
Genetic and Epigenetic Diversity of Pinus pinea L.: Conservation Implications for Priority Populations in Greece

 ,
,  ,
,  ,
,
Abstract
1. Introduction
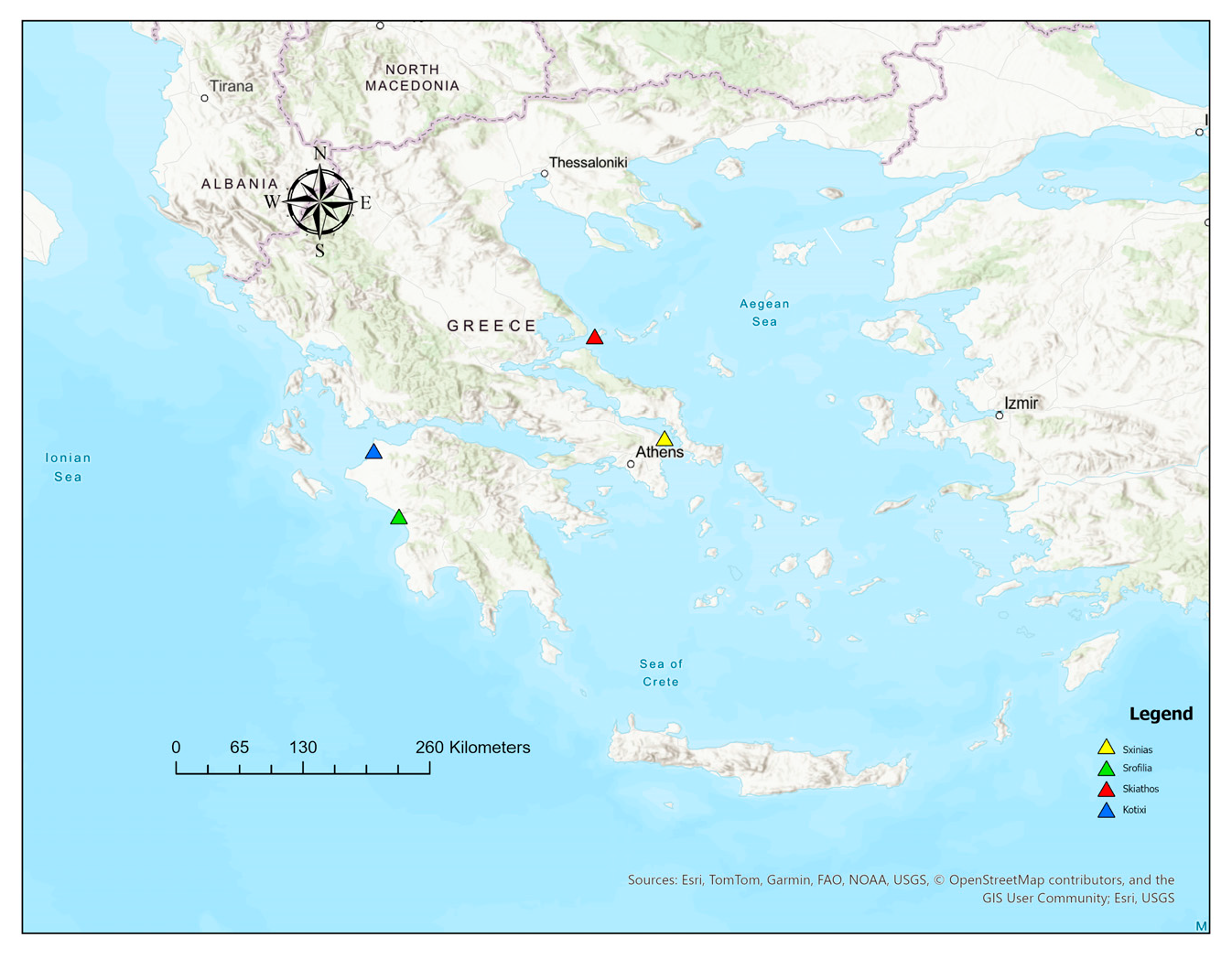
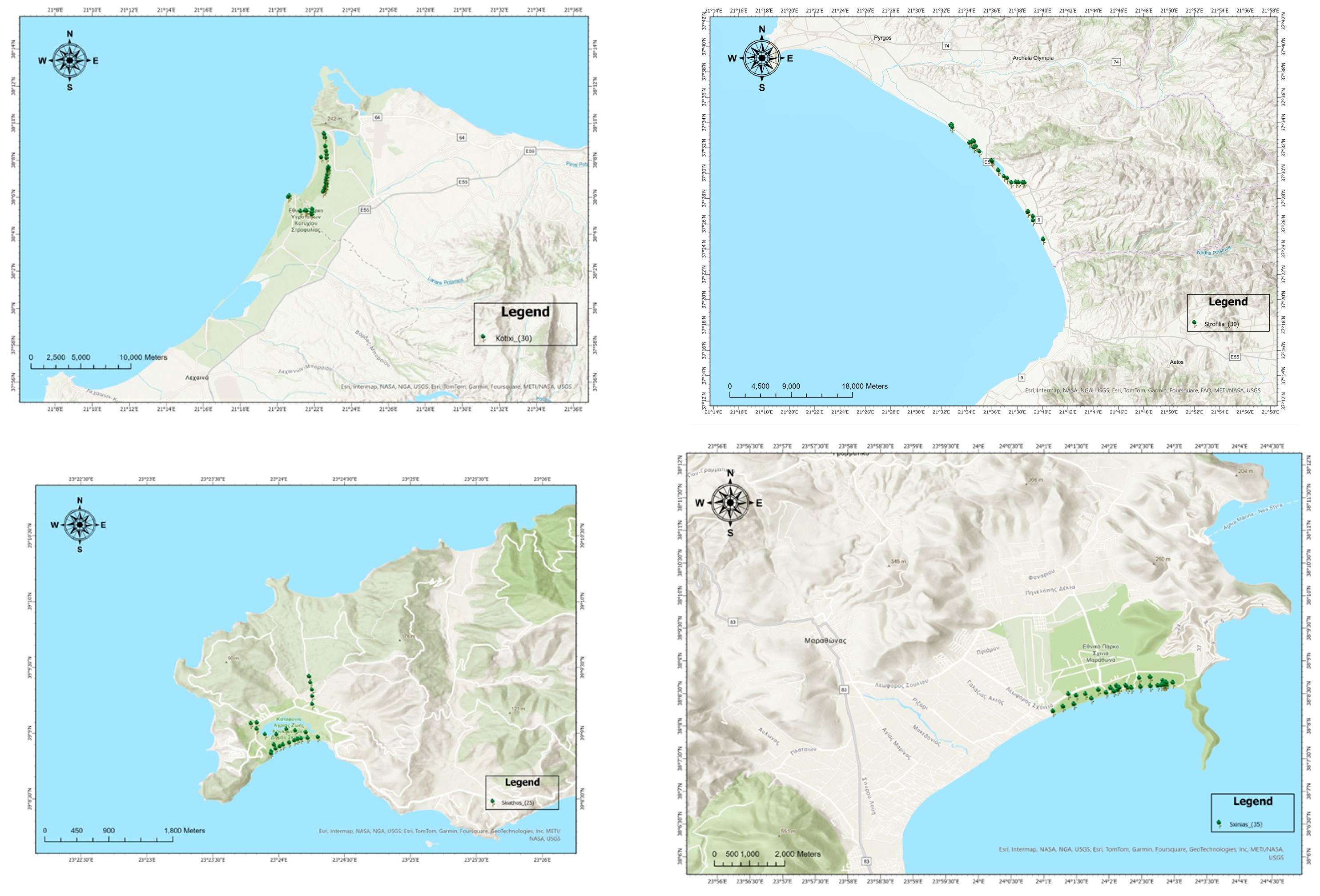
2. Materials and Methods
2.1. AFLP Procedure
2.2. MSAP Procedure
2.3. Needle Water Potential
2.4. Data Collection and Statistical Analysis
3. Results
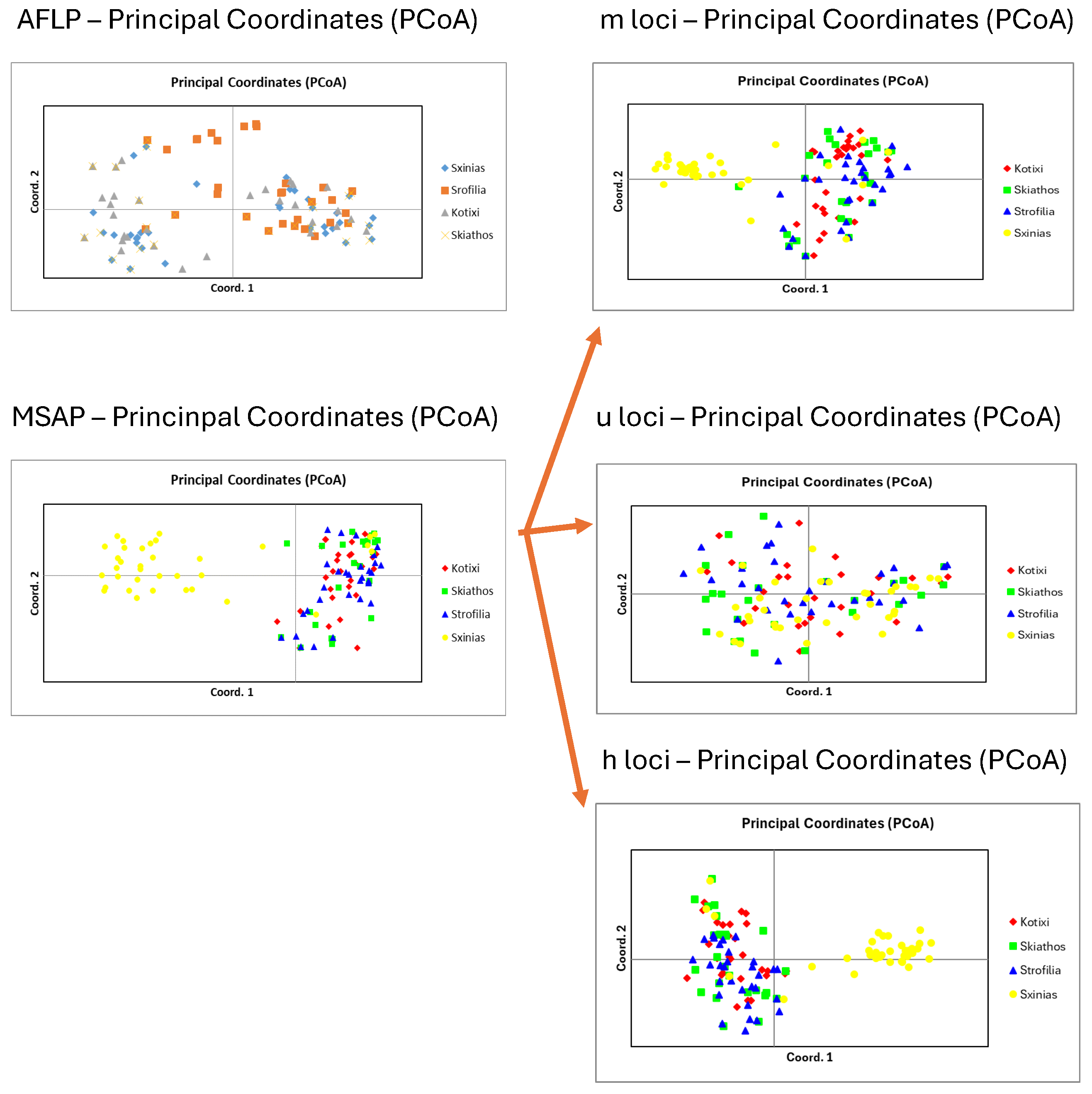
3.1. Genetic Diversity
3.2. Epigenetic Diversity
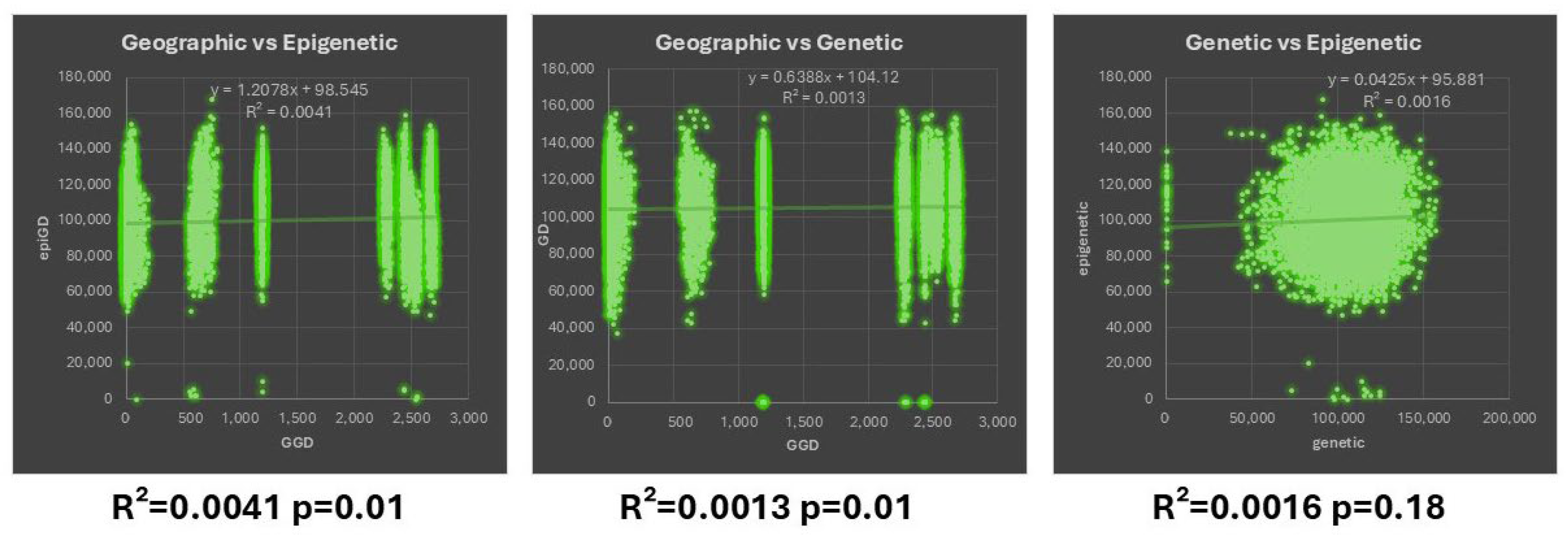
3.3. Correlation Between Geographic—Genetic and Epigenetic Variability
3.4. Correlation Between Epigenetic Variability and Tree Water Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Correia, A.C.; Faias, S.P.; Ruiz-Peinado, R.; Chianucci, F.; Cutini, A.; Fontes, L.; Manetti, M.; Montero, G.; Soares, P.; Tomé, M. Generalized biomass equations for Stone pine (Pinus pinea L.) across the Mediterranean basin. For. Ecol. Manag. 2018, 429, 425–436. [Google Scholar] [CrossRef]
- Mutke, S.; Calama, R.; González-Martínez, S.C.; Montero, G.; Javier Gordo, F.; Bono, D.; Gil, L. 4 mediterranean stone pine: Botany and horticulture. Hortic. Rev. 2012, 39, 153–201. [Google Scholar]
- Fady, B.; Esposito, E.; Abulaila, K.; Aleksic, J.M.; Alia, R.; Alizoti, P.; Apostol, E.-N.; Aravanopoulos, P.; Ballian, D.; Kharrat, M.B.D. Forest genetics research in the Mediterranean Basin: Bibliometric analysis, knowledge gaps, and perspectives. Curr. For. Rep. 2022, 8, 277–298. [Google Scholar]
- Martínez, F.; Montero, G. The Pinus pinea L. woodlands along the coast of South-western Spain: Data for a new geobotanical interpretation. Plant Ecol. 2004, 175, 1–18. [Google Scholar]
- Hlaiem, S.; Yangui, I.; Rocca, G.D.; Barberini, S.; Danti, R.; Jamâa, M.L.B. First report of Pestalotiopsis biciliata associated with twig canker and dieback of Pinus pinea in Tunisia. J. Plant Pathol. 2022, 104, 393–394. [Google Scholar] [CrossRef]
- Viñas, R.A.; Caudullo, G.; Oliveira, S.; De Rigo, D. Pinus pinea in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; Publication Office of the European Union: Luxembourg, 2016; p. 204. [Google Scholar]
- Mutke, S.; Vendramin, G.G.; Fady, B.; Bagnoli, F.; González-Martínez, S.C. Molecular and quantitative genetics of stone pine (Pinus pinea). In Genetic Diversity in Horticultural Plants; Springer: Cham, Switzerland, 2019; pp. 61–84. [Google Scholar]
