
Novel Genome-Engineered H Alleles Differentially Affect Lateral Inhibition and Cell Dichotomy Processes during Bristle Organ Development

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fly Work
2.2. Immunohistology
2.3. Behavioral Assays
2.4. Yeast 2-Hybrid Analysis
2.5. Western Blots
2.6. qRT-PCR
2.7. Statistics
3. Results
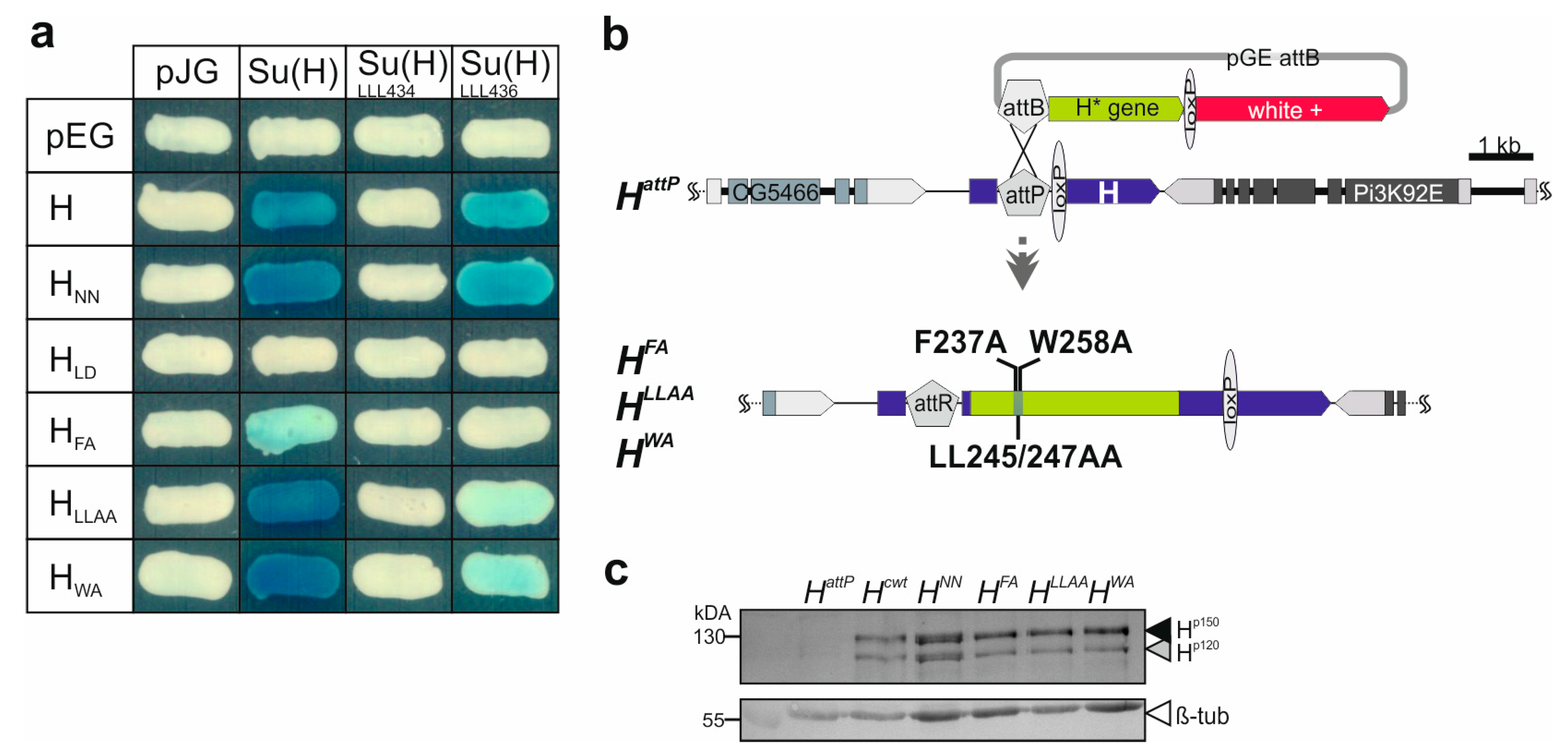
