
Aberrant Expression of Non-Coding RNAs in Pediatric T Acute Lymphoblastic Leukemia and Their Potential Application as Biomarkers

 ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
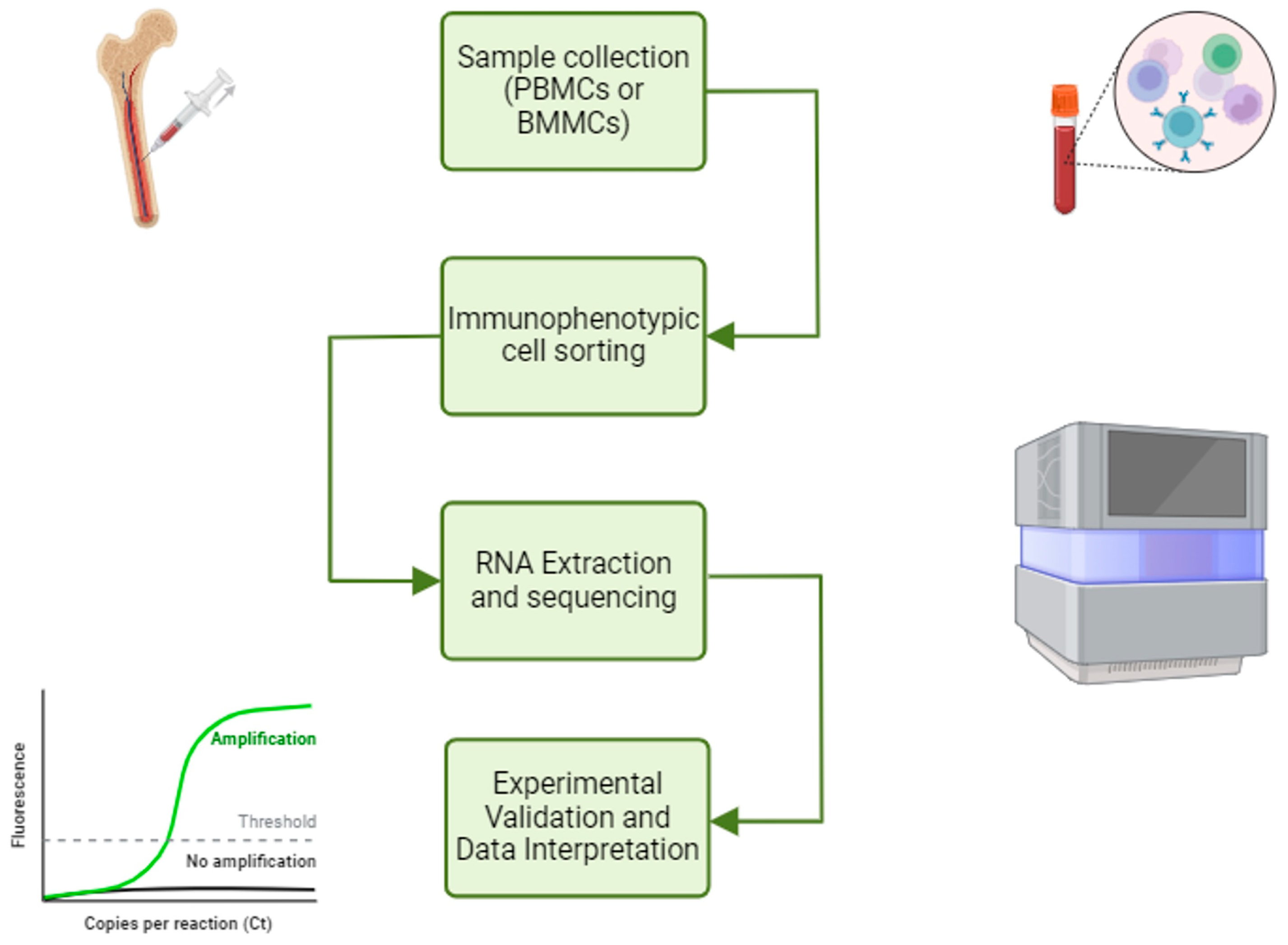
2. Analytic Workflow to Enlighten the Roles of ncRNA in T-ALL
3. ncRNAs as Potential Therapeutic Target in Pediatric T-ALL Patients
3.1. Tumor-Suppressor ncRNAs
3.2. Oncogenic ncRNAs
4. ncRNAs as Potential Diagnostic Tool in Pediatric T-ALL Patients
ncRNAs and Central Nervous System (CNS) Invasion in the Management of Pediatric T-ALL
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ADR | Adriamycin |
| BMMC | Mononuclear cells from bone marrow |
| CNS | Central nervous system |
| LncRNA | Long non-coding RNA |
| miRNA | microRNA |
| ncRNA | Non-coding RNA |
| NGS | Next-generation sequencing |
| OGM | Optical genome mapping |
| PBMC | Mononuclear cells from peripheral blood |
| RNA-seq | RNA-sequencing |
| T-ALL | T acute lymphoblastic leukemia |
References
- Lato, M.W.; Przysucha, A.; Grosman, S.; Zawitkowska, J.; Lejman, M. The New Therapeutic Strategies in Pediatric T-Cell Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2021, 22, 4502. [Google Scholar] [CrossRef] [PubMed]
- Ekpa, Q.L.; Akahara, P.C.; Anderson, A.M.; Adekoya, O.O.; Ajayi, O.O.; Alabi, P.O.; Okobi, O.E.; Jaiyeola, O.; Ekanem, M.S. A Review of Acute Lymphocytic Leukemia (ALL) in the Pediatric Population: Evaluating Current Trends and Changes in Guidelines in the Past Decade. Cureus 2023, 15, e49930. [Google Scholar]
- Inaba, H.; Pui, C.H. Advances in the Diagnosis and Treatment of Pediatric Acute Lymphoblastic Leukemia. J. Clin. Med. 2021, 10, 1926. [Google Scholar] [CrossRef] [PubMed]
- Slack, F.J.; Chinnaiyan, A.M. The Role of Non-coding RNAs in Oncology. Cell 2019, 179, 1033–1055. [Google Scholar]
- Toden, S.; Zumwalt, T.J.; Goel, A. Non-coding RNAs and potential therapeutic targeting in cancer. Biochim. Biophys. Acta Rev. Cancer 2021, 1875, 188491. [Google Scholar]
- Dakal, T.C.; Philip, R.R.; Bhushan, R.; Sonar, P.V.; Rajagopal, S.; Kumar, A. Genetic and epigenetic regulation of non-coding RNAs: Implications in cancer metastasis, stemness and drug resistance. Pathol. Res. Pract. 2025, 266, 155728. [Google Scholar] [PubMed]
- Ghazimoradi, M.H.; Karimpour-Fard, N.; Babashah, S. The Promising Role of Non-Coding RNAs as Biomarkers and Therapeutic Targets for Leukemia. Genes 2023, 14, 131. [Google Scholar] [CrossRef]
- Orlandella, F.M.; Mariniello, R.M.; Mirabelli, P.; De Stefano, A.E.; Iervolino, P.L.C.; Lasorsa, V.A.; Capasso, M.; Giannatiempo, R.; Rongo, M.; Incoronato, M.; et al. miR-622 is a novel potential biomarker of breast carcinoma and impairs motility of breast cancer cells through targeting NUAK1 kinase. Br. J. Cancer 2020, 123, 426–437. [Google Scholar]