- Fady, B.; Fineschi, S.; Vendramin, G. EUFORGEN Technical Guidelines for Genetic Conservation and Use for Italian Stone Pine (Pinus pinea); Bioversity International: Rome, Italy, 2004. [Google Scholar]
- Nergiz, C.; Dönmez, I. Chemical composition and nutritive value of Pinus pinea L. seeds. Food Chem. 2004, 86, 365–368. [Google Scholar]
- Calama, R.; Madrigal, G.; Candela, J.; Montero, G. Effects of fertilization on the production of an edible forest fruit: Stone pine (Pinus pinea L.) nuts in south-west Andalusia. For. Syst. 2007, 16, 241–252. [Google Scholar]
- Simões, A.S.B.; Borges, M.M.; Grazina, L.; Nunes, J. Stone pine (Pinus pinea L.) high-added-value genetics: An overview. Genes 2024, 15, 84. [Google Scholar] [CrossRef]
- Awan, H.U.M.; Pettenella, D. Pine nuts: A review of recent sanitary conditions and market development. Forests 2017, 8, 367. [Google Scholar] [CrossRef]
- Mechergui, K.; Naghmouchi, S.; Altamimi, A.S.; Jaouadi, W. Evaluation of biomass, carbon storage capability, agroforestry interest of Pinus pinea L. and management practices to increase carbon stocks: A review. CERNE 2021, 27, e102938. [Google Scholar]
- Manso, R.; Fortin, M.; Calama, R.; Pardos, M. Modelling seed germination in forest tree species through survival analysis. The Pinus pinea L. case study. For. Ecol. Manag. 2013, 289, 515–524. [Google Scholar] [CrossRef]
- Pereira, S.; Prieto, A.; Calama, R.; Diaz-Balteiro, L. Optimal management in Pinus pinea L. stands combining silvicultural schedules for timber and cone production. Silva Fenn. 2015, 49, 1226. [Google Scholar] [CrossRef]
- Biocca, M.; Gallo, P.; Sperandio, G. Technical and economic analysis of Stone pine (Pinus pinea L.) maintenance in urban areas. Trees For. People 2021, 6, 100162. [Google Scholar] [CrossRef]
- Jaramillo-Correa, J.P.; Bagnoli, F.; Grivet, D.; Fady, B.; Aravanopoulos, F.A.; Vendramin, G.G.; González-Martínez, S.C. Evolutionary rate and genetic load in an emblematic Mediterranean tree following an ancient and prolonged population collapse. Mol. Ecol. 2020, 29, 4797–4811. [Google Scholar] [CrossRef]
