
Comparative Analysis of Salt Tolerance and Transcriptomics in Two Varieties of Agropyron desertorum at Different Developmental Stages

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatment Conditions
2.1.1. Seed Stage
2.1.2. Seedling Stage
2.1.3. Plant Treatments for Transcriptome Sequencing
2.2. RNA Extraction and Sequencing
2.3. Quality Control, Transcriptome Assembly, and Annotation of Unigenes
2.4. Differential Gene Expression Analysis
3. Results
3.1. Salt Treatment Performance at Seed Stage
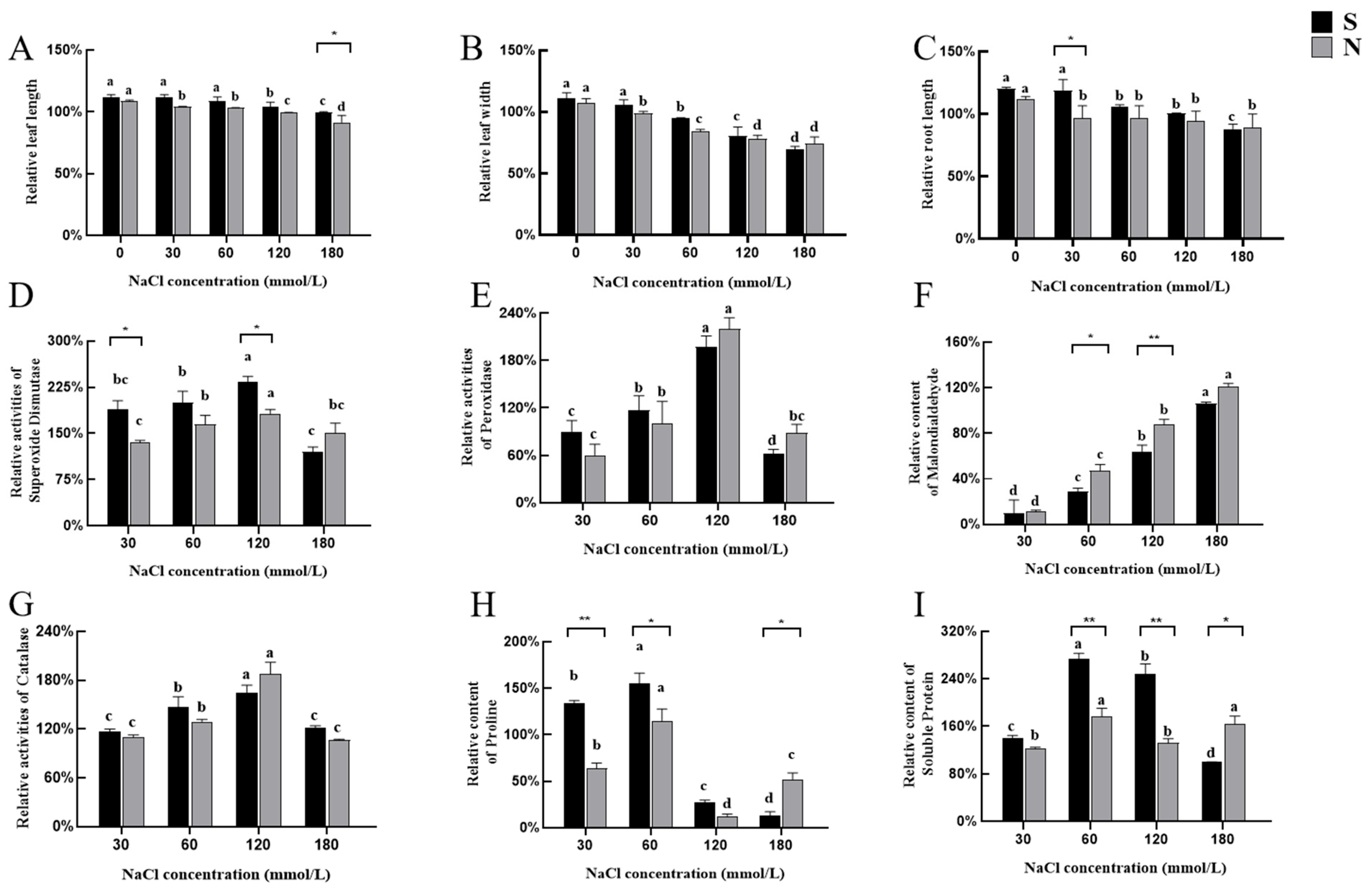
3.2. Morphological and Physiological Responses of Seedlings Under NaCl Stress
3.2.1. Morphological Responses of Seedlings to NaCl Stress
3.2.2. Physiological Responses of Seedlings to NaCl Stress
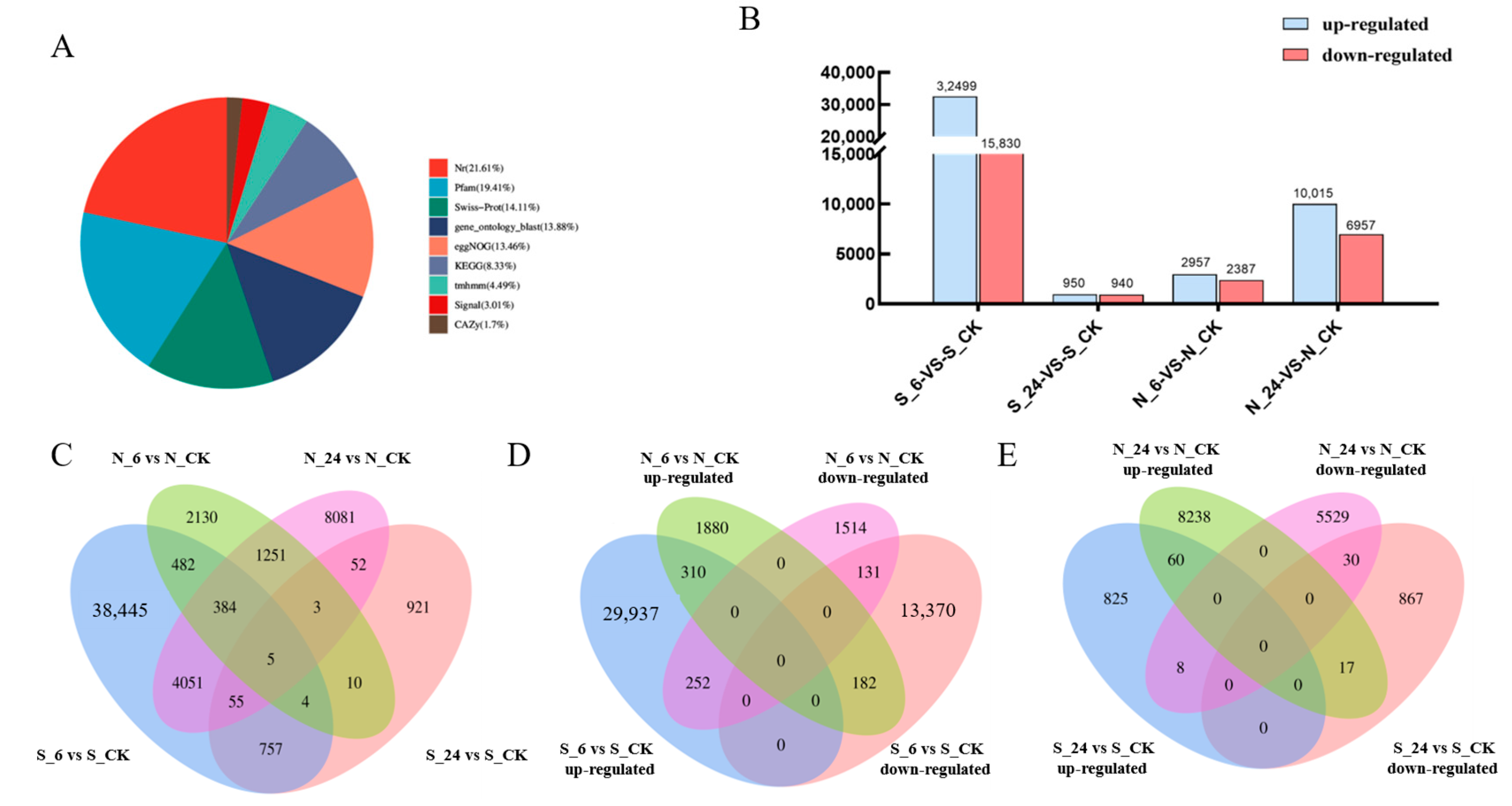
3.3. A. desertorum Genes Assembled from Transcriptome Data
3.3.1. Functional Annotation of All Non-Redundant Unigenes
3.3.2. Overall Gene Expression Patterns of Two Varieties Under NaCl Stress
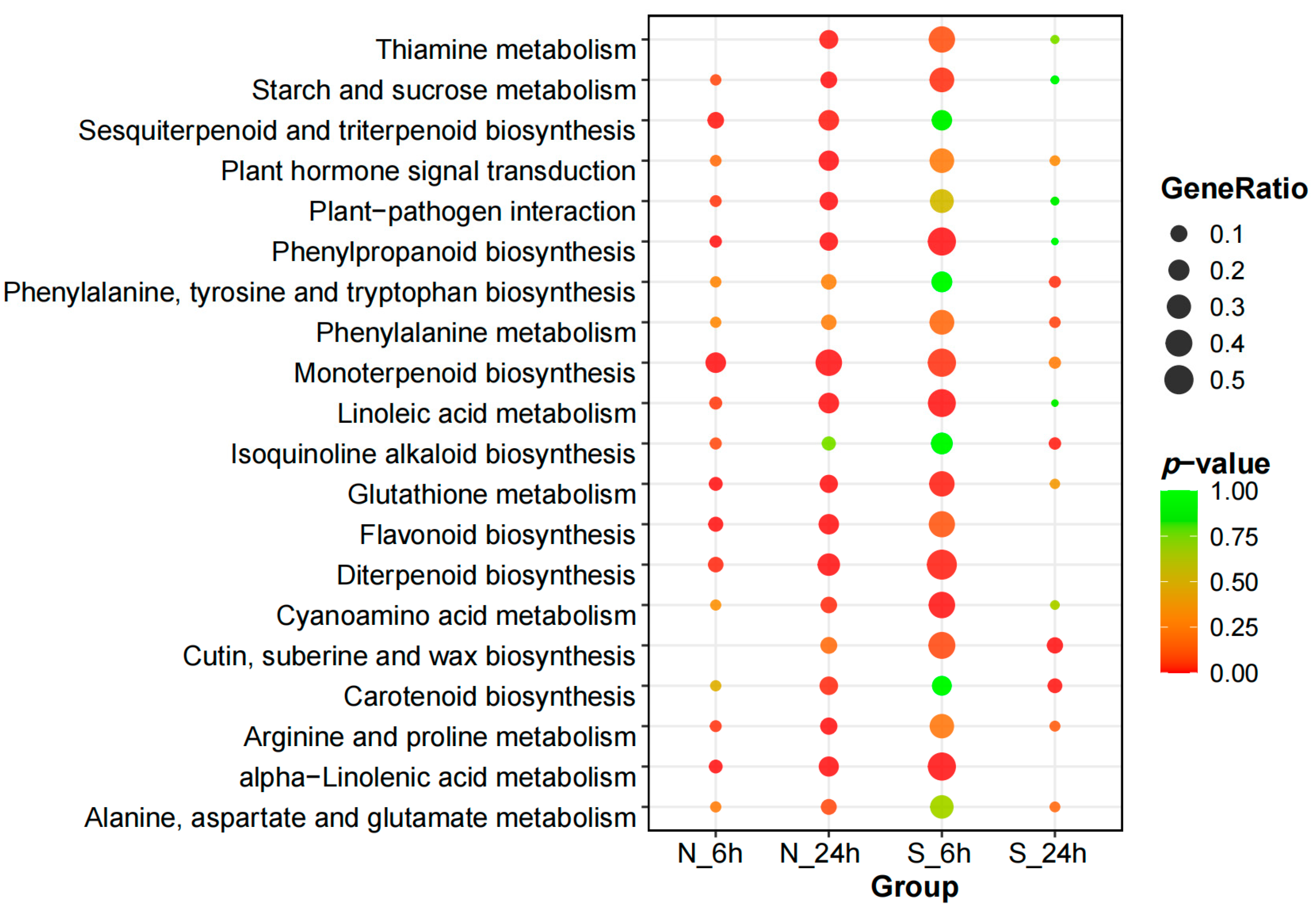
3.4. Functional Classification of DEGs According to GO and KEGG Pathways Under Salt Stress
3.4.1. Gene Ontology Enrichment Analysis of DEGs
3.4.2. Kyoto Encyclopedia of Genes and Genomes Pathway Enrichment Analysis of DEGs
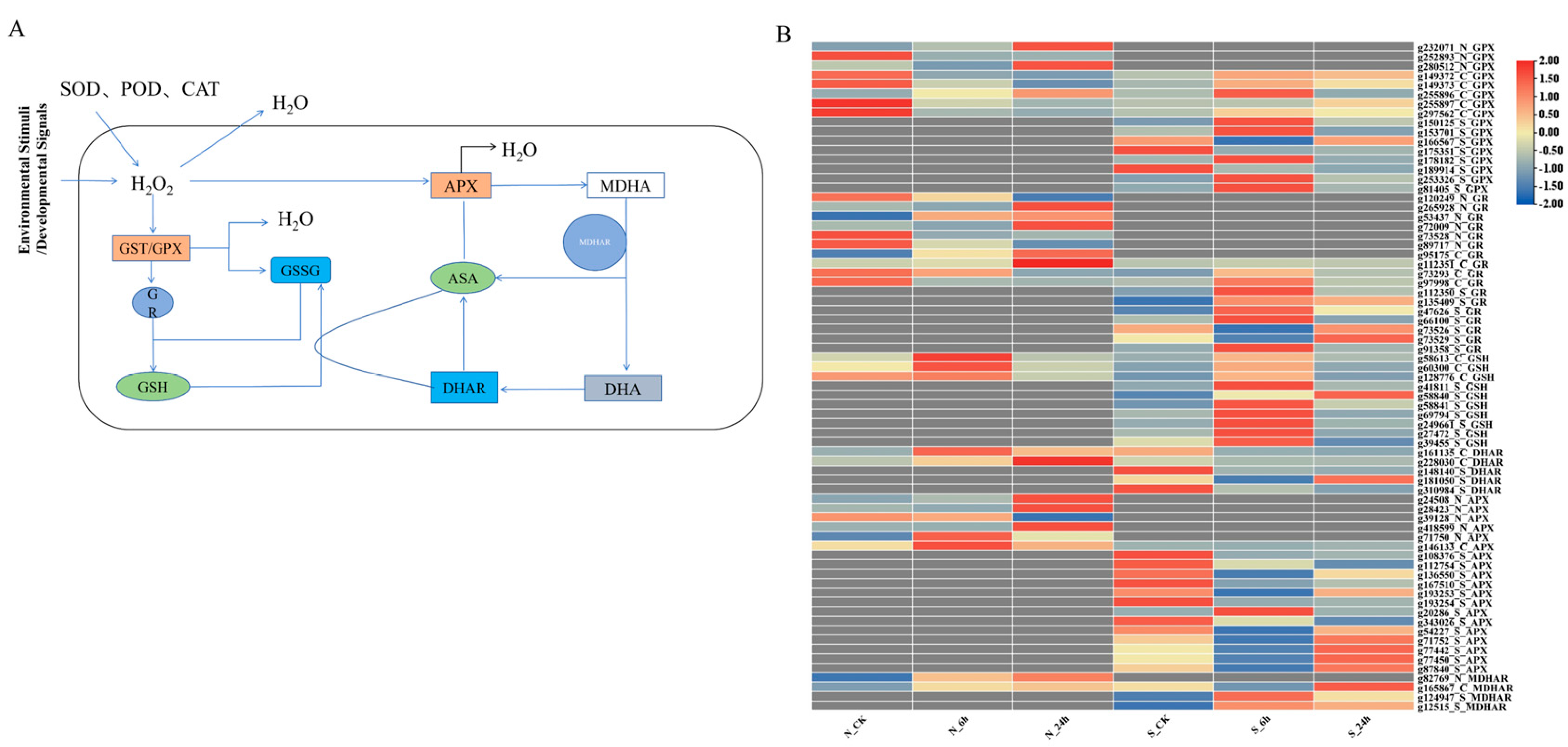
3.5. Analysis of DEGs Associated with ROS Scavenging
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hassani, A.; Azapagic, A.; Shokri, N. Global predictions of primary soil salinization under changing climate in the 21st century. Nat. Commun. 2021, 12, 6663. [Google Scholar] [PubMed]