- Orlandella, F.M.; Imperlini, E.; Pane, K.; Luciano, N.; Braile, M.; De Stefano, A.E.; Iervolino, P.L.C.; Ruocco, A.; Orrù, S.; Franzese, M.; et al. miR-331-5p Affects Motility of Thyroid Cancer Cell Lines and Regulates BID Expression. Biomedicines 2024, 12, 658. [Google Scholar] [CrossRef]
- Braile, M.; Luciano, N.; Carlomagno, D.; Salvatore, G.; Orlandella, F.M. Insight into the Role of the miR-584 Family in Human Cancers. Int. J. Mol. Sci. 2024, 25, 7448. [Google Scholar] [CrossRef]
- Hussen, B.M.; Hidayat, H.J.; Salihi, A.; Sabir, D.K.; Taheri, M.; Ghafouri-Fard, S. MicroRNA: A signature for cancer progression. Biomed. Pharmacother. 2021, 138, 111528. [Google Scholar]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell Physiol. 2019, 234, 5451–5465. [Google Scholar]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [PubMed]
- Hu, G.; Niu, F.; Humburg, B.A.; Liao, K.; Bendi, S.; Callen, S.; Fox, H.S.; Buch, S. Molecular mechanisms of long noncoding RNAs and their role in disease pathogenesis. Oncotarget 2018, 9, 18648–18663. [Google Scholar]
- Sharma, S. Unraveling the role of long non-coding RNAs in therapeutic resistance in acute myeloid leukemia: New prospects & challenges. Noncoding RNA Res. 2024, 9, 1203–1221. [Google Scholar] [PubMed]
- Orlandella, F.M.; Smaldone, G.; Salvatore, G.; Vitagliano, L.; Cianflone, A.; Parasole, R.; Beneduce, G.; Menna, G.; Salvatore, M.; Mirabelli, P. The lncRNA TEX41 is upregulated in pediatric B-Cells Acute Lymphoblastic Leukemia and it is necessary for leukemic cell growth. Biomark Res. 2021, 9, 54. [Google Scholar]
- Affinito, O.; Pane, K.; Smaldone, G.; Orlandella, F.M.; Mirabelli, P.; Beneduce, G.; Parasole, R.; Ripaldi, M.; Salvatore, M.; Franzese, M. lncRNAs-mRNAs Co-Expression Network Underlying Childhood B-Cell Acute Lymphoblastic Leukaemia: A Pilot Study. Cancers 2020, 12, 2489. [Google Scholar] [CrossRef]
- Dremsek, P.; Schwarz, T.; Weil, B.; Malashka, A.; Laccone, F.; Neesen, J. Optical Genome Mapping in Routine Human Genetic Diagnostics-Its Advantages and Limitations. Genes 2021, 12, 1958. [Google Scholar] [CrossRef]
- Dawidowska, M.; Maćkowska-Maślak, N.; Drobna-Śledzińska, M.; Kosmalska, M.; Jaksik, R.; Szymczak, D.; Jarmuż-Szymczak, M.; Sadowska-Klasa, A.; Wojtaszewska, M.; Sędek, Ł.; et al. Small RNA-Seq Reveals Similar miRNA Transcriptome in Children and Young Adults with T-ALL and Indicates miR-143-3p as Novel Candidate Tumor Suppressor in This Leukemia. Int. J. Mol. Sci. 2022, 23, 10117. [Google Scholar] [CrossRef]
- Mets, E.; Van der Meulen, J.; Van Peer, G.; Boice, M.; Mestdagh, P.; Van de Walle, I.; Lammens, T.; Goossens, S.; De Moerloose, B.; Benoit, Y.; et al. MicroRNA-193b-3p acts as a tumor suppressor by targeting the MYB oncogene in T-cell acute lymphoblastic leukemia. Leukemia 2015, 29, 798–806. [Google Scholar] [PubMed]
