
Transcriptional Regulatory Network of the Embryonic Diapause Termination Process in Artemia

Abstract
1. Introduction
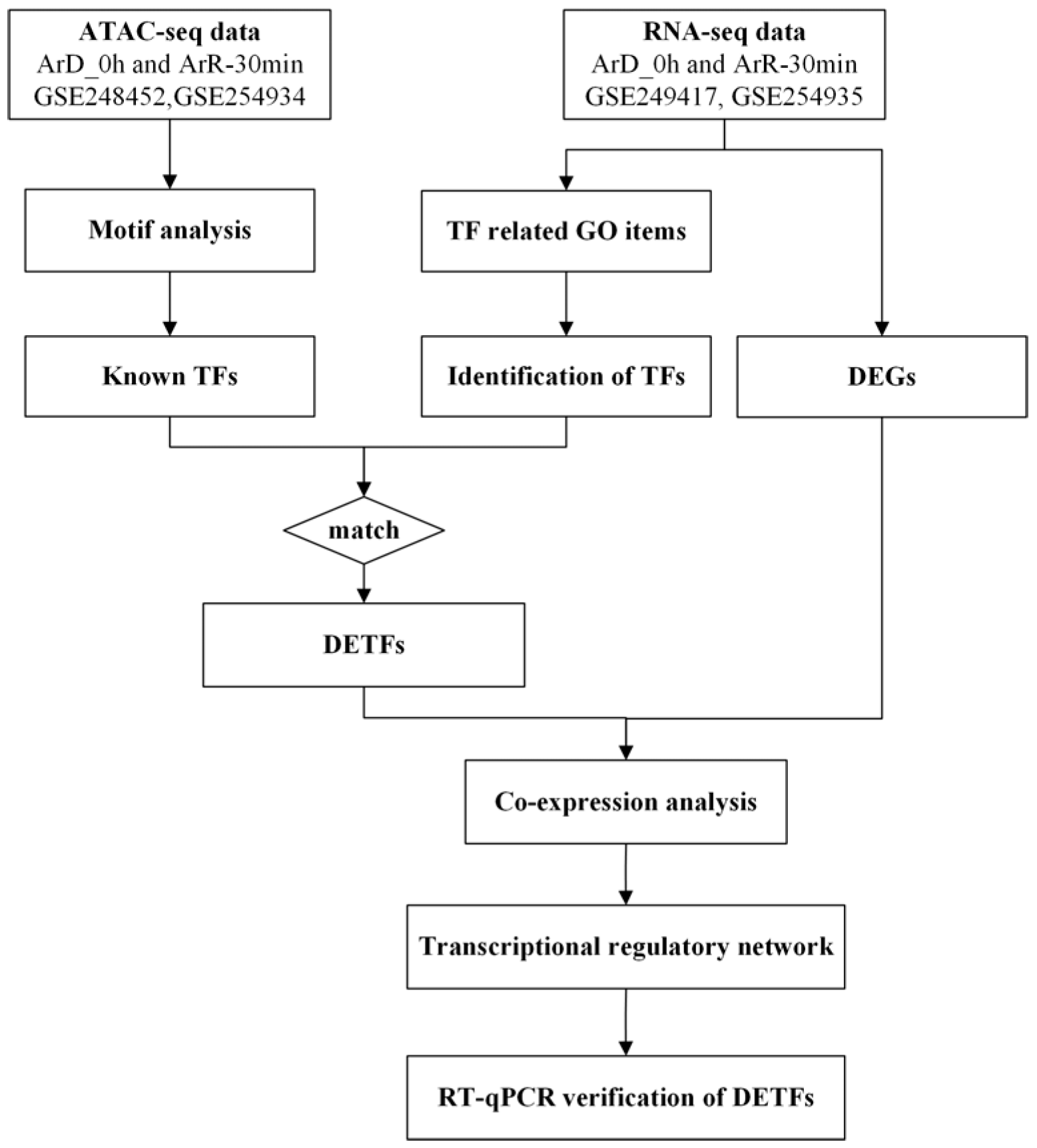
2. Materials and Methods
2.1. Design of the Experiments
2.2. Motif Analysis Based on ATAC-Seq
2.3. Identification of EDT Related TFs
2.4. Co-Expression Network Construction and GO Enrichment Analysis
2.5. Artemia Hatching and Culture
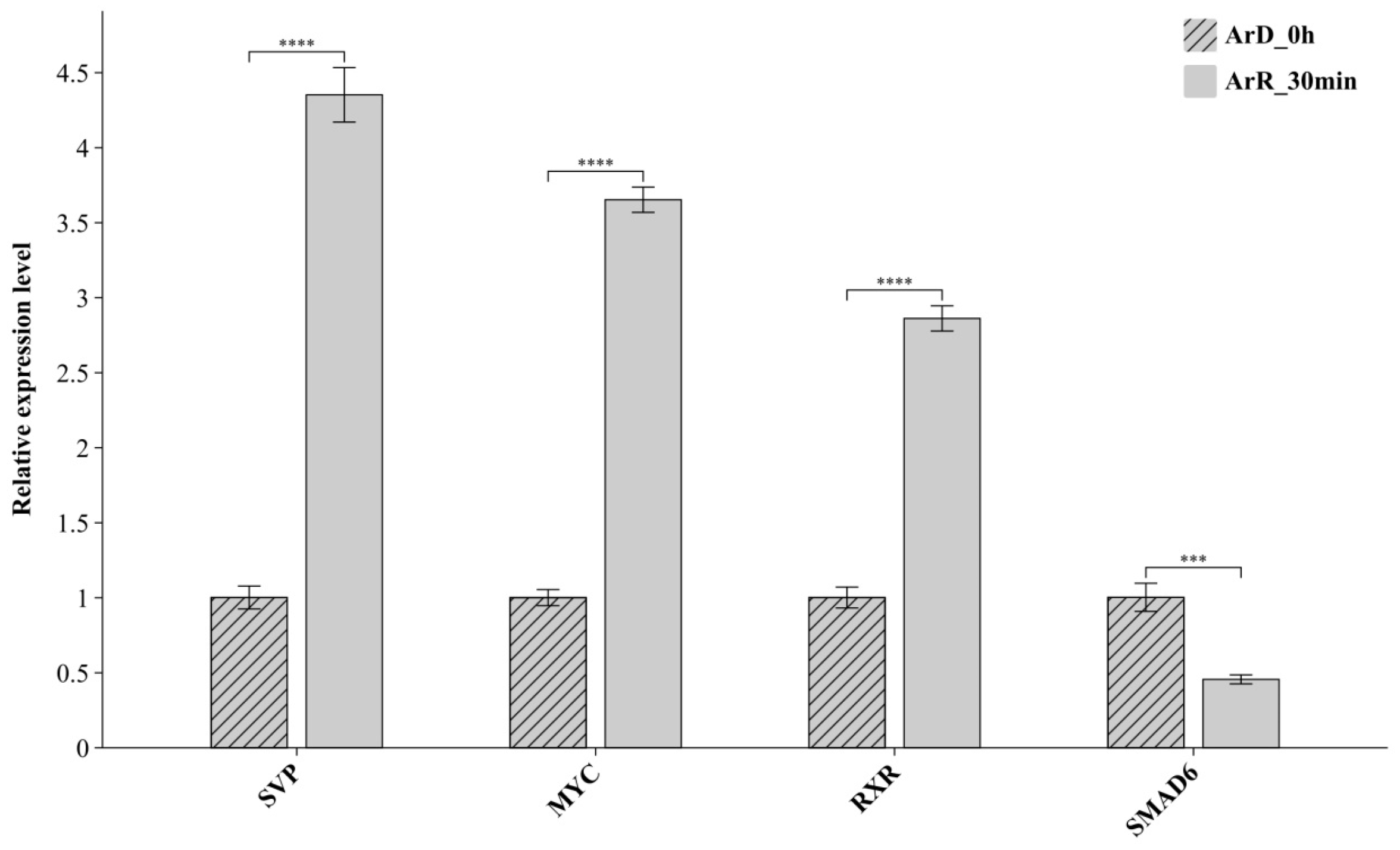
2.6. RT-qPCR Verification of DETF
2.7. Expression of the Target Genes from 0 h to 5 h After EDT
3. Results
3.1. Enriched Motif Analysis and Functional Analysis of ATAC Peaks in Diapause and Activated Cysts
3.2. Identification of EDT Related TFs
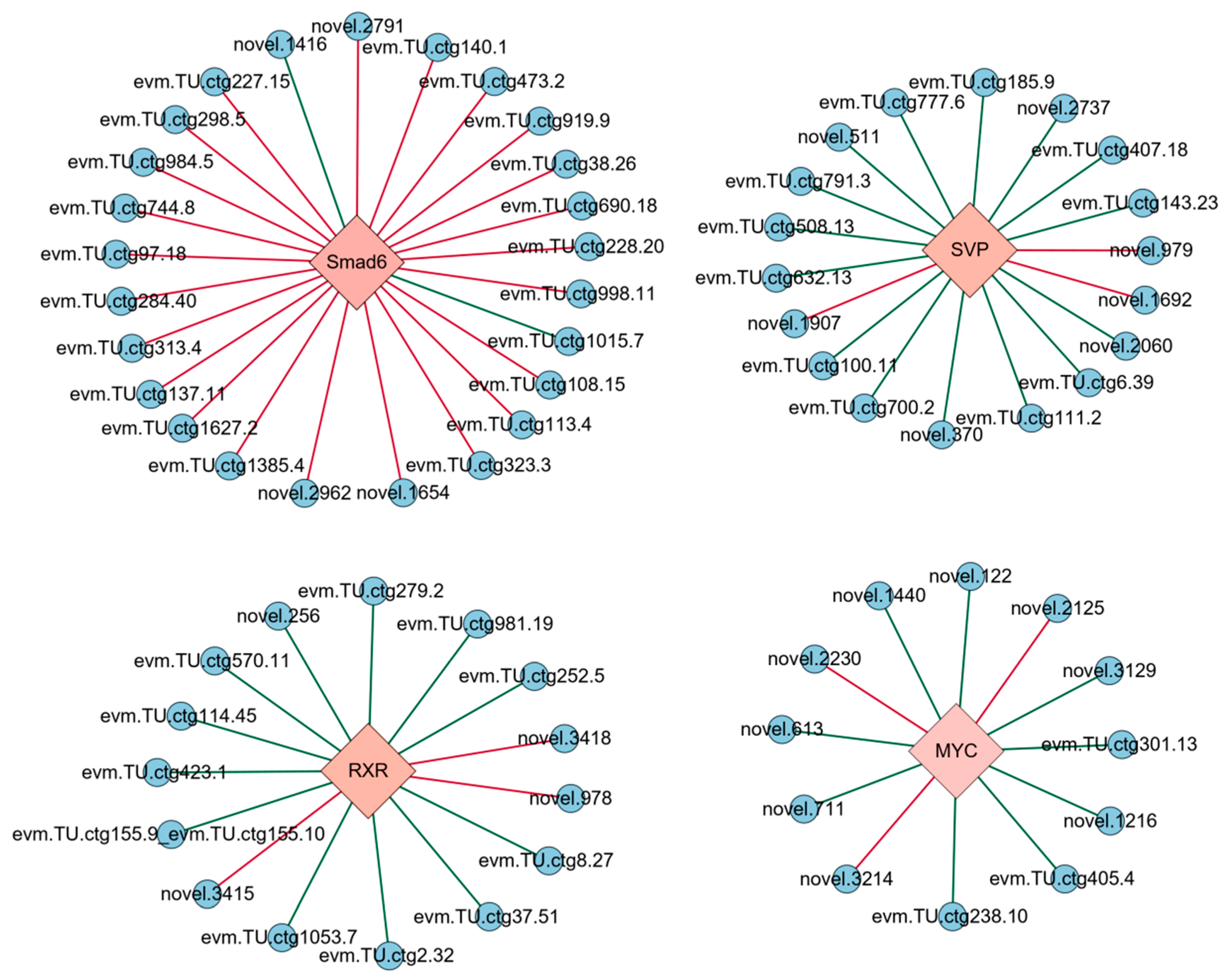
3.3. Transcriptional Regulatory Network of the EDT Process in Artemia
3.4. RT-qPCR Verification of DETFs
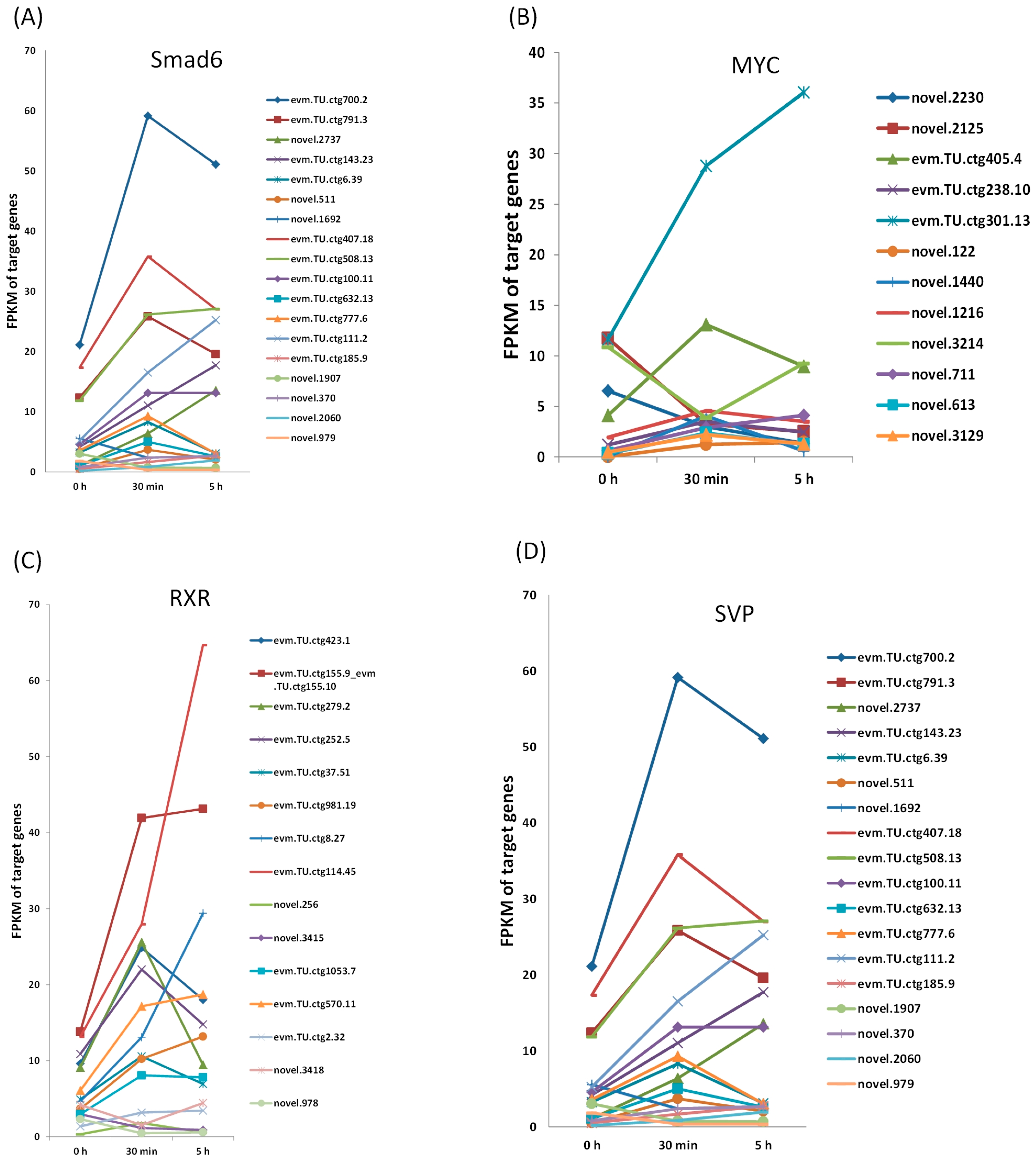
3.5. Expression of the Target Genes from 0 h to 5 h After EDT
4. Discussion
4.1. TFs in the EDT Process of Artemia
4.2. Function of the TFs in the EDT Process of Artemia
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Liu, Y.-L.; Zhao, Y.; Dai, Z.-M.; Chen, H.-M.; Yang, W.-J. Formation of Diapause Cyst Shell in Brine Shrimp, Artemia parthenogenetica, and Its Resistance Role in Environmental Stresses. J. Biol. Chem. 2009, 284, 16931–16938. [Google Scholar] [CrossRef]
- Qiu, Z.; MacRae, T.H. A Molecular Overview of Diapause in Embryos of the Crustacean, Artemia franciscana; Springer: Berlin/Heidelberg, Germany, 2010; Volume 21. [Google Scholar]
- MacRae, T.H. Stress tolerance during diapause and quiescence of the brine shrimp, Artemia. Cell Stress Chaperones 2016, 21, 9–18. [Google Scholar] [CrossRef]