- Zhou, T.; Lv, Q.; Zhang, L.; Fan, J.; Wang, T.; Meng, Y.; Xia, H.; Ren, X.; Hu, S. Converted paddy to upland in saline-sodic land could improve soil ecosystem multifunctionality by enhancing soil quality and alleviating microbial metabolism limitation. Sci. Total Environ. 2024, 924, 171707. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Gang, N.I.; Feng, G.; Burrill, H.M.; Jianfang, L.I.; Zhang, J.; Zhang, F. Saline-alkali soil reclamation and utilization in China: Progress and prospects. Front. Agric. Sci. 2024, 11, 216–228. [Google Scholar]
- Tang, H.; Du, L.; Xia, C.; Luo, J. Bridging gaps and seeding futures: A synthesis of soil salinization and the role of plant-soil interactions under climate change. iScience 2024, 27, 110804. [Google Scholar] [CrossRef]
- Johnson, R.C. Salinity Resistance, Water Relations, and Salt Content of Crested and Tall Wheatgrass Accessions. Crop Sci. 1991, 31, 730–734. [Google Scholar]
- Bahrani, M.J.; Bahrami, H.; Haghighi, A.A.K. Effect of water stress on ten forage grasses native or introduced to Iran. Grassl. Sci. 2010, 56, 1–5. [Google Scholar] [CrossRef]
- Pourrezaei, J.; Khajeddin, S.J.; Karimzadeh, H.R.; Vahabi, M.R.; Mozaffarian, V.; Esfahani, M.T. Roadside flora in arid and semi-arid natural areas (Case study: Northern Khorasan province, Iran). Univ. Tehran 2017, 22, 229–237. [Google Scholar]
- Jafari, A.A.; Seydemohammadi, A.; Abdi, N.A.; Madaharefi, H. Seed and hay production in 31 genotypes of desert wheatgrass (Agropyron desertorum) using drought tolerance indices. Iran. J. Range Desert Res. 2008, 15, 114–128. [Google Scholar]
- Rittenhouse, L.R.; Sneva, F.A. Early growth of Nordan crested wheatgrass and Sherman big bluegrass. J. Range Manag. 1977, 30, 236–237. [Google Scholar] [CrossRef]
- Wenshu, Z. The Test of Twenty—Four Canadian Forage Varieties in Beijing(1986–1989). Pratacultural Sci. 1993, 4, 7–10. [Google Scholar]