- Sáez-Laguna, E.; Guevara, M.-Á.; Díaz, L.-M.; Sánchez-Gómez, D.; Collada, C.; Aranda, I.; Cervera, M.-T. Epigenetic variability in the genetically uniform forest tree species Pinus pinea L. PLoS ONE 2014, 9, e103145. [Google Scholar] [CrossRef]
- Maggini, F.; Baldassini, S. Ribosomal RNA genes in the genus Pinus. I. Caryologia 1995, 48, 17–25. [Google Scholar] [CrossRef]
- Krupkin, A.; Liston, A.; Strauss, S. Phylogenetic analysis of the hard pines (Pinus subgenus Pinus, Pinaceae) from chloroplast DNA restriction site analysis. Am. J. Bot. 1996, 83, 489–498. [Google Scholar] [CrossRef]
- Georgolopoulos, G.; Parducci, L.; Drouzas, A.D. A short phylogenetically informative cpDNA fragment for the identification of Pinus species. Biochem. Syst. Ecol. 2016, 66, 166–172. [Google Scholar] [CrossRef]
- Ballin, N.Z.; Mikkelsen, K. Polymerase chain reaction and chemometrics detected several Pinus species including Pinus armandii involved in pine nut syndrome. Food Control 2016, 64, 234–239. [Google Scholar] [CrossRef]
- Alonso, P.; Cortizo, M.; Cantón, F.R.; Fernández, B.; Rodríguez, A.; Centeno, M.L.; Cánovas, F.M.; Ordás, R.J. Identification of genes differentially expressed during adventitious shoot induction in Pinus pinea cotyledons by subtractive hybridization and quantitative PCR. Tree Physiol. 2007, 27, 1721–1730. [Google Scholar] [CrossRef]
- Gad, M.; Mohamed, S. Phylogenetic evaluation of some Pinus species from different genetic resources using protein, isozymes, RAPD and ISSR analyses. J. Am. Sci. 2012, 8, 311–321. [Google Scholar]
- González-Martínez, S.C.; Robledo-Arnuncio, J.J.; Collada, C.; Díaz, A.; Williams, C.; Alía, R.; Cervera, M.T. Cross-amplification and sequence variation of microsatellite loci in Eurasian hard pines. Theor. Appl. Genet. 2004, 109, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Vendramin, G.G.; Fady, B.; González-Martínez, S.C.; Hu, F.S.; Scotti, I.; Sebastiani, F.; Soto, A.; Petit, R.J. Genetically depauperate but widespread: The case of an emblematic Mediterranean pine. Evolution 2008, 62, 680–688. [Google Scholar] [CrossRef]
- Pinzauti, F.; Sebastiani, F.; Budde, K.B.; Fady, B.; González-Martínez, S.C.; Vendramin, G. Nuclear microsatellites for Pinus pinea (Pinaceae), a genetically depauperate tree, and their transferability to P. halepensis. Am. J. Bot. 2012, 99, e362–e365. [Google Scholar]
