
Genome-Wide In Silico Analysis of Leucine-Rich Repeat R-Genes in Perilla citriodora: Classification and Expression Insights

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of P. citriodora ‘Jeju17’ NBS-LRR Genes and Classification
2.2. Chromosomal Localization, Gene Density, and Phylogenetic Analysis of NBS-LRR Genes
2.3. NLR Gene Synteny and Duplication Investigation
2.4. Differentially Expressed NLR Gene Analysis
3. Results
3.1. Identification and Classification of ‘Jeju17’ R-Genes
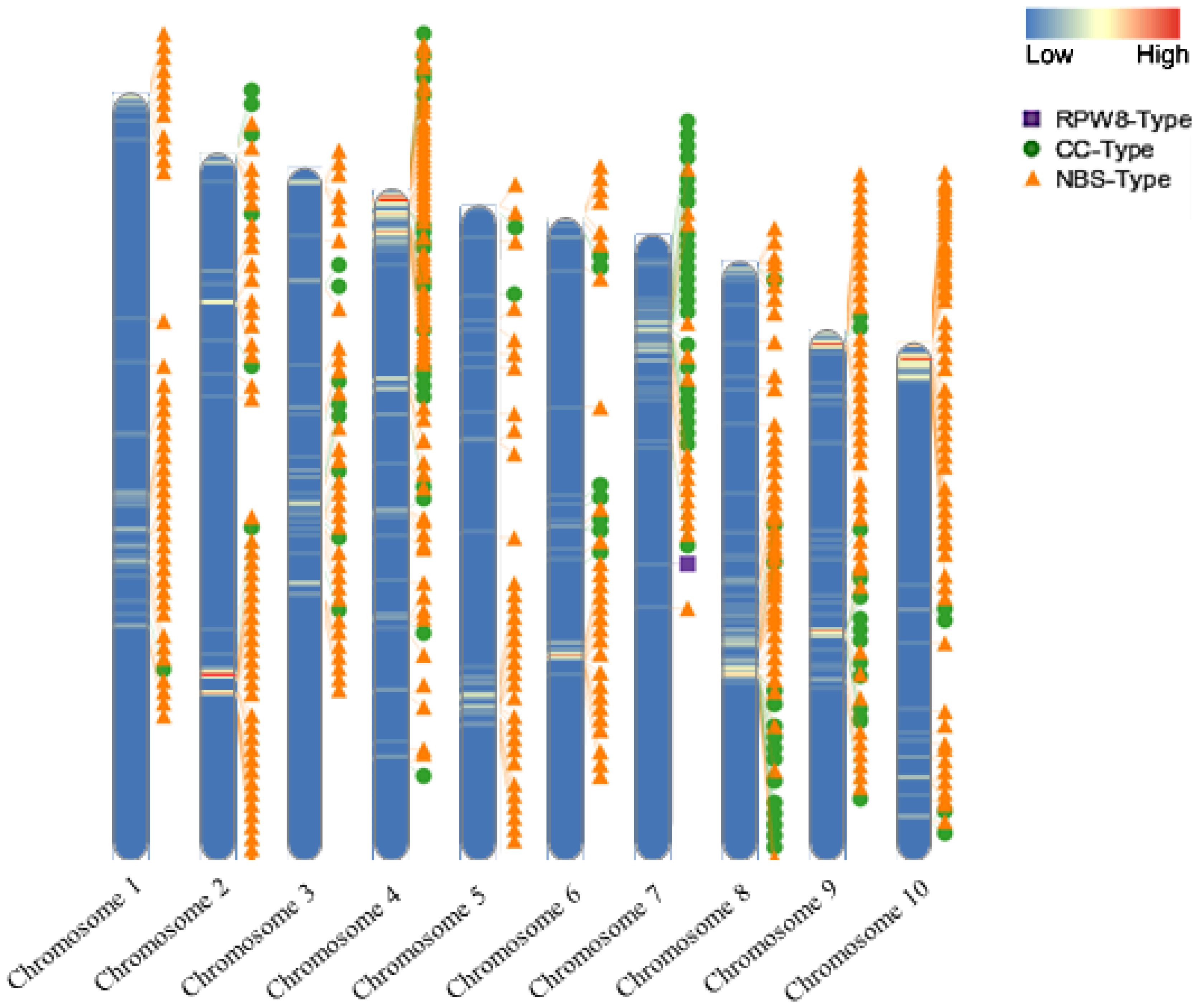
3.2. NLR Gene Distribution and Phylogenetic Tree Construction in the ‘Jeju17’ Genome
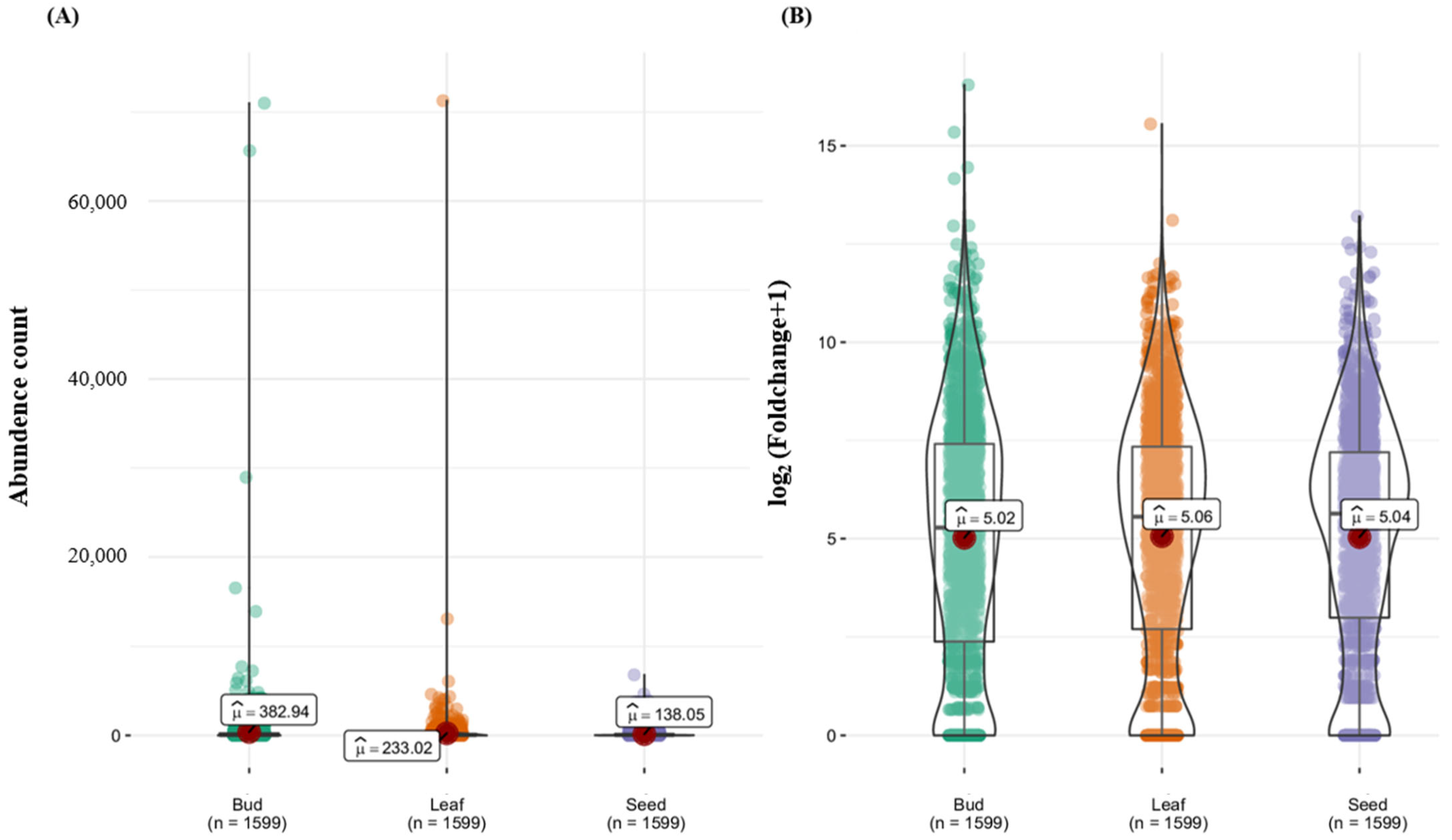
3.3. Expression Analysis of R-Genes in Tissues (Bud, Leaf, and Seed)
4. Discussion
4.1. Diversity and Evolution of R-Genes in ‘Jeju17’
4.2. Chromosomal Distribution and Defense Mechanisms of NBS-LRR Genes
4.3. Functional Importance and Uniqueness of RPW8 Gene
4.4. Evolutionary Conservation and Functional Roles of R-Gene Clusters
4.5. Breeding Applications of R-Gene Clusters
4.6. Tissue-Specific Expression and Functional Roles of R-Genes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mun, J.H.; Yu, H.J.; Park, S.; Park, B.S. Genome-wide identification of NBS-encoding resistance genes in Brassica rapa. Mol. Genet. Genom. 2009, 282, 617–631. [Google Scholar] [CrossRef]
- Seo, E.; Kim, S.; Yeom, S.I.; Choi, D. Genome-Wide Comparative Analyses Reveal the Dynamic Evolution of Nucleotide-Binding Leucine-Rich Repeat Gene Family among Solanaceae Plants. Front. Plant. Sci. 2016, 7, 1205. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Shao, Z.Q.; Wang, Q.; Hang, Y.Y.; Xue, J.Y.; Wang, B.; Chen, J.Q. Uncovering the dynamic evolution of nucleotide-binding site-leucine-rich repeat (NBS-LRR) genes in Brassicaceae. J. Integr. Plant Biol. 2016, 58, 165–177. [Google Scholar] [CrossRef]
- Moon, J.Y.; Park, J.M. Cross-Talk in Viral Defense Signaling in Plants. Front. Microbiol. 2016, 7, 2068. [Google Scholar] [CrossRef]
- Dangl, J.L.; Jones, J.D.G. Plant pathogens and integrated defence responses to infection. Nature 2001, 411, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Meyers, B.C.; Kozik, A.; Griego, A.; Kuang, H.; Michelmore, R.W. Genome-Wide Analysis of NBS-LRR–Encoding Genes in Arabidopsis. Plant Cell 2003, 15, 809–834. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Chen, M.; Sun, L.; Wang, Y.; Yin, J.; Liu, J.; Sun, X.Q.; Hang, Y.-Y. Genome-Wide Identification and Evolutionary Analysis of NBS-LRR Genes from Dioscorea rotundata. Front. Genet. 2020, 11, 484. [Google Scholar] [CrossRef]