- Fenelon, J.C.; Banerjee, A.; Murphy, B.D. Embryonic diapause: Development on hold. Int. J. Dev. Biol. 2014, 58, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Renfree, M.B.; Shaw, G. Diapause. Annu. Rev. Physiol. 2000, 62, 353–375. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.D. Under Arrest: The Embryo in Diapause. Dev. Cell 2020, 52, 139–140. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.; MacRae, T.H. The small heat shock protein p26 aids development of encysting Artemia embryos, prevents spontaneous diapause termination and protects against stress. PLoS ONE 2012, 7, e43723. [Google Scholar] [CrossRef]
- Qiu, Z.; Macrae, T.H. ArHsp21, a developmentally regulated small heat-shock protein synthesized in diapausing embryos of Artemia franciscana. Biochem. J. 2008, 411, 605–611. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.; Toxopeus, J.; MacRae, T.H. Functional differentiation of small heat shock proteins in diapause-destined Artemia embryos. FEBS J. 2013, 280, 4761–4772. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; MacRae, T.H. ArHsp22, a developmentally regulated small heat shock protein produced in diapause-destined Artemia embryos, is stress inducible in adults. FEBS J. 2008, 275, 3556–3566. [Google Scholar] [CrossRef]
- Rowarth, N.M.; MacRae, T.H. Post-diapause synthesis of ArHsp40-2, a type 2 J-domain protein from Artemia franciscana, is developmentally regulated and induced by stress. PLoS ONE 2018, 13, e0201477. [Google Scholar] [CrossRef]
- Ye, H.L.; Li, D.R.; Yang, J.S.; Chen, D.F.; De Vos, S.; Vuylsteke, M.; Sorgeloos, P.; Van Stappen, G.; Bossier, P.; Nagasawa, H.; et al. Molecular characterization and functional analyses of a diapause hormone receptor-like gene in parthenogenetic Artemia. Peptides 2017, 90, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yao, F.; Zhang, H.; Li, N.; Zou, X.; Sui, L.; Hou, L. The Potential Roles of the Apoptosis-Related Protein PDRG1 in Diapause Embryo Restarting of Artemia sinica. Int. J. Mol. Sci. 2018, 19, 126. [Google Scholar] [CrossRef]