- Feng, H.; Li, F.; Tang, P. Circ_0000745 regulates NOTCH1-mediated cell proliferation and apoptosis in pediatric T-cell acute lymphoblastic leukemia through adsorbing miR-193b-3p. Hematology 2021, 26, 885–895. [Google Scholar]
- Guo, Y.; Zhu, X.; Sun, X. COTI-2 induces cell apoptosis in pediatric acute lymphoblastic leukemia via upregulation of miR-203. Bioengineered 2020, 11, 201–208. [Google Scholar]
- Renou, L.; Boelle, P.Y.; Deswarte, C.; Spicuglia, S.; Benyoucef, A.; Calvo, J.; Uzan, B.; Belhocine, M.; Cieslak, A.; Landman-Parker, J.; et al. Homeobox protein TLX3 activates miR-125b expression to promote T-cell acute lymphoblastic leukemia. Blood Adv. 2017, 1, 733–747. [Google Scholar]
- Hou, Y.; Sun, J.; Huang, J.; Yao, F.; Chen, X.; Zhu, B.; Zhao, D. Circular RNA circRNA_0000094 sponges microRNA-223-3p and up-regulate F-box and WD repeat domain containing 7 to restrain T cell acute lymphoblastic leukemia progression. Human Cell 2021, 34, 977–989. [Google Scholar] [PubMed]
- Mansour, M.R.; Sanda, T.; Lawton, L.N.; Li, X.; Kreslavsky, T.; Novina, C.D.; Brand, M.; Gutierrez, A.; Kelliher, M.A.; Jamieson, C.H.; et al. The TAL1 complex targets the FBXW7 tumor suppressor by activating miR-223 in human T cell acute lymphoblastic leukemia. J. Exp. Med. 2013, 210, 1545–1557. [Google Scholar]
- Shu, Y.; Wang, Y.; Lv, W.Q.; Peng, D.Y.; Li, J.; Zhang, H.; Jiang, G.J.; Yang, B.J.; Liu, S.; Zhang, J.; et al. ARRB1-Promoted NOTCH1 Degradation Is Suppressed by OncomiR miR-223 in T-cell Acute Lymphoblastic Leukemia. Cancer Res. 2020, 80, 988–998. [Google Scholar] [PubMed]
- Drobna, M.; Szarzyńska, B.; Jaksik, R.; Sędek, Ł.; Kuchmiy, A.; Taghon, T.; Van Vlierberghe, P.; Szczepański, T.; Witt, M.; Dawidowska, M. hsa-miR-20b-5p and hsa-miR-363-3p Affect Expression of PTEN and BIM Tumor Suppressor Genes and Modulate Survival of T-ALL Cells In Vitro. Cells 2020, 9, 1137. [Google Scholar] [CrossRef]
- Drobna-Śledzińska, M.; Maćkowska-Maślak, N.; Jaksik, R.; Kosmalska, M.; Szarzyńska, B.; Lejman, M.; Sędek, Ł.; Szczepański, T.; Taghon, T.; Van Vlierberghe, P.; et al. Multiomics to investigate the mechanisms contributing to repression of PTPRC and SOCS2 in pediatric T-ALL: Focus on miR-363-3p and promoter methylation. Genes Chromosomes Cancer 2022, 61, 720–733. [Google Scholar]
- Liu, X.; Zhang, H.; Zhang, B.; Zhang, X. Expression and Role of MicroRNA-663b in Childhood Acute Lymphocytic Leukemia and its Mechanism. Open Med. 2019, 14, 863–871. [Google Scholar]