- He, X.; Li, Q.-F.; Lu, H.-P. Studies on seeding characteristics in Elymus sibilicus and Agropyron desertorum cv. nordan. Pratacult. Sci. 2004, 21, 31–39. [Google Scholar]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Zheng, B. Phytohormones Regulate Accumulation of Osmolytes Under Abiotic Stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef] [PubMed]
- Pandhair, V.; Sekhon, B.S. Reactive Oxygen Species and Antioxidants in Plants: An Overview. J. Plant Biochem. Biotechnol. 2006, 15, 71–78. [Google Scholar]
- Anjum, N.A.; Chan, M.T.; Umar, S. Ascorbate-Glutathione Pathway and Stress Tolerance in Plants; Springer: Dordrecht, The Netherlands, 2010. [Google Scholar]
- Latowski, D.; Surówka, E.; Strzałka, K. Regulatory Role of Components of Ascorbate–Glutathione Pathway in Plant Stress Tolerance. In Ascorbate-Glutathione Pathway and Stress Tolerance in Plants; Anjum, N.A., Chan, M.-T., Umar, S., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 1–53. [Google Scholar]
- Pang, C.H.; Wang, B.S. Role of Ascorbate Peroxidase and Glutathione Reductase in Ascorbate–Glutathione Cycle and Stress Tolerance in Plants; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Murtaza, G.; Deng, G.; Usman, M.; Jamil, A.; Qasim, M.; Iqbal, J.; Ercisli, S.; Akram, M.I.; Rizwan, M.; Elshikh, M.S.; et al. Impact of Acacia-derived biochar to mitigate salinity stress in Zea mays L. by morpho-physiological and biochemical indices. Sci. Rep. 2024, 14, 1–15. [Google Scholar]
- Imran, A.; Ghosh, A. Evolutionary expansion, functional diversification, and transcript profiling of plant Glutathione Peroxidases. Plant Sci. 2024, 341, 111991. [Google Scholar] [CrossRef]