- McHale, L.; Tan, X.; Koehl, P.; Michelmore, R.W. Plant NBS-LRR proteins: Adaptable guards. Genome Biol. 2006, 7, 212. [Google Scholar] [CrossRef]
- Arya, P.; Kumar, G.; Acharya, V.; Singh, A.K. Genome-Wide Identification and Expression Analysis of NBS-Encoding Genes in Malus x domestica and Expansion of NBS Genes Family in Rosaceae. PLoS ONE 2014, 9, e107987. [Google Scholar] [CrossRef]
- Dubey, N.; Singh, K. Role of NBS-LRR Proteins in Plant Defense. In Molecular Aspects of Plant-Pathogen Interaction; Singh, A., Singh, I.K., Eds.; Springer Singapore: Singapore, 2018; pp. 115–138. ISBN 978-981-10-7370-0. [Google Scholar]
- Zhang, Y.; Goritschnig, S.; Dong, X.; Li, X. A Gain-of-Function Mutation in a Plant Disease Resistance Gene Leads to Constitutive Activation of Downstream Signal Transduction Pathways in suppressor of npr1-1, constitutive 1. Plant Cell 2003, 15, 2636–2646. [Google Scholar] [CrossRef]
- Zhou, T.; Wang, Y.; Chen, J.Q.; Araki, H.; Jing, Z.; Jiang, K.; Shen, J.; Tian, D. Genome-wide identification of NBS genes in japonica rice reveals significant expansion of divergent non-TIR NBS-LRR genes. Mol. Genet. Genom. 2004, 271, 402–415. [Google Scholar] [CrossRef]
- Dai, L.Y.; Liu, X.L.; Xiao, Y.H.; Wang, G.L. Recent Advances in Cloning and Characterization of Disease Resistance Genes in Rice. J. Integr. Plant Biol. 2007, 49, 112–119. [Google Scholar] [CrossRef]
- Ashikawa, I.; Hayashi, N.; Yamane, H.; Kanamori, H.; Wu, J.; Matsumoto, T.; Ono, K.; Yano, M. Two Adjacent Nucleotide-Binding Site–Leucine-Rich Repeat Class Genes Are Required to Confer Pikm-Specific Rice Blast Resistance. Genetics 2008, 180, 2267–2276. [Google Scholar] [CrossRef]
- Risk, J.M.; Selter, L.L.; Krattinger, S.G.; Viccars, L.A.; Richardson, T.M.; Buesing, G.; Herren, G.; Lagudah, E.S.; Keller, B. Functional variability of the Lr34 durable resistance gene in transgenic wheat. Plant Biotechnol. J. 2012, 10, 477–487. [Google Scholar] [CrossRef]