- Katsidi, E.C.; Avramidou, E.V.; Ganopoulos, I.; Barbas, E.; Doulis, A.; Triantafyllou, A.; Aravanopoulos, F.A. Genetics and epigenetics of Pinus nigra populations with differential exposure to air pollution. Front. Plant Sci. 2023, 14, 1139331. [Google Scholar] [CrossRef]
- Lira-Medeiros, C.F.; Parisod, C.; Fernandes, R.A.; Mata, C.S.; Cardoso, M.A.; Ferreira, P.C.G. Epigenetic variation in mangrove plants occurring in contrasting natural environment. PLoS ONE 2010, 5, e10326. [Google Scholar]
- Ganatsas, P.; Thanasis, G. Pinus halepensis invasion in Pinus pinea habitat in Strofylia forest (Site of NATURA 2000 network), southern Greece. J. Nat. Conserv. 2010, 18, 106–117. [Google Scholar]
- Gaitanis, A.; Kalogeropoulos, K.; Detsis, V.; Chalkias, C. Monitoring 60 years of Land cover change in the Marathon Area, Greece. Land 2015, 4, 337–354. [Google Scholar] [CrossRef]
- Petrakis, P.V.; Koulelis, P.P.; Solomou, A.D.; Spanos, K.; Spanos, I.; Feest, A. Insect diversity in the coastal pinewood and Marsh at Schinias, Marathon, Greece: Impact of management decisions on a degraded biotope. Forests 2023, 14, 392. [Google Scholar] [CrossRef]
- Keeley, J.E. Ecology and evolution of pine life histories. Ann. For. Sci. 2012, 69, 445–453. [Google Scholar] [CrossRef]
- Fady, B. Biogeography of neutral genes and recent evolutionary history of pines in the Mediterranean Basin. Ann. For. Sci. 2012, 69, 421–428. [Google Scholar] [CrossRef]
- Pham, T.M.; Nguyen, H.C.; Nguyen, V.K.; Pham, H.H.; Nguyen, N.T.; Dang, G.T.H.; Dinh, H.T.; Pham, T.A. Application of the Worldwide Bioclimatic Classification System to determine bioclimatic features and potential natural vegetation distribution in Van Chan district, Vietnam. Trop. Ecol. 2023, 64, 765–780. [Google Scholar] [CrossRef]
- Avramidou, E.V.; Korakaki, E.; Malliarou, E.; Boutsios, S. Studying the genetic and the epigenetic diversity of the endangered species Juniperus drupacea Labill. towards safeguarding its conservation in Greece. Forests 2023, 14, 1271. [Google Scholar] [CrossRef]
- Vekemans, X.; Beauwens, T.; Lemaire, M.; Roldán-Ruiz, I. Data from amplified fragment length polymorphism (AFLP) markers show indication of size homoplasy and of a relationship between degree of homoplasy and fragment size. Mol. Ecol. 2002, 11, 139–151. [Google Scholar] [CrossRef]
