ren, D.; et al. Genome-Wide Association Study Reveals Novel QTLs and Candidate Genes for Seed Vigor in Rice. Front. Plant Sci. 2022, 13, 1005203. [Google Scholar] [CrossRef]
- Zhao, H.; Yao, W.; Ouyang, Y.; Yang, W.; Wang, G.; Lian, X.; Xing, Y.; Chen, L.; Xie, W. RiceVarMap: A Comprehensive Database of Rice Genomic Variations. Nucleic Acids Res. 2015, 43, D1018–D1022. [Google Scholar] [CrossRef]
- Wang, D.; Guo, Y.; Wu, C.; Yang, G.; Li, Y.; Zheng, C. Genome-Wide Analysis of CCCH Zinc Finger Family in Arabidopsis and Rice. BMC Genom. 2008, 9, 44. [Google Scholar] [CrossRef]
- Peng, X.; Zhao, Y.; Cao, J.; Zhang, W.; Jiang, H.; Li, X.; Ma, Q.; Zhu, S.; Cheng, B. CCCH-Type Zinc Finger Family in Maize: Genome-Wide Identification, Classification and Expression Profiling under Abscisic Acid and Drought Treatments. PLoS ONE 2012, 7, e40120. [Google Scholar] [CrossRef]
- Ai, Q.; Pan, W.; Zeng, Y.; Li, Y.; Cui, L. CCCH Zinc Finger Genes in Barley: Genome-Wide Identification, Evolution, Expression and Haplotype Analysis. BMC Plant Biol. 2022, 22, 117. [Google Scholar] [CrossRef]
- Hu, X.; Zuo, J. The CCCH Zinc Finger Family of Soybean (Glycine max L.): Genome-Wide Identification, Expression, Domestication, GWAS and Haplotype Analysis. BMC Genom. 2021, 22, 511. [Google Scholar] [CrossRef]
- Yuan, S.; Xu, B.; Zhang, J.; Xie, Z.; Cheng, Q.; Yang, Z.; Cai, Q.; Huang, B. Comprehensive Analysis of CCCH-Type Zinc Finger Family Genes Facilitates Functional Gene Discovery and Reflects Recent Allopolyploidization Event in Tetraploid Switchgrass. BMC Genom. 2015, 16, 129. [Google Scholar] [CrossRef]
- Pi, B.; Pan, J.; Xiao, M.; Hu, X.; Zhang, L.; Chen, M.; Liu, B.; Ruan, Y.; Huang, Y. Systematic Analysis of CCCH Zinc Finger Family in Brassica Napus Showed That BnRR-TZFs Are Involved in Stress Resistance. BMC Plant Biol. 2021, 21, 555. [Google Scholar] [CrossRef]
- Pi, B.; He, X.; Ruan, Y.; Jang, J.-C.; Huang, Y. Genome-Wide Analysis and Stress-Responsive Expression of CCCH Zinc Finger Family Genes in Brassica rapa. BMC Plant Biol. 2018, 18, 373. [Google Scholar] [CrossRef] [PubMed]
- Xu, R. Genome-Wide Analysis and Identification of Stress-Responsive Genes of the CCCH Zinc Finger Family in Solanum lycopersicum. Mol. Genet. Genom. 2014, 289, 965–979. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhong, Y.; Cheng, Z. (Max) Evolution and Expression Analysis of the CCCH Zinc Finger Gene Family in Vitis vinifera. Plant Genome 2014, 7, plantgenome2014.05.0019. [Google Scholar] [CrossRef]
- Liu, S.; Khan, M.R.G.; Li, Y.; Zhang, J.; Hu, C. Comprehensive Analysis of CCCH-Type Zinc Finger Gene Family in Citrus (Clementine mandarin) by Genome-Wide Characterization. Mol. Genet. Genom. 2014, 289, 855–872. [Google Scholar] [CrossRef]
- Chai, G.; Hu, R.; Zhang, D.; Qi, G.; Zuo, R.; Cao, Y.; Chen, P.; Kong, Y.; Zhou, G. Comprehensive Analysis of CCCH Zinc Finger Family in Poplar (Populus trichocarpa). BMC Genom. 2012, 13, 253. [Google Scholar] [CrossRef]
- Guo, C.; Chen, L.; Cui, Y.; Tang, M.; Guo, Y.; Yi, Y.; Li, Y.; Liu, L.; Chen, L. RNA Binding Protein OsTZF7 Traffics Between the Nucleus and Processing Bodies/Stress Granules and Positively Regulates Drought Stress in Rice. Front. Plant Sci. 2022, 13, 802337. [Google Scholar] [CrossRef]