- Feuillet, C.; Travella, S.; Stein, N.; Albar, L.; Nublat, A.; Keller, B. Map-based isolation of the leaf rust disease resistance gene Lr10 from the hexaploid wheat (Triticum aestivum L.) genome. Proc. Natl. Acad. Sci. USA 2003, 100, 15253–15258. [Google Scholar] [CrossRef]
- Jupe, F.; Pritchard, L.; Etherington, G.J.; MacKenzie, K.; Cock, P.J.A.; Wright, F.; Sharma, S.K.; Bolser, D.; Bryan, G.J.; Jones, J.D.G.; et al. Identification and localisation of the NB-LRR gene family within the potato genome. BMC Genom. 2012, 13, 75. [Google Scholar] [CrossRef]
- Wei, H.; Li, W.; Sun, X.; Zhu, S.; Zhu, J. Systematic Analysis and Comparison of Nucleotide-Binding Site Disease Resistance Genes in a Diploid Cotton Gossypium raimondii. PLoS ONE 2013, 8, e68435. [Google Scholar] [CrossRef]
- Neupane, S.; Andersen, E.J.; Neupane, A.; Nepal, M.P. Genome-Wide Identification of NBS-Encoding Resistance Genes in Sunflower (Helianthus annuus L.). Genes 2018, 9, 384. [Google Scholar] [CrossRef] [PubMed]
- Vining, K.J.; Johnson, S.R.; Ahkami, A.; Lange, I.; Parrish, A.N.; Trapp, S.C.; Croteau, R.B.; Straub, S.C.K.; Pandelova, I.; Lange, B.M. Draft Genome Sequence of Mentha longifolia and Development of Resources for Mint Cultivar Improvement. Mol. Plant 2017, 10, 323–339. [Google Scholar] [CrossRef]
- Kourelis, J.; van der Hoorn, R.A.L. Defended to the Nines: 25 Years of Resistance Gene Cloning Identifies Nine Mechanisms for R Protein Function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef]
- Marone, D.; Russo, M.; Laidò, G.; De Leonardis, A.; Mastrangelo, A. Plant nucleotide binding site–leucine-rich repeat (NBS-LRR) genes: Active guardians in host defense responses. Int. J. Mol. Sci. 2013, 14, 7302–7326. [Google Scholar] [CrossRef]
- Luo, S.; Zhang, Y.; Hu, Q.; Chen, J.; Li, K.; Lu, C.; Liu, J. Dynamic nucleotide-binding site and leucine-rich repeat-encoding genes in the grass family. Plant Physiol. 2012, 159, 197–210. [Google Scholar] [CrossRef]
- Shivaprasad, P.; Chen, H.; Patel, K.; Bond, D.; Santos, B.; Baulcombe, D. A microRNA superfamily regulates nucleotide binding site–leucine-rich repeats and other mRNAs. Plant Cell 2012, 24, 859–874. [Google Scholar] [CrossRef] [PubMed]