- Gardarin, A.; Coste, F.O.; Wagner, M.-H.; Dürr, C. How do seed and seedling traits influence germination and emergence parameters in crop species? A comparative analysis. Seed Sci. Res. 2017, 26, 317–331. [Google Scholar]
- Ouyang, S.Q.; Liu, Y.F.; Liu, P.; Lei, G.; Chen, S.Y. Receptor-like kinase ossik1 improves drought and salt stress tolerance in rice (Oryza sativa) plants. Plant J. 2010, 62, 316–329. [Google Scholar]
- Mehta, P.; Jajoo, A.; Mathur, S.; Bharti, S. Chlorophyll a fluorescence study revealing effects of high salt stress on photosystem ii in wheat leaves. Plant Physiol. Biochem. 2010, 48, 16–20. [Google Scholar]
- Stepien, P.; Johnson, G.N. Contrasting Responses of Photosynthesis to Salt Stress in the Glycophyte Arabidopsis and the Halophyte Thellungiella: Role of the Plastid Terminal Oxidase as an Alternative Electron Sink. Plant Physiol. 2009, 149, 1154–1165. [Google Scholar]
- Bidgoli, R.D. Exploring the Saltiness Resilience of one Rangeland’s Species (Agropyron desertorum) in Several Phenological Stages. Desert Ecosyst. Eng. J. 2023, 12, 15–24. [Google Scholar]
- Long, N.V.; Dolstra, O.; Malosetti, M.; Kilian, B.; Graner, A. Association mapping of salt tolerance in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2013, 126, 2335–2351. [Google Scholar] [PubMed]
- Zhang, Z.; Liu, L.; Li, H.; Zhang, S.; Fu, X.; Zhai, X.; Yang, N.; Shen, J.; Li, R.; Li, D. Exogenous Melatonin Promotes the Salt Tolerance by Removing Active Oxygen and Maintaining Ion Balance in Wheat. Front. Plant Sci. 2021, 12, 787062. [Google Scholar]
- Wang, J.; Yan, D.; Liu, R.; Wang, T.; Lian, Y.; Lu, Z.; Hong, Y.; Wang, Y.; Li, R. The Physiological and Molecular Mechanisms of Exogenous Melatonin Promote the Seed Germination of Maize (Zea mays L.) under Salt Stress. Plants 2024, 13, 2142. [Google Scholar] [CrossRef]