- Schulz, B.; Eckstein, R.L.; Durka, W. Scoring and analysis of methylation-sensitive amplification polymorphisms for epigenetic population studies. Mol. Ecol. Resour. 2013, 13, 642–653. [Google Scholar] [CrossRef]
- Durka, W. MSAP_calc.r-R-FUNCTIONS for Transformation and Analysis of MSAP Data. 2012. Available online: https://scholar.google.com/scholar?hl=el&as_sdt=0%2C5&q=39.%09Durka%2C+W.+MSAP_calc.+r-R-functions+for+transformation+and+analysis+of+MSAP+data.++2012.&btnG= (accessed on 17 January 2024).
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar]
- Richards, E.J. Inherited epigenetic variation—Revisiting soft inheritance. Nat. Rev. Genet. 2006, 7, 395–401. [Google Scholar]
- Bräutigam, K.; Vining, K.J.; Lafon-Placette, C.; Fossdal, C.G.; Mirouze, M.; Marcos, J.G.; Fluch, S.; Fraga, M.F.; Guevara, M.Á.; Abarca, D. Epigenetic regulation of adaptive responses of forest tree species to the environment. Ecol. Evol. 2013, 3, 399–415. [Google Scholar] [CrossRef]
- Alvarez-Venegas, R.; De la Peña, C.; Casas-Mollano, J.A.; Pascual, J.; Cañal, M.J.; Correia, B.; Escandon, M.; Hasbún, R.; Meijón, M.; Pinto, G. Can epigenetics help forest plants to adapt to climate change? In Epigenetics in Plants of Agronomic Importance: Fundamentals and Applications: Transcriptional Regulation and Chromatin Remodelling in Plants; Springer: Cham, Switzerland, 2014; pp. 125–146. [Google Scholar]
- Amaral, J.; Ribeyre, Z.; Vigneaud, J.; Sow, M.D.; Fichot, R.; Messier, C.; Pinto, G.; Nolet, P.; Maury, S. Advances and promises of epigenetics for forest trees. Forests 2020, 11, 976. [Google Scholar] [CrossRef]
- Sow, M.D.; Allona, I.; Ambroise, C.; Conde, D.; Fichot, R.; Gribkova, S.; Jorge, V.; Le-Provost, G.; Pâques, L.; Plomion, C. Epigenetics in forest trees: State of the art and potential implications for breeding and management in a context of climate change. Adv. Bot. Res. 2018, 88, 387–453. [Google Scholar]
- García-García, I.; Méndez-Cea, B.; Martín-Gálvez, D.; Seco, J.I.; Gallego, F.J.; Linares, J.C. Challenges and perspectives in the epigenetics of climate change-induced forests decline. Front. Plant Sci. 2022, 12, 797958. [Google Scholar] [CrossRef] [PubMed]