- Warren, R.F.; Henk, A.; Mowery, P.; Holub, E.; Innes, R.W. A Mutation within the Leucine-Rich Repeat Domain of the Arabidopsis Disease Resistance Gene RPS5 Partially Suppresses Multiple Bacterial and Downy Mildew Resistance Genes. Plant Cell 1998, 10, 1439–1452. [Google Scholar] [CrossRef]
- Dodds, P.N.; Lawrence, G.J.; Ellis, J.G. Six Amino Acid Changes Confined to the Leucine-Rich Repeat β-Strand/β-Turn Motif Determine the Difference between the P and P2 Rust Resistance Specificities in Flax. Plant Cell 2001, 13, 163–178. [Google Scholar] [CrossRef]
- van Ooijen, G.; Mayr, G.; Kasiem, M.M.A.; Albrecht, M.; Cornelissen, B.J.C.; Takken, F.L.W. Structure–function analysis of the NB-ARC domain of plant disease resistance proteins. J. Exp. Bot. 2008, 59, 1383–1397. [Google Scholar] [CrossRef]
- DeYoung, B.J.; Innes, R.W. Plant NBS-LRR proteins in pathogen sensing and host defense. Nat. Immunol. 2006, 7, 1243–1249. [Google Scholar] [CrossRef]
- Meyers, B.C.; Dickerman, A.W.; Michelmore, R.W.; Sivaramakrishnan, S.; Sobral, B.W.; Young, N.D. Plant disease resistance genes encode members of an ancient and diverse protein family within the nucleotide-binding superfamily. Plant J. 1999, 20, 317–332. [Google Scholar] [CrossRef]
- Xiao, S.; Ellwood, S.; Calis, O.; Patrick, E.; Li, T.; Coleman, M.; Turner, J.G. Broad-spectrum mildew resistance in Arabidopsis thaliana mediated by RPW8. Science 2001, 291, 118–120. [Google Scholar] [CrossRef]
- Bonardi, V.; Tang, S.; Stallmann, A.; Roberts, M.; Cherkis, K.; Dangl, J.L. Expanded functions for a family of plant intracellular immune receptors beyond specific recognition of pathogen effectors. Proc. Natl. Acad. Sci. USA 2011, 108, 16463–16468. [Google Scholar] [CrossRef]
- Collier, S.M.; Hamel, L.P.; Moffett, P. Cell Death Mediated by the N-Terminal Domains of a Unique and Highly Conserved Class of NB-LRR Protein. Mol. Plant Microbe Interact. 2011, 24, 918–931. [Google Scholar] [CrossRef]
- Lozano, R.; Ponce, O.; Ramirez, M.; Mostajo, N.; Orjeda, G. Genome-Wide Identification and Mapping of NBS-Encoding Resistance Genes in Solanum tuberosum Group Phureja. PLoS ONE 2012, 7, e34775. [Google Scholar] [CrossRef]
- Shao, Z.Q.; Xue, J.Y.; Wu, P.; Zhang, Y.M.; Wu, Y.; Hang, Y.Y.; Wang, B.; Chen, J.Q. Large-Scale Analyses of Angiosperm Nucleotide-Binding Site-Leucine-Rich Repeat Genes Reveal Three Anciently Diverged Classes with Distinct Evolutionary Patterns. Plant Physiol. 2016, 170, 2095–2109. [Google Scholar] [CrossRef]