- Zhang, S.; Yao, F.; Jing, T.; Zhang, M.; Zhao, W.; Zou, X.; Sui, L.; Hou, L. Cloning, expression pattern, and potential role of apoptosis inhibitor 5 in the termination of embryonic diapause and early embryo development of Artemia sinica. Gene 2017, 628, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.H.; Li, A.Q.; Feng, J.Y.; Ding, Y.F.; Ye, S.; Yang, J.S.; Yang, W.J. DEK terminates diapause by activation of quiescent cells in the crustacean Artemia. Biochem. J. 2019, 476, 1753–1769. [Google Scholar] [CrossRef]
- Wang, X.; Yao, F.; Liang, X.; Zhu, X.; Zheng, R.; Jia, B.; Hou, L.; Zou, X. Cloning and expression of retinoblastoma-binding protein 4 gene in embryo diapause termination and in response to salinity stress from brine shrimp Artemia sinica. Gene 2016, 591, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Yao, F.; Chu, B.; Li, X.; Liu, Y.; Wu, Y.; Mei, Y.; Wang, P.; Hou, L.; Zou, X. Identification of the glycerol kinase gene and its role in diapause embryo restart and early embryo development of Artemia sinica. Gene 2014, 537, 51–62. [Google Scholar] [CrossRef]
- Hussein, A.M.; Balachandar, N.; Mathieu, J.; Ruohola-Baker, H. Molecular Regulators of Embryonic Diapause and Cancer Diapause-like State. Cells 2022, 11, 2929. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ran, L.-L.; Li, Y.; Liu, Y.-H. Comparative proteomics provides insights into diapause program of Bactrocera minax (Diptera: Tephritidae). PLoS ONE 2020, 15, e0244493. [Google Scholar] [CrossRef]
- Ouellet, J.; Li, S.; Roy, R. Notch signalling is required for both dauer maintenance and recovery in C. elegans. Development 2008, 135, 2583–2592. [Google Scholar] [CrossRef]