- Rey, O.; Eizaguirre, C.; Angers, B.; Baltazar-Soares, M.; Sagonas, K.; Prunier, J.G.; Blanchet, S. Linking epigenetics and biological conservation: Towards a conservation epigenetics perspective. Funct. Ecol. 2020, 34, 414–427. [Google Scholar] [CrossRef]
- Williams, B.R.; Miller, A.J.; Edwards, C.E. How do threatened plant species with low genetic diversity respond to environmental stress? Insights from comparative conservation epigenomics and phenotypic plasticity. Mol. Ecol. Resour. 2023. [Google Scholar] [CrossRef]
- Wu, W.-Q.; Yi, M.R.; Wang, X.-F.; Ma, L.-L.; Jiang, L.; Li, X.-W.; Xiao, H.-X.; Sun, M.-Z.; Li, L.-F.; Liu, B. Genetic and epigenetic differentiation between natural Betula ermanii (Betulaceae) populations inhabiting contrasting habitats. Tree Genet. Genomes 2013, 9, 1321–1328. [Google Scholar] [CrossRef]
- Richards, C.L.; Alonso, C.; Becker, C.; Bossdorf, O.; Bucher, E.; Colomé-Tatché, M.; Durka, W.; Engelhardt, J.; Gaspar, B.; Gogol-Döring, A. Ecological plant epigenetics: Evidence from model and non-model species, and the way forward. Ecol. Lett. 2017, 20, 1576–1590. [Google Scholar] [CrossRef]
- Batalova, A.Y.; Krutovsky, K.V. Genetic and epigenetic mechanisms of longevity in forest trees. Int. J. Mol. Sci. 2023, 24, 10403. [Google Scholar] [CrossRef]
- Fallour, D.; Fady, B.; Lefevre, F. Study on isozyme variation in Pinus pinea L.: Evidence for low polymorphism. Silvae Genet. 1997, 46, 201–206. [Google Scholar]
- Mutke, S.; Gordo, J.; Khouja, M.; Fady, B. Low genetic and high environmental diversity at adaptive traits in Pinus pinea from provenance tests in France and Spain. Options Méditerranéennes A 2013, 105, 73–79. [Google Scholar]
- Do Nascimento, A.M.M.; Montalbán, I.A.; Llamazares De Miguel, D.; Goicoa, T.; Ugarte, M.D.; Moncaleán, P. High temperature and water deficit cause epigenetic changes in somatic plants of Pinus radiata D. Don. Plant Cell Tissue Organ Cult. (PCTOC) 2022, 151, 107–121. [Google Scholar] [CrossRef]
- Miryeganeh, M.; Armitage, D.W. Epigenetic responses of trees to environmental stress in the context of climate change. Biol. Rev. 2024, 100, 131–148. [Google Scholar] [CrossRef] [PubMed]
- Chambel, M.R.; Climent, J.; Alía, R. Divergence among species and populations of Mediterranean pines in biomass allocation of seedlings grown under two watering regimes. Ann. For. Sci. 2007, 64, 87–97. [Google Scholar]