- Seong, S.Y.; Shim, J.S.; Bang, S.W.; Kim, J.-K. Overexpression of OsC3H10, a CCCH-Zinc Finger, Improves Drought Tolerance in Rice by Regulating Stress-Related Genes. Plants 2020, 9, 1298. [Google Scholar] [CrossRef]
- Selvaraj, M.G.; Jan, A.; Ishizaki, T.; Valencia, M.; Dedicova, B.; Maruyama, K.; Ogata, T.; Todaka, D.; Yamaguchi-Shinozaki, K.; Nakashima, K.; et al. Expression of the CCCH-Tandem Zinc Finger Protein Gene OsTZF5 under a Stress-Inducible Promoter Mitigates the Effect of Drought Stress on Rice Grain Yield under Field Conditions. Plant Biotechnol. J. 2020, 18, 1711–1721. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Isoelectric Point | Molecular Weight | Protein Length | Subcellular Location |
|---|---|---|---|---|---|
| OsC3H1 | LOC_Os01g07930 | 8.69 | 29,860.95 | 279 | nucleus |
| OsC3H2 | LOC_Os01g09620 | 6.41 | 41,422.51 | 386 | nucleus |
| OsC3H3 | LOC_Os01g14870 | 8.88 | 17,888.01 | 167 | nucleus |
| OsC3H4 | LOC_Os01g15300 | 6.66 | 113,227.4 | 1007 | nucleus |
| OsC3H5 | LOC_Os01g15350 | 8.21 | 49,089.1 | 466 | nucleus |
| OsC3H6 | LOC_Os01g15460 | 7.50 | 49,834.79 | 476 | nucleus |
| OsC3H7 | LOC_Os01g39100 | 5.86 | 87,552.21 | 810 | nucleus |
| OsC3H8 | LOC_Os01g42970 | 6.90 | 50,358.15 | 461 | nucleus |
| OsC3H9 | LOC_Os01g45730 | 8.08 | 36,086.52 | 333 | nucleus |
| OsC3H10 | LOC_Os01g53650 | 6.06 | 24,982.17 | 225 | nucleus |
| OsC3H11 | LOC_Os01g61830 | 5.21 | 41,645.61 | 366 | nucleus |
| OsC3H12 | LOC_Os01g68860 | 8.89 | 47,852.24 | 439 | nucleus |
| OsC3H13 | LOC_Os02g06584 | 9.18 | 49,077.49 | 426 | nucleus |
| OsC3H14 | LOC_Os02g10080 | 9.58 | 31,153.44 | 300 | nucleus |
| OsC3H15 | LOC_Os02g19804 | 7.09 | 35,778.46 | 426 | nucleus |
| OsC3H16 | LOC_Os02g35150 | 8.85 | 78,052.14 | 678 | nucleus |
| OsC3H17 | LOC_Os02g45480 | 8.19 | 46,639.84 | 435 | chloroplast |
| OsC3H18 | LOC_Os02g55000 | 5.27 | 55,622.91 | 504 | nucleus |
| OsC3H19 | LOC_Os02g58440 | 5.29 | 71,469.18 | 647 | nucleus |
| OsC3H20 | LOC_Os03g02160 | 4.68 | 89,420.17 | 831 | nucleus |
| OsC3H21 | LOC_Os03g05210 | 6.47 | 46,317.29 | 427 | chloroplast |
| OsC3H22 | LOC_Os03g18950 | 5.88 | 48,549.22 | 457 | nucleus |
| OsC3H23 | LOC_Os03g21140 | 6.00 | 73,135.88 | 688 | nucleus |
| OsC3H24 | LOC_Os03g21160 | 6.07 | 71,950.88 | 677 | nucleus |
| OsC3H25 | LOC_Os03g49170 | 6.63 | 81,043.54 | 764 | endoplasmic reticulum |
| OsC3H26 | LOC_Os03g61110 | 9.25 | 36,258.57 | 312 | nucleus |
| OsC3H27 | LOC_Os04g01480 | 5.2 | 92,340.68 | 850 | nucleus |
| OsC3H28 | LOC_Os04g02730 | 6.25 | 75,572.46 | 685 | nucleus |
| OsC3H29 | LOC_Os04g32340 | 8.70 | 97,243.83 | 903 | nucleus |
| OsC3H30 | LOC_Os04g35800 | 9.61 | 38,697.91 | 346 | nucleus |
| OsC3H31 | LOC_Os04g41060 | 9.41 | 15,845.9 | 146 | extracellular |
| OsC3H32 | LOC_Os04g48375 | 8.38 | 34,286.86 | 315 | cytosol |
| OsC3H33 | LOC_Os04g56750 | 6.27 | 50,472.86 | 469 | chloroplast |
| OsC3H34 | LOC_Os04g57010 | 9.48 | 31,787.8 | 309 | cytosol |
| OsC3H35 | LOC_Os04g57600 | 5.33 | 80,159.97 | 711 | nucleus |
| OsC3H36 | LOC_Os05g03760 | 8.65 | 63,235.89 | 601 | chloroplast |
| OsC3H37 | LOC_Os05g08400 | 6.04 | 55,632.5 | 493 | nucleus |
| OsC3H38 | LOC_Os05g10670 | 9.02 | 49,684.7 | 464 | chloroplast |
| OsC3H39 | LOC_Os05g41790 | 9.25 | 76,541.3 | 703 | nucleus |