- Lin, C.; Jia, S.-N.; Yang, F.; Jia, W.-H.; Yu, X.-J.; Yang, J.-S.; Yang, W.-J. The transcription factor p8 regulates autophagy during diapause embryo formation in Artemia parthenogenetica. Cell Stress Chaperones 2016, 21, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.-J.; Dong, X.-L.; Kang, K.; Chen, H.; Dai, X.-Y.; Wu, G.-A.; Zheng, L.; Yu, Y.; Zhai, Y.-F. FoxO Transcription Factor Regulate Hormone Mediated Signaling on Nymphal Diapause. Front. Physiol. 2018, 9, 1654. [Google Scholar] [CrossRef]
- Chang, V.; Meuti, M.E. Circadian transcription factors differentially regulate features of the adult overwintering diapause in the Northern house mosquito, Culex pipiens. Insect Biochem. Mol. Biol. 2020, 121, 103365. [Google Scholar] [CrossRef] [PubMed]
- Acebal, M.C.; Hansen, B.W.; Jorgensen, T.S.; Dalgaard, L.T. Analysis of the transcriptional pathways associated with the induction of quiescent embryonic arrest in the calanoid copepod Acartia tonsa. Dev. Biol. 2023, 504, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Roncalli, V.; Cieslak, M.C.; Castelfranco, A.M.; Hopcroft, R.R.; Hartline, D.K.; Lenz, P.H. Correction to: Post-diapause transcriptomic restarts: Insight from a high-latitude copepod. BMC Genom. 2021, 22, 659. [Google Scholar] [CrossRef] [PubMed]
- Lenz, P.H.; Roncalli, V.; Cieslak, M.C.; Tarrant, A.M.; Castelfranco, A.M.; Hartline, D.K. Diapause vs. reproductive programs: Transcriptional phenotypes in a keystone copepod. Commun. Biol. 2021, 4, 426. [Google Scholar] [CrossRef] [PubMed]
- Hao, T.; Zhang, M.; Song, Z.; Gou, Y.; Wang, B.; Sun, J. Reconstruction of Eriocheir sinensis Protein–Protein Interaction Network Based on DGO-SVM Method. Curr. Issues Mol. Biol. 2024, 46, 7353–7372. [Google Scholar] [CrossRef]