- Xu, J.; Zhou, S.; Gong, X.; Song, Y.; van Nocker, S.; Ma, F.; Guan, Q. Single-base methylome analysis reveals dynamic epigenomic differences associated with water deficit in apple. Plant Biotechnol. J. 2018, 16, 672–687. [Google Scholar]
- Le Gac, A.-L.; Lafon-Placette, C.; Chauveau, D.; Segura, V.; Delaunay, A.; Fichot, R.; Marron, N.; Le Jan, I.; Berthelot, A.; Bodineau, G. Winter-dormant shoot apical meristem in poplar trees shows environmental epigenetic memory. J. Exp. Bot. 2018, 69, 4821–4837. [Google Scholar]
- González-Benito, M.E.; Ibáñez, M.Á.; Pirredda, M.; Mira, S.; Martín, C. Application of the MSAP technique to evaluate epigenetic changes in plant conservation. Int. J. Mol. Sci. 2020, 21, 7459. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | 5′ to 3′ Sequence |
|---|---|
| EcoRI adapter | CTCGTAGACTGCGTACC AATTGGTACGCAGTC |
| MseI adapter | GACGATGAGTCCTGAG TACTCAGGACTCAT |
| HpaII/MspI adapter | GACGATGAGTCTCGAT CGATCGAGACTCAT |
| Pre-selective EcoRI primer | GACTGCGTACCAATTC-A |
| Pre-selective MseI primer | GATGAGTCCTGAGTAA-C |
| Pre-selective HpaII/MspI primer | ATGAGTCTCGATCGG-A |
| Selective EcoRI primers | GACTGCGTACCAATTC + ATG (FAM) GACTGCGTACCAATTC + ACT (HEX) GACTGCGTACCAATTC + AAC (ROX) GACTGCGTACCAATTC + AAG (TAMRA) |
| Selective MseI primer | GATGAGTCCTGAGTAA-CAA GATGAGTCCTGAGTAA-CAC GATGAGTCCTGAGTAA-CGT GATGAGTCCTGAGTAA-CTC |
| Selective HpaII/MspI primer | ATGAGTCTCGATCGGATC ATGAGTCTCGATCGGACT ATGAGTCTCGATCGGAAT |
| EcoRI adapter | CTCGTAGACTGCGTACC AATTGGTACGCAGTC |
| AFLP | MSAP | |||||||
|---|---|---|---|---|---|---|---|---|
| Population | P | I | He | Pepi | Iepi | Hepi | N.B. | N.P.B. |
| Schinias | 71.14 | 0.14 | 0.077 | 77.37 | 0.16 | 0.088 | 564 | 122 |
| Strofilia | 62.61 | 0.14 | 0.074 | 61.73 | 0.14 | 0.080 | 452 | 29 |
| Kotixi | 64.09 | 0.13 | 0.070 | 60.22 | 0.13 | 0.074 | 441 | 34 |
| Skiathos | 66.02 | 0.14 | 0.076 | 58.71 | 0.14 | 0.078 | 429 | 13 |
| Mean | 65.97 | 0.14 | 0.074 | 64.51 | 0.14 | 0.080 | 471.5 | 49.5 |
| Loci/Groups | Source of Variation | d.f. | Variance Component | Total Variance (%) | Φ-Statistics (ΦST) | p-Value |
|---|---|---|---|---|---|---|
| AFLP loci | Among Populations | 3 | 0.835 | 2 | 0.016 | >0.001 |
| Within Populations | 116 | 52.891 | 98 | |||
| Total | 119 | 53.726 | 100 | |||