| OsC3H40 | LOC_Os05g45020 | 5.39 | 28,257.88 | 255 | nucleus |
| OsC3H41 | LOC_Os05g48960 | 9.25 | 34,692.17 | 304 | nucleus |
| OsC3H42 | LOC_Os05g50080 | 8.65 | 37,169.12 | 343 | cytosol |
| OsC3H43 | LOC_Os06g07350 | 8.12 | 54,009.89 | 482 | chloroplast |
| OsC3H44 | LOC_Os06g21390 | 7.96 | 41,673.18 | 368 | nucleus |
| OsC3H45 | LOC_Os06g32720 | 6.74 | 31,058.7 | 279 | nucleus |
| OsC3H46 | LOC_Os06g32860 | 6.01 | 80,907.75 | 711 | nucleus |
| OsC3H47 | LOC_Os06g41384 | 9.53 | 30,451.7 | 295 | nucleus |
| OsC3H48 | LOC_Os06g41390 | 6.85 | 64,563.78 | 600 | nucleus |
| OsC3H49 | LOC_Os06g43120 | 7.97 | 207,156.46 | 1890 | nucleus |
| OsC3H50 | LOC_Os06g46400 | 6.32 | 72,597.53 | 665 | nucleus |
| OsC3H51 | LOC_Os06g46890 | 9.24 | 42,478.83 | 372 | nucleus |
| OsC3H52 | LOC_Os06g49080 | 8.94 | 42,663.15 | 390 | nucleus |
| OsC3H53 | LOC_Os07g04580 | 6.72 | 40,200.85 | 388 | nucleus |
| OsC3H54 | LOC_Os07g04650 | 4.78 | 51,701.7 | 496 | nucleus |
| OsC3H55 | LOC_Os07g18050 | 6.72 | 54,334.99 | 486 | cytosol |
| OsC3H56 | LOC_Os07g38090 | 6.61 | 69,391.93 | 657 | endoplasmic reticulum |
| OsC3H57 | LOC_Os07g39440 | 9.07 | 49,917.89 | 438 | nucleus |
| OsC3H58 | LOC_Os07g47240 | 8.04 | 31,590.77 | 280 | nucleus |
| OsC3H59 | LOC_Os07g48410 | 6.15 | 72,494.47 | 682 | nucleus |
| OsC3H60 | LOC_Os08g03310 | 6.73 | 71,817.92 | 653 | cytosol |
| OsC3H61 | LOC_Os08g04170 | 8.76 | 105,583.58 | 958 | nucleus |
| OsC3H62 | LOC_Os08g06330 | 6.82 | 37,957.8 | 367 | nucleus |
| OsC3H63 | LOC_Os08g38370 | 5.69 | 68,736.17 | 605 | nucleus |
| OsC3H64 | LOC_Os09g13530 | 7.05 | 26,161.09 | 252 | nucleus |
| OsC3H65 | LOC_Os09g19940 | 8.11 | 57,894.38 | 527 | chloroplast |
| OsC3H66 | LOC_Os09g31482 | 8.92 | 34,087.52 | 290 | nucleus |
| OsC3H67 | LOC_Os09g36090 | 4.70 | 198,678.84 | 1835 | nucleus |
| OsC3H68 | LOC_Os10g25220 | 8.78 | 81,200.6 | 702 | nucleus |
| OsC3H69 | LOC_Os11g28270 | 8.77 | 48,006.1 | 444 | nucleus |
| OsC3H70 | LOC_Os12g03554 | 6.38 | 60,054.42 | 527 | nucleus |
| OsC3H71 | LOC_Os12g18120 | 6.14 | 57,306.11 | 529 | nucleus |
| OsC3H72 | LOC_Os12g21700 | 8.50 | 48,663.52 | 454 | nucleus |
| OsC3H73 | LOC_Os12g33090 | 6.04 | 64,725.96 | 619 | plasma membrane |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Li, S.; Wu, H.; Huang, L.; Fu, L.; Zhan, C.; Lu, X.; Yang, L.; Dai, L.; Zeng, D. Identification and Expression Analysis of CCCH Zinc Finger Family Genes in Oryza sativa. Genes 2025, 16, 429. https://doi.org/10.3390/genes16040429
Wang Z, Li S, Wu H, Huang L, Fu L, Zhan C, Lu X, Yang L, Dai L, Zeng D. Identification and Expression Analysis of CCCH Zinc Finger Family Genes in Oryza sativa. Genes. 2025; 16(4):429. https://doi.org/10.3390/genes16040429
Chicago/Turabian StyleWang, Zhihan, Shunyuan Li, Hongkai Wu, Linzhou Huang, Liangbo Fu, Chengfang Zhan, Xueli Lu, Long Yang, Liping Dai, and Dali Zeng. 2025. "Identification and Expression Analysis of CCCH Zinc Finger Family Genes in Oryza sativa" Genes 16, no. 4: 429. https://doi.org/10.3390/genes16040429
APA StyleWang, Z., Li, S., Wu, H., Huang, L., Fu, L., Zhan, C., Lu, X., Yang, L., Dai, L., & Zeng, D. (2025). Identification and Expression Analysis of CCCH Zinc Finger Family Genes in Oryza sativa. Genes, 16(4), 429. https://doi.org/10.3390/genes16040429