- Wang, W.; Meng, B.; Chen, W.; Ge, X.; Liu, S.; Yu, J. A proteomic study on postdiapaused embryonic development of brine shrimp (Artemia franciscana). Proteomics 2007, 7, 3580–3591. [Google Scholar] [CrossRef]
- Li, A.; Song, Z.; Zhang, M.; Duan, H.; Sui, L.; Wang, B.; Hao, T. Integrating ATAC-seq and RNA-seq reveals the signal regulation involved in the Artemia embryonic reactivation process. Gene 2024, 15, 1083. [Google Scholar] [CrossRef] [PubMed]
- Saito, R.; Smoot, M.E.; Ono, K.; Ruscheinski, J.; Wang, P.-L.; Lotia, S.; Pico, A.R.; Bader, G.D.; Ideker, T. A travel guide to Cytoscape plugins. Nat. Methods 2012, 9, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Ge, X.; Wang, W.; Yu, J.; Hu, S. A gene catalogue for post-diapause development of an anhydrobiotic arthropod Artemia franciscana. BMC Genom. 2009, 10, 52. [Google Scholar] [CrossRef]
- Tan, A.; Palli, S.R. Identification and characterization of nuclear receptors from the red flour beetle, Tribolium castaneum. Insect Biochem. Mol. Biol. 2008, 38, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Miura, K.; Chen, L.; Raikhel, A.S. Cyclicity of mosquito vitellogenic ecdysteroid-mediated signaling is modulated by alternative dimerization of the RXR homologue Ultraspiracle. Proc. Natl. Acad. Sci. USA 2003, 100, 544–549. [Google Scholar] [CrossRef]
- Borras-Castells, F.; Nieva, C.; Maestro, J.L.; Maestro, O.; Belles, X.; Martín, D. Juvenile hormone biosynthesis in adult Blattella germanica requires nuclear receptors seven-up and FTZ-F1. Sci. Rep. 2017, 7, 40234. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.; Etebari, K.; Nouzova, M.; Noriega, F.G.; Asgari, S. Differential gene expression and microRNA profile in corpora allata-corpora cardiaca of Aedes aegypti mosquitoes with weak juvenile hormone signalling. BMC Genom. 2024, 25, 113. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, P.M.; Durica, D.; Tracy, W. RXR isoforms and endogenous retinoids in the fiddler crab, Uca pugilator. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 2008, 151, 602–614. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.A.J.; Brierley, A.S.; Pond, D.W.; Smith, V.J. Changes in seasonal expression patterns of ecdysone receptor, retinoid X receptor and an A-type allatostatin in the copepod, Calanus finmarchicus, in a sea loch environment: An investigation of possible mediators of diapause. Gen. Comp. Endocrinol. 2013, 189, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Girgrah, R.A.-R. SMAD Transcription Factor Expression in Anoxia-Tolerant Trachemys Scripta Elegans and Dehydration-Tolerant Xenopus Laevis: A Comparative Study. Master’s Thesis, Carleton University, Ottawa, ON, Canada, 2016. [Google Scholar]
- Park, D.; Estevez, A.; Riddle, D.L. Antagonistic Smad transcription factors control the dauer/non-dauer switch in C. elegans. Development 2010, 137, 477–485. [Google Scholar] [CrossRef]
- Eubelen, M.; Bostaille, N.; Cabochette, P.; Gauquier, A.; Tebabi, P.; Dumitru, A.C.; Koehler, M.; Gut, P.; Alsteens, D.; Stainier, D.Y.R.; et al. A molecular mechanism for Wnt ligand-specific signaling. Science 2018, 361, eaat1178. [Google Scholar] [CrossRef]
- Kim, S.; Chung, S.; Yoon, J.; Choi, K.-W.; Yim, J. Ectopic expression of Tollo/Toll-8 antagonizes Dpp signaling and induces cell sorting in the Drosophila wing. Genesis 2006, 44, 541–549. [Google Scholar] [CrossRef]
- Jefferson, A.B.; Majerus, P.W. Properties of type II inositol polyphosphate 5-phosphatase. J. Biol. Chem. 1995, 270, 9370–9377. [Google Scholar] [CrossRef] [PubMed]