- Bae, S.H.; Lee, M.H.; Lee, J.H.; Yu, Y.; Lee, J.; Kim, T.H. The Genome of the Korean Island-Originated Perilla citriodora ‘Jeju17’ Sheds Light on Its Environmental Adaptation and Fatty Acid and Lipid Production Pathways. Genes 2023, 14, 1898. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2018, 47, D427–D432. [Google Scholar] [CrossRef] [PubMed]
- Steuernagel, B.; Witek, K.; Krattinger, S.G.; Ramirez-Gonzalez, R.H.; Schoonbeek, H.-j.; Yu, G.; Baggs, E.; Witek, A.I.; Yadav, I.; Krasileva, K.V.; et al. The NLR-Annotator Tool Enables Annotation of the Intracellular Immune Receptor Repertoire. Plant Physiol. 2020, 183, 468–482. [Google Scholar] [CrossRef]
- Nepal, M.P.; Andersen, E.J.; Neupane, S.; Benson, B.V. Comparative Genomics of Non-TNL Disease Resistance Genes from Six Plant Species. Genes 2017, 8, 249. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Lv, D.; Ge, Y.; Shi, J.; Weijers, D.; Yu, G.; Chen, J. RIdeogram: Drawing SVG graphics to visualize and map genome-wide data on the idiograms. PeerJ. Comput. Sci. 2020, 6, e251. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.i.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.-H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- The R Foundation. R: A Language and Environment for Statistical Computing; R Foundation for Statistical: Vienna, Austria, 2013; Available online: www.R-project.org/ (accessed on 24 April 2024).
- Wang, L.; Yu, S.; Tong, C.; Zhao, Y.; Liu, Y.; Song, C.; Zhang, Y.; Zhang, X.; Wang, Y.; Hua, W.; et al. Genome sequencing of the high oil crop sesame provides insight into oil biosynthesis. Genome Biol. 2014, 15, R39. [Google Scholar] [CrossRef]
- Bettaieb, I.; Bouktila, D. Genome-wide analysis of NBS-encoding resistance genes in the Mediterranean olive tree (Olea europaea subsp. europaea var. europaea): Insights into their molecular diversity, evolution and function. Tree Genet. Genom. 2020, 16, 23. [Google Scholar] [CrossRef]
- Tarr, D.E.K.; Alexander, H.M. TIR-NBS-LRR genes are rare in monocots: Evidence from diverse monocot orders. BMC Res. Notes 2009, 2, 197. [Google Scholar] [CrossRef]