- Chunthaburee, S.; Dongsansuk, A.; Sanitchon, J.; Pattanagul, W.; Theerakulpisut, P. Physiological and biochemical parameters for evaluation and clustering of rice cultivars differing in salt tolerance at seedling stage. Saudi J. Biol. Sci. 2016, 23, 467–477. [Google Scholar]
- Wang, W.; Zhao, Y.; Wang, W.; Xu, K.; Niu, L.; Yu, L.; Zou, J.W.; Wang, Z.; Zhang, Y.J.; Wang, F.; et al. Evaluation of Salt Tolerance Mechanism and Study on Salt Tolerance Relationship of Different Salt-Tolerant Wheat Varieties. Comput. Intell. Neurosci. 2022, 2022, 2395568. [Google Scholar]
- Ali, Q.; Sami, A.; Haider, M.Z.; Ashfaq, M.; Javed, M.A. Antioxidant production promotes defense mechanism and different gene expression level in Zeamays under abiotic stress. Sci. Rep. 2024, 14, 7114. [Google Scholar]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar]
- Dewey, C.N.; Bo, L. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar]
- Anders, S.; Huber, W. Differential expression of RNA-Seq data at the gene level–the DESeq package. Eur. Mol. Biol. Lab. (EMBL) 2012, 10, f1000research. [Google Scholar]
- Ullah, F.; Yi, M. Mechanisms of ROS Regulation of Plant Development and Stress Responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Parvin, K.; Bhuiyan, T.F.; Fujita, M. Regulation of ROS Metabolism in Plants under Environmental Stress: A Review of Recent Experimental Evidence. Int. J. Mol. Sci. 2020, 21, 8695. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.-Y.; Li, Y.-T.; Liu, L.-J.; Dong, X.-Y.; Zhang, X.-F.; Wang, X.-H.; Zhao, Y. Germination characteristics and evaluation of 14 green manure plants under salt stress. Grassland 2019, 39, 98–105. [Google Scholar]