- Liu, S.; Wang, H.; Guo, W.; Zhou, X.; Shu, Y.; Liu, H.; Yang, L.; Tang, S.; Su, H.; Liu, Z.; et al. MiR-652-5p elevated glycolysis level by targeting TIGAR in T-cell acute lymphoblastic leukemia. Cell Death Dis. 2022, 13, 148. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, P.; Lin, R.; Rong, L.; Xue, Y.; Fang, Y. LncRNA NALT interaction with NOTCH1 promoted cell proliferation in pediatric T cell acute lymphoblastic leukemia. Sci. Rep. 2015, 5, 13749. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xu, H.G.; Lu, C. A novel long non-coding RNA T-ALL-R-LncR1 knockdown and Par-4 cooperate to induce cellular apoptosis in T-cell acute lymphoblastic leukemia cells. Leuk. Lymphoma 2014, 55, 1373–1382. [Google Scholar] [CrossRef]
- Chen, L.; Shi, Y.; Li, J.; Yang, X.; Li, R.; Zhou, X.; Zhu, L. LncRNA CDKN2B-AS1 contributes to tumorigenesis and chemoresistance in pediatric T-cell acute lymphoblastic leukemia through miR-335-3p/TRAF5 axis. Anticancer Drugs 2020. [Google Scholar] [CrossRef]
- Schotte, D.; e Menezes, R.X.; Akbari Moqadam, F.; Khankahdani, L.M.; Lange-Turenhout, E.; Chen, C.; Pieters, R.; Den Boer, M.L. MicroRNA characterize genetic diversity and drug resistance in pediatric acute lymphoblastic leukemia. Haematologica 2011, 96, 703–711. [Google Scholar] [CrossRef]
- Schotte, D.; Lange-Turenhout, E.A.; Stumpel, D.J.; Stam, R.W.; Buijs-Gladdines, J.G.; Meijerink, J.P.; Pieters, R.; Den Boer, M.L. Expression of miR-196b is not exclusively MLL-driven but is especially linked to activation of HOXA genes in pediatric acute lymphoblastic leukemia. Haematologica 2010, 95, 1675–1682. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.S.; Chang, W.S.; Chen, C.C.; Mong, M.C.; Hsu, S.W.; Hsu, P.C.; Hsu, Y.N.; Wang, Y.C.; Tsai, C.W.; Bau, D.T. Novel Contribution of Long Non-coding RNA MEG3 Genotype to Prediction of Childhood Leukemia Risk. Cancer Genom. Proteom. 2022, 19, 27–34. [Google Scholar]
- Mofidi, M.; Rahgozar, S.; Pouyanrad, S. Increased level of long non coding RNA H19 is correlated with the downregulation of miR-326 and BCL-2 genes in pediatric acute lymphoblastic leukemia, a possible hallmark for leukemogenesis. Mol. Biol. Rep. 2021, 48, 1531–1538. [Google Scholar] [CrossRef]
- El-Khazragy, N.; Abdel Aziz, M.A.; Hesham, M.; Matbouly, S.; Mostafa, S.A.; Bakkar, A.; Abouelnile, M.; Noufal, Y.; Mahran, N.A.; Abd Elkhalek, M.A.; et al. Upregulation of leukemia-induced non-coding activator RNA (LUNAR1) predicts poor outcome in pediatric T-acute lymphoblastic leukemia. Immunobiology 2021, 226, 152149. [Google Scholar] [CrossRef]
- Sharma PKaur, P.; Bhatia, P.; Trehan, A.; Sreedharanunni, S.; Singh, M. Novel lncRNAs LINC01221, RP11-472G21.2 and CRNDE are markers of differential expression in pediatric patients with T cell acute lymphoblastic leukemia. Cancer Cell Int. 2024, 24, 65. [Google Scholar] [CrossRef]