- Tian, Y.; Fan, L.; Thurau, T.; Jung, C.; Cai, D. The Absence of TIR-Type Resistance Gene Analogues in the Sugar Beet (Beta vulgaris L.) Genome. J. Mol. Evol. 2004, 58, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Peart, J.R.; Mestre, P.; Lu, R.; Malcuit, I.; Baulcombe, D.C. NRG1, a CC-NB-LRR Protein, together with N, a TIR-NB-LRR Protein, Mediates Resistance against Tobacco Mosaic Virus. Curr. Biol. 2005, 15, 968–973. [Google Scholar] [CrossRef] [PubMed]
- Qi, T.; Seong, K.; Thomazella, D.P.T.; Kim, J.R.; Pham, J.; Seo, E.; Cho, M.-J.; Schultink, A.; Staskawicz, B.J. NRG1 functions downstream of EDS1 to regulate TIR-NLR-mediated plant immunity in Nicotiana benthamiana. Proc. Natl. Acad. Sci. USA 2018, 115, E10979–E10987. [Google Scholar] [CrossRef]
- Song, X.; Li, C.; Liu, Z.; Zhou, R.; Shen, S.; Yu, T.; Li, N. Genome-Wide Analysis of the NBS-LRR Gene Family and SSR Molecular Markers Development in Solanaceae. Horticulturae 2024, 10, 1293. [Google Scholar] [CrossRef]
- Mondragón-Palomino, M.; Meyers, B.C.; Michelmore, R.W.; Gaut, B.S. Patterns of positive selection in the complete NBS-LRR gene family of Arabidopsis thaliana. Genome Res. 2002, 12, 1305–1315. [Google Scholar] [CrossRef]
- Zhou, T.; Wang, Y.; Chen, J.; Liu, Z. Evolutionary dynamics of NBS-LRR resistance genes in rice and their role in plant immunity. J. Plant Sci. 2019, 187, 67–76. [Google Scholar] [CrossRef]
- Kuang, H.; Woo, S.S.; Meyers, B.C.; Nevo, E.; Michelmore, R.W. Multiple genetic processes result in heterogeneous rates of evolution within the major cluster disease resistance genes in lettuce. Plant Cell 2004, 16, 2870–2884. [Google Scholar] [CrossRef]
- Xiao, S.; Calis, O.; Patrick, E.; Zhang, G.; Charoenwattana, P.; Muskett, P.; Turner, J. The atypical resistance gene, RPW8, recruits components of basal defence for powdery mildew resistance in Arabidopsis. Plant J. 2005, 42, 95–110. [Google Scholar] [CrossRef]
- Zhong, Y.; Cheng, Z.M. A unique RPW8-encoding class of genes that originated in early land plants and evolved through domain fission, fusion, and duplication. Sci. Rep. 2016, 6, 32923. [Google Scholar] [CrossRef]
- Barragan, C.A.; Wu, R.; Kim, S.T.; Xi, W.; Habring, A.; Hagmann, J.; Van de Weyer, A.L.; Zaidem, M.; Ho, W.W.H.; Wang, G.; et al. RPW8/HR repeats control NLR activation in Arabidopsis thaliana. PLoS Genet. 2019, 15, e1008313. [Google Scholar] [CrossRef]