- Lei, S.; Jie, Z.; Meng-Chen, Z.; Dong-Mei, W. Study on the Genotypes of Soybean Varieties under Different NaCl Concentration Stress. Acta Agric. Boreali-Sin. 2010, 2, 2104–2108. [Google Scholar]
- Li, X.-H.; Chen, Q.-Y.; Ou, X.-Q.; Qiao, H.; Wang, Z.-J. Effects of NaCl Stress on Seed Germination and Seedling Growth of Different Wheat Varieties. Hubei Agric. Sci. 2017, 56, 3222–3224. [Google Scholar]
- Ali, S.; Tyagi, A.; Bae, H. ROS interplay between plant growth and stress biology: Challenges and future perspectives. Plant Physiol. Biochem. 2023, 203, 108032. [Google Scholar] [CrossRef]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar]
- Singh, A.; Satheeshkumar, P.K. Reactive Oxygen Species (ROS) and ROS Scavengers in Plant Abiotic Stress Response. In Stress Biology in Photosynthetic Organisms: Molecular Insights and Cellular Responses; Mishra, A.K., Ed.; Springer: Singapore, 2024; pp. 41–63. [Google Scholar]
- Luchli, A.; Grattan, S.R. Plant Growth And Development Under Salinity Stress; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Breusegem, F.V. ROS signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar]
- Díaz De León, J.; Escoppinichi, R.; Zavala-Fonseca, R.; Castellanos, T.; Roder, M.; Mujeeb-Kazi, A. Phenotypic and genotypic characterization of salt-tolerant wheat genotypes. Cereal Res. Commun. 2010, 38, 15–22. [Google Scholar]
- Sheoran, P.; Kamboi, P.; Kumar, A. Varietal led ecological resilience: Through KRL 210 in salt stressed wheat. Indian Farming 2022, 72, 47–49. [Google Scholar]
- Patwa, N.; Pandey, V.; Gupta, O.P.; Yadav, A.; Meena, M.R.; Ram, S.; Singh, G. Unravelling wheat genotypic responses: Insights into salinity stress tolerance in relation to oxidative stress, antioxidant mechanisms, osmolyte accumulation and grain quality parameters. BMC Plant Biol. 2024, 24, 1–17. [Google Scholar]