| MSAP all subepiloci | Among Populations | 3 | 4.862 | 9 | 0.095 | >0.001 |
| Within Populations | 116 | 46.514 | 91 | |||
| Total | 119 | 51.376 | 100 | |||
| MSAP m-subepiloci | Among Populations | 3 | 1.386 | 13 | 0.129 | >0.001 |
| Within Populations | 116 | 9.390 | 87 | |||
| Total | 119 | 10.776 | 100 | |||
| MSAP h-subepiloci | Among Populations | 3 | 2.540 | 11 | 0.114 | >0.001 |
| Within Populations | 116 | 19.677 | 89 | |||
| Total | 119 | 22.217 | 100 | |||
| MSAP n-subepiloci | Among Populations | 3 | 0.000 | 0 | 0.000 | <0.001 |
| Within Populations | 116 | 7.579 | 100 | |||
| Total | 119 | 7.579 | 100 |
| U Alleles | M Alleles | H Alleles | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Population | Ne | I | He | Ne | I | He | Ne | I | He |
| Kotixi | 1.174 | 0.178 | 0.110 | 1.139 | 0.167 | 0.096 | 1.097 | 0.134 | 0.072 |
| Skiathos | 1.186 | 0.193 | 0.118 | 1.155 | 0.190 | 0.109 | 1.098 | 0.129 | 0.071 |
| Strofilia | 1.185 | 0.202 | 0.122 | 1.182 | 0.206 | 0.122 | 1.094 | 0.127 | 0.069 |
| Schinias | 1.168 | 0.196 | 0.114 | 1.131 | 0.193 | 0.102 | 1.106 | 0.155 | 0.082 |
| Mean | 1.178 | 0.192 | 0.116 | 1.152 | 0.189 | 0.107 | 1.099 | 0.136 | 0.074 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avramidou, E.V.; Malliarou, E.; Korakaki, E.; Mantakas, G.; Kaoukis, K. Genetic and Epigenetic Diversity of Pinus pinea L.: Conservation Implications for Priority Populations in Greece. Genes 2025, 16, 361. https://doi.org/10.3390/genes16040361
Avramidou EV, Malliarou E, Korakaki E, Mantakas G, Kaoukis K. Genetic and Epigenetic Diversity of Pinus pinea L.: Conservation Implications for Priority Populations in Greece. Genes. 2025; 16(4):361. https://doi.org/10.3390/genes16040361
Chicago/Turabian StyleAvramidou, Evangelia V., Ermioni Malliarou, Evangelia Korakaki, George Mantakas, and Konstantinos Kaoukis. 2025. "Genetic and Epigenetic Diversity of Pinus pinea L.: Conservation Implications for Priority Populations in Greece" Genes 16, no. 4: 361. https://doi.org/10.3390/genes16040361
APA StyleAvramidou, E. V., Malliarou, E., Korakaki, E., Mantakas, G., & Kaoukis, K. (2025). Genetic and Epigenetic Diversity of Pinus pinea L.: Conservation Implications for Priority Populations in Greece. Genes, 16(4), 361. https://doi.org/10.3390/genes16040361