- Zhong, Y.; Li, Y.; Huang, K.; Cheng, Z.-M. Species-specific duplications of NBS-encoding genes in Chinese chestnut (Castanea mollissima). Sci. Rep. 2015, 5, 16638. [Google Scholar] [CrossRef]
- Schulze, S.; Yu, L.; Hua, C.; Zhang, L.; Kolb, D.; Weber, H.; Nürnberger, T. The Arabidopsis TIR-NBS-LRR protein CSA1 guards BAK1-BIR3 homeostasis and mediates convergence of pattern-and effector-induced immune responses. Cell Host Microbe 2022, 30, 1717–1731.e6. [Google Scholar] [CrossRef]
- Neupane, S.; Ma, Q.; Mathew, F.M.; Varenhorst, A.J.; Andersen, E.J.; Nepal, M.P. Evolutionary Divergence of TNL Disease-Resistant Proteins in Soybean (Glycine max) and Common Bean (Phaseolus vulgaris). Biochem. Genet. 2018, 56, 397–422. [Google Scholar] [CrossRef]
- Friedman, A.R.; Baker, B.J. The evolution of resistance genes in multi-protein plant resistance systems. Curr. Opin. Genet. Dev. 2007, 17, 493–499. [Google Scholar] [CrossRef]
- Joshi, R.K.; Nayak, S. Perspectives of genomic diversification and molecular recombination towards R-gene evolution in plants. Physiol. Mol. Biol. Plants 2013, 19, 1–9. [Google Scholar] [CrossRef]
- Shao, Z.Q.; Zhang, Y.M.; Hang, Y.Y.; Xue, J.Y.; Zhou, G.C.; Wu, P.; Wu, X.Y.; Wu, X.Z.; Wang, Q.; Wang, B.; et al. Long-Term Evolution of Nucleotide-Binding Site-Leucine-Rich Repeat Genes: Understanding Gained from and beyond the Legume Family. Plant Physiol. 2014, 166, 217–234. [Google Scholar] [CrossRef]
- Mcintosh, R.A. Catalogue of gene symbols for wheat. Cereal Res. Commun. 1979, 2, 1299–1309. [Google Scholar]
- Mcintosh, R.A. Wheat Rusts: An Atlas of Resistance Genes; Csiro Publishing: Collingwood, Australia, 1995. [Google Scholar]
- Cloutier, S.; McCallum, B.; Loutre, C.; Banks, T.; Wicker, T.; Feuillet, C.; Jordan, M. Leaf rust resistance gene Lr1, isolated from bread wheat (Triticum aestivum L.) is a member of the large psr567 gene family. Plant Mol. Biol. 2007, 65, 93–106. [Google Scholar] [CrossRef]
- Richly, E.; Kurth, J.; Leister, D. Mode of amplification and reorganization of resistance genes during recent Arabidopsis thaliana evolution. Mol. Biol. Evol. 2002, 19, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Meyers, B.C.; Shen, K.A.; Rohani, P.; Gaut, B.S.; Michelmore, R.W. Receptor-like genes in the major resistance locus of lettuce are subject to divergent selection. Plant Cell 1998, 10, 1833–1846. [Google Scholar] [CrossRef]