- Zhang, Y.F.; Bo, Y. Influences of salt and alkali mixed stresses on antioxidative activity and MDA content of Medicago sativa at seedling stage. Acta Prataculturae Sin. 2009, 18, 46. [Google Scholar]
- Ezatollah, E.; Fariborz, S.; Farid, S.; Manouchehr, E. The effect of salt stress on antioxidant enzymes’ activity and lipid peroxidation on the wheat seedling. Not. Bot. Horti Agrobot. Cluj-Napoca 2007, 35, 48. [Google Scholar]
- Raheleh, M.M.; Zahra-Sadat, S.; Nadali, B.J.; Reza, G.M.; Ghorban-Ali, N.; Saeedeh, A. Dissecting molecular mechanisms underlying salt tolerance in rice: A comparative transcriptional profiling of the contrasting genotypes. J. Rice 2019, 12, 13. [Google Scholar]
- An, S.; Wei, W.; Shugao, F.; Xiao, X.; Yanling, Y.; Amombo, E.; Xiaoning, L.; Guangyang, W.; Hongli, W.; Jinmin, F. Comprehensive transcriptional analysis reveals salt stress-regulated key pathways, hub genes and time-specific responsive gene categories in common bermudagrass (Cynodon dactylon (L.) Pers.) roots %J BMC Plant Biology. BMC Plant Biol. 2021, 21, 175. [Google Scholar]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Fujita, M. Regulation of Ascorbate-Glutathione Pathway in Mitigating Oxidative Damage in Plants under Abiotic Stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef]
- Dossa, K.; Mmadi, M.A.; Zhou, R.; Zhang, T.; Zhang, X. Depicting the Core Transcriptome Modulating Multiple Abiotic Stresses Responses in Sesame (Sesamum indicum L.). Int. J. Mol. Sci. 2019, 20, 3930. [Google Scholar] [CrossRef]