- das Chagas, P.F.; de Sousa, G.R.; Kodama, M.H.; de Biagi Junior, C.A.O.; Yunes, J.A.; Brandalise, S.R.; Calin, G.A.; Tone, L.G.; Scrideli, C.A.; de Oliveira, J.C. Ultraconserved long non-coding RNA uc.112 is highly expressed in childhood T versus B-cell acute lymphoblastic leukemia. Hematol. Transfus. Cell Ther. 2021, 43, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Asadi, M.; Gholampour, M.A.; Kompani, F.; Alizadeh, S. Expression of Long Non-Coding RNA H19 in Acute Lymphoblastic Leukemia. Cell J. 2023, 25, 1–10. [Google Scholar] [PubMed]
- Thastrup, M.; Duguid, A.; Mirian, C.; Schmiegelow, K.; Halsey, C. Central nervous system involvement in childhood acute lymphoblastic leukemia: Challenges and solutions. Leukemia 2022, 36, 2751–2768. [Google Scholar] [CrossRef]
- Egyed, B.; Kutszegi, N.; Sági, J.C.; Gézsi, A.; Rzepiel, A.; Visnovitz, T.; Lőrincz, P.; Müller, J.; Zombori, M.; Szalai, C.; et al. MicroRNA-181a as novel liquid biopsy marker of central nervous system involvement in pediatric acute lymphoblastic leukemia. J. Transl. Med. 2020, 18, 250. [Google Scholar] [CrossRef] [PubMed]
- Li, X.J.; Ren, Z.J.; Tang, J.H. MicroRNA-34a: A potential therapeutic target in human cancer. Cell Death Dis. 2014, 5, e1327. [Google Scholar]
- Buono, L.; Iside, C.; De Matteo, A.; Stellato, P.; Beneduce, G.; de Vera d’Aragona, R.P.; Parasole, R.; Salvatore, M.; Smaldone, G.; Mirabelli, P. Specific lncRNA signatures discriminate childhood acute leukaemias: A pilot study. Cancer Cell Int. 2022, 22, 373. [Google Scholar]


{kind=link}
{kind=link}
| miRNAs | Regulator | Target Genes | Function | Experimental Model | Ref. |
|---|---|---|---|---|---|
| miR-143-3p | ns | KRAS, FGF1 and FGF9 | Inhibits T-ALL proliferation | In vitro | [20] |
| miR-193b-3p | ns | MYB | Inhibits T-ALL onset | In vitro and in vivo | [21] |
| circ_0000745 | NOTCH1 | Inhibits T-ALL proliferation | In vitro | [22] | |
| miR-203 | COTI-2 treatment | ns | Promotes apoptosis | In vitro and in vivo | [23] |
| miRNAs | Regulator | Target Genes | Function | Experimental Model | Ref. |
|---|---|---|---|---|---|
| miR-125b | TLX3/ LINC00478 | Ets1, CBFb | Impairs T-cell differentiation | In vitro and in vivo | [24] |
| miR-223-3p | circ_0000094 | FBW7 | Promotes T-ALL proliferation, migration and invasion, reduces apoptosis | In vitro | [25] |
| TAL1 | FBW7 | Promotes T-ALL proliferation, reduces apoptosis | In vitro | [26] | |
| ns | ARRB1 | Promotes T-ALL progression | In vitro and in vivo | [27] | |
| miR-20b-5p/ miR-363-3p | ns | PTEN, BIM | Enhance T-ALL proliferation, inhibit apoptosis | In vitro | [28] |
| miR-363-3p | Promoter hypermethylation | PTPRC and SOCS2 | Enhances cell growth by inhibiting JAK–STAT pathway | In vitro | [29] |
| miR-663b | ns | CD99 | Enhances proliferation and motility, inhibits apoptosis | In vitro | [30] |
| miR-652-5p | ns | TIGAR | Reduces glucose metabolism, slowing leukemia progression | In vitro and in vivo | [31] |