- Liu, F.; Xu, X.; Zheng, H. Integrating genome-wide association studies with molecular breeding for stress resistance in crops. Trends Plant Sci. 2021, 26, 601–613. [Google Scholar] [CrossRef]
- Schneider, E.; Hunke, S. ATP-binding-cassette (ABC) transport systems: Functional and structural aspects of the ATP-hydrolyzing subunits/domains. FEMS Microbiol. Rev. 1998, 22, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, Y.; Hagiya, Y.; Adachi, T.; Hoshijima, K.; Kuo, M.T.; Ishikawa, T. MRP class of human ATP binding cassette (ABC) transporters: Historical background and new research directions. Xenobiotica 2008, 38, 833–862. [Google Scholar] [CrossRef]
- Crouzet, J.; Trombik, T.; Fraysse, Å.S.; Boutry, M. Organization and function of the plant pleiotropic drug resistance ABC transporter family. FEBS Lett. 2006, 580, 1123–1130. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Class | Type | Number of Genes |
|---|---|---|---|
| ‘Jeju17’ | 1 | CC-NB-ARC | 87 |
| 2 | CC-NB-ARC-LRR | 17 | |
| 3 | NB-ARC | 395 | |
| 4 | NB-ARC-LRR | 35 | |
| 5 | RPW8-NB-ARC | 1 | |
| Total | 535 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, S.-H.; Zoclanclounon, Y.A.B.; Park, G.-H.; Lee, J.-D.; Kim, T.-H. Genome-Wide In Silico Analysis of Leucine-Rich Repeat R-Genes in Perilla citriodora: Classification and Expression Insights. Genes 2025, 16, 200. https://doi.org/10.3390/genes16020200
Bae S-H, Zoclanclounon YAB, Park G-H, Lee J-D, Kim T-H. Genome-Wide In Silico Analysis of Leucine-Rich Repeat R-Genes in Perilla citriodora: Classification and Expression Insights. Genes. 2025; 16(2):200. https://doi.org/10.3390/genes16020200
Chicago/Turabian StyleBae, Seon-Hwa, Yedomon Ange Bovys Zoclanclounon, Gyu-Hwang Park, Jun-Dae Lee, and Tae-Ho Kim. 2025. "Genome-Wide In Silico Analysis of Leucine-Rich Repeat R-Genes in Perilla citriodora: Classification and Expression Insights" Genes 16, no. 2: 200. https://doi.org/10.3390/genes16020200
APA StyleBae, S.-H., Zoclanclounon, Y. A. B., Park, G.-H., Lee, J.-D., & Kim, T.-H. (2025). Genome-Wide In Silico Analysis of Leucine-Rich Repeat R-Genes in Perilla citriodora: Classification and Expression Insights. Genes, 16(2), 200. https://doi.org/10.3390/genes16020200