- Hua, M.J.; Lei, Z.H.; Qiu, Z.Z.; Guang, Z.C. Progress in Mechanisms of Plant Resistance to Salt Stress. Life Sci. Res. 2001, 12, 2253. [Google Scholar]
- Zhang, L.; Wang, S.; Yang, X.; He, L.; Hu, L.; Tang, R.; Li, J.; Liu, Z. Physiological and Multi-Omics Integrative Analysis Provides New Insights into Tolerance to Waterlogging Stress in Sesame (Sesamum indicum L.). Int. J. Mol. Sci. 2025, 26, 351. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salt Tolerance Index | Concentration/% | |
|---|---|---|
| NaCl | ||
| S | N | |
| Suitable concentration of salt tolerance | <90 mm | <30 mM |
| Salt-tolerant half-lethal concentration | <60 mM | <60 mM |
| Salt tolerance limit concentration | <180 mM | <180 mM |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Huang, X.; Han, X.; Yang, H.; Zhao, Y. Comparative Analysis of Salt Tolerance and Transcriptomics in Two Varieties of Agropyron desertorum at Different Developmental Stages. Genes 2025, 16, 367. https://doi.org/10.3390/genes16040367
Li Y, Huang X, Han X, Yang H, Zhao Y. Comparative Analysis of Salt Tolerance and Transcriptomics in Two Varieties of Agropyron desertorum at Different Developmental Stages. Genes. 2025; 16(4):367. https://doi.org/10.3390/genes16040367
Chicago/Turabian StyleLi, Yuchen, Xintian Huang, Xiao Han, Hui Yang, and Yan Zhao. 2025. "Comparative Analysis of Salt Tolerance and Transcriptomics in Two Varieties of Agropyron desertorum at Different Developmental Stages" Genes 16, no. 4: 367. https://doi.org/10.3390/genes16040367
APA StyleLi, Y., Huang, X., Han, X., Yang, H., & Zhao, Y. (2025). Comparative Analysis of Salt Tolerance and Transcriptomics in Two Varieties of Agropyron desertorum at Different Developmental Stages. Genes, 16(4), 367. https://doi.org/10.3390/genes16040367