- Raza, Q.; Choi, J.Y.; Li, Y.; O’Dowd, R.M.; Watkins, S.C.; Chikina, M.; Hong, Y.; Clark, N.L.; Kwiatkowski, A.V. Evolutionary rate covariation analysis of E-cadherin identifies Raskol as a regulator of cell adhesion and actin dynamics in Drosophila. PLoS Genet. 2019, 15, e1007720. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Number of Enriched Motifs | Number of Known TFs |
|---|---|---|
| ArD_0h_1 | 33 | 154 |
| ArD_0h_2 | 39 | 276 |
| ArD_0h_3 | 36 | 248 |
| ArR_30min_1 | 35 | 456 |
| ArR_30min_2 | 42 | 317 |
| ArR_30min_3 | 44 | 404 |
| GO Items | Description | Enriched DEGs | Matched TFs |
|---|---|---|---|
| GO:0000988 | transcription factor activity, protein binding | 9 | 1 |
| GO:0000989 | transcription factor activity, transcription factor binding | 9 | 1 |
| GO:0001071 | nucleic acid binding transcription factor activity | 37 | 10 |
| GO:0003700 | transcription factor activity, sequence-specific DNA binding | 37 | 10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, B.; He, Z.; Zhang, M.; Zhang, R.; Song, Z.; Li, A.; Hao, T. Transcriptional Regulatory Network of the Embryonic Diapause Termination Process in Artemia. Genes 2025, 16, 175. https://doi.org/10.3390/genes16020175
Wang B, He Z, Zhang M, Zhang R, Song Z, Li A, Hao T. Transcriptional Regulatory Network of the Embryonic Diapause Termination Process in Artemia. Genes. 2025; 16(2):175. https://doi.org/10.3390/genes16020175
Chicago/Turabian StyleWang, Bin, Zhen He, Mingzhi Zhang, Ruiqi Zhang, Zhentao Song, Anqi Li, and Tong Hao. 2025. "Transcriptional Regulatory Network of the Embryonic Diapause Termination Process in Artemia" Genes 16, no. 2: 175. https://doi.org/10.3390/genes16020175
APA StyleWang, B., He, Z., Zhang, M., Zhang, R., Song, Z., Li, A., & Hao, T. (2025). Transcriptional Regulatory Network of the Embryonic Diapause Termination Process in Artemia. Genes, 16(2), 175. https://doi.org/10.3390/genes16020175