| lncRNAs | Regulator | Target Genes | Function | Experimental Model | Ref. |
|---|---|---|---|---|---|
| NALT | ns | NOTCH1 | Cell proliferation | In vivo and in vitro | [32] |
| T-ALL-R-LncR1 | ns | PAR-4 | Apoptosis | In vitro | [33] |
| CDKN2B-AS1 | ns | miR-335-3p | Drug resistance | In vivo and in vitro | [34] |
| miRNAs | Expression | Pediatric Cohort | Methods | Ref. |
|---|---|---|---|---|
| miR-363-3p | Up | 34 T-ALL vs. 5 healthy children | q-RT-PCR | [28] |
| 81 ALL vs. 17 healthy children | microRNA array | [35] | ||
| miR-663b | Up | 30 T-ALL vs. 30 healthy donors | q-RT-PCR | [30] |
| miR-196b | Up | 22 T-ALL vs. 50 B-ALL | q-RT-PCR | [36] |
| miR-652-5p | Up | 13 T-ALL vs. 5 healthy children | q-RT-PCR | [31] |
| lncRNAs | Expression | Pediatric Cohort | Methods | Ref. |
|---|---|---|---|---|
| MEG3 rs7158663 AG/AA | Higher susceptibility | 266 ALL vs. matched healthy controls | q-RT-PCR | [37] |
| H19 | Up | 37 ALL vs. 17 healthy controls | In silico and q-RT-PCR | [38] |
| NALT | Up | 20 T-ALL vs. 10 healthy controls | q-RT-PCR | [32] |
| CDKN2B-AS1 | Up | 42 T-ALL vs.20 healthy controls 21 T-ALL ADR resistance vs. 21 T-ALL non-ADR | q-RT-PCR | [34] |
| T-ALL-R-LncR1 | Up | 21 T-ALL | q-RT-PCR | [33] |
| LUNAR1 | Up | 185 T-ALL vs. 40 healthy controls | q-RT-PCR | [39] |
| LINC01221, RP11-472G21.2 CRNDE | Up | 51 T-ALL vs. 32 B-ALL vs.16 healthy controls | RNA sequencing | [40] |
| lncRNA uc.112, | Up | 32 T-ALL vs.30 B-ALL | q-RT-PCR | [41] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luciano, N.; Coppola, L.; Salvatore, G.; Primo, P.; Parasole, R.; Mirabelli, P.; Orlandella, F.M. Aberrant Expression of Non-Coding RNAs in Pediatric T Acute Lymphoblastic Leukemia and Their Potential Application as Biomarkers. Genes 2025, 16, 420. https://doi.org/10.3390/genes16040420
Luciano N, Coppola L, Salvatore G, Primo P, Parasole R, Mirabelli P, Orlandella FM. Aberrant Expression of Non-Coding RNAs in Pediatric T Acute Lymphoblastic Leukemia and Their Potential Application as Biomarkers. Genes. 2025; 16(4):420. https://doi.org/10.3390/genes16040420
Chicago/Turabian StyleLuciano, Neila, Luigi Coppola, Giuliana Salvatore, Pasquale Primo, Rosanna Parasole, Peppino Mirabelli, and Francesca Maria Orlandella. 2025. "Aberrant Expression of Non-Coding RNAs in Pediatric T Acute Lymphoblastic Leukemia and Their Potential Application as Biomarkers" Genes 16, no. 4: 420. https://doi.org/10.3390/genes16040420
APA StyleLuciano, N., Coppola, L., Salvatore, G., Primo, P., Parasole, R., Mirabelli, P., & Orlandella, F. M. (2025). Aberrant Expression of Non-Coding RNAs in Pediatric T Acute Lymphoblastic Leukemia and Their Potential Application as Biomarkers. Genes, 16(4), 420. https://doi.org/10.3390/genes16040420