3.1. Repressor Complex Formation Addressed by Protein–Protein Interaction Studies and the Generation of Novel H Replacement Mutants Specifically Affecting Su(H) Binding
3.2. Phenotypic Analysis of the Novel H Alleles
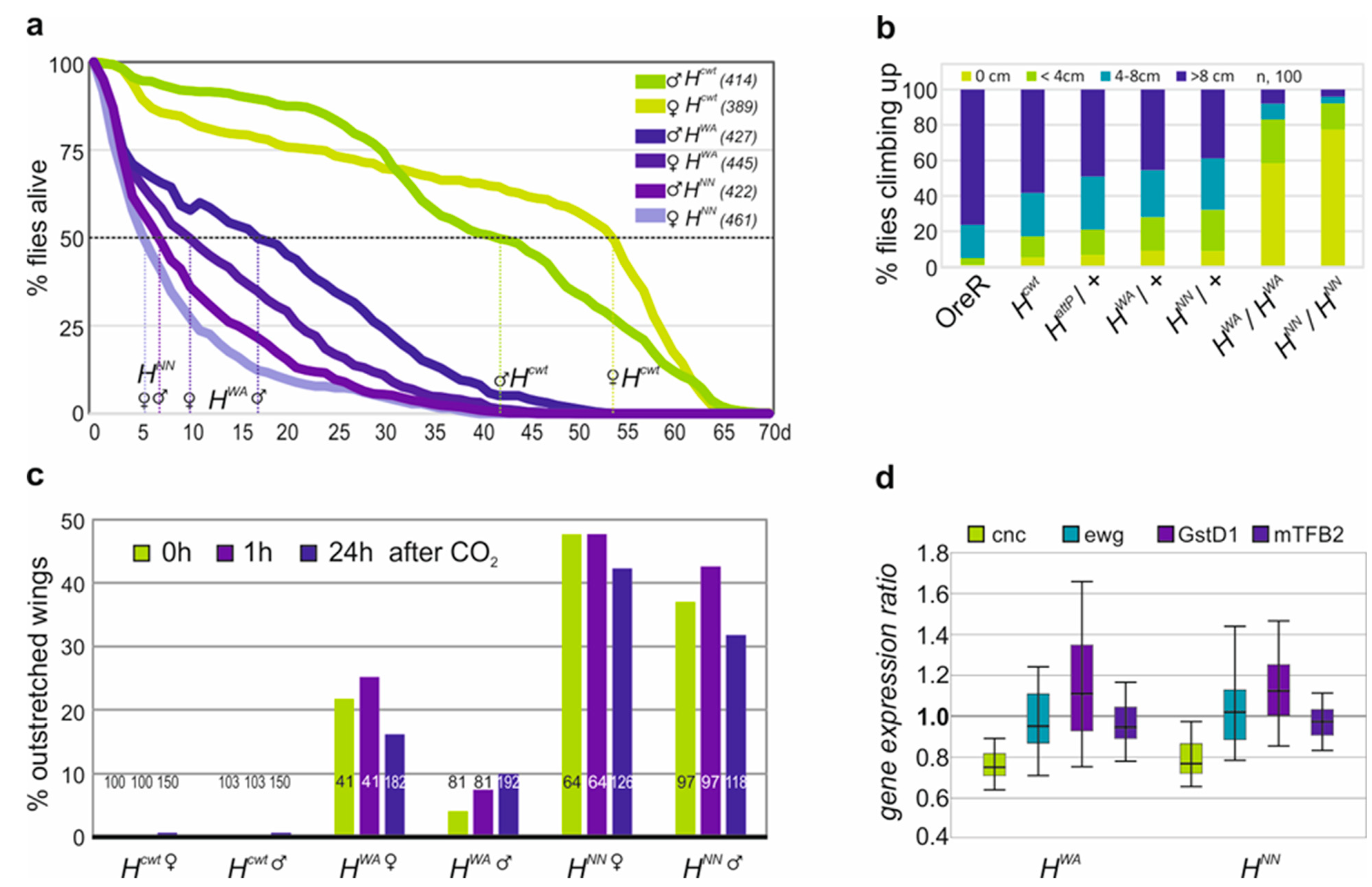
3.3. Homozygous HWA and HNN Alleles Display Strong H Loss of Function Phenotypes and Are Defective in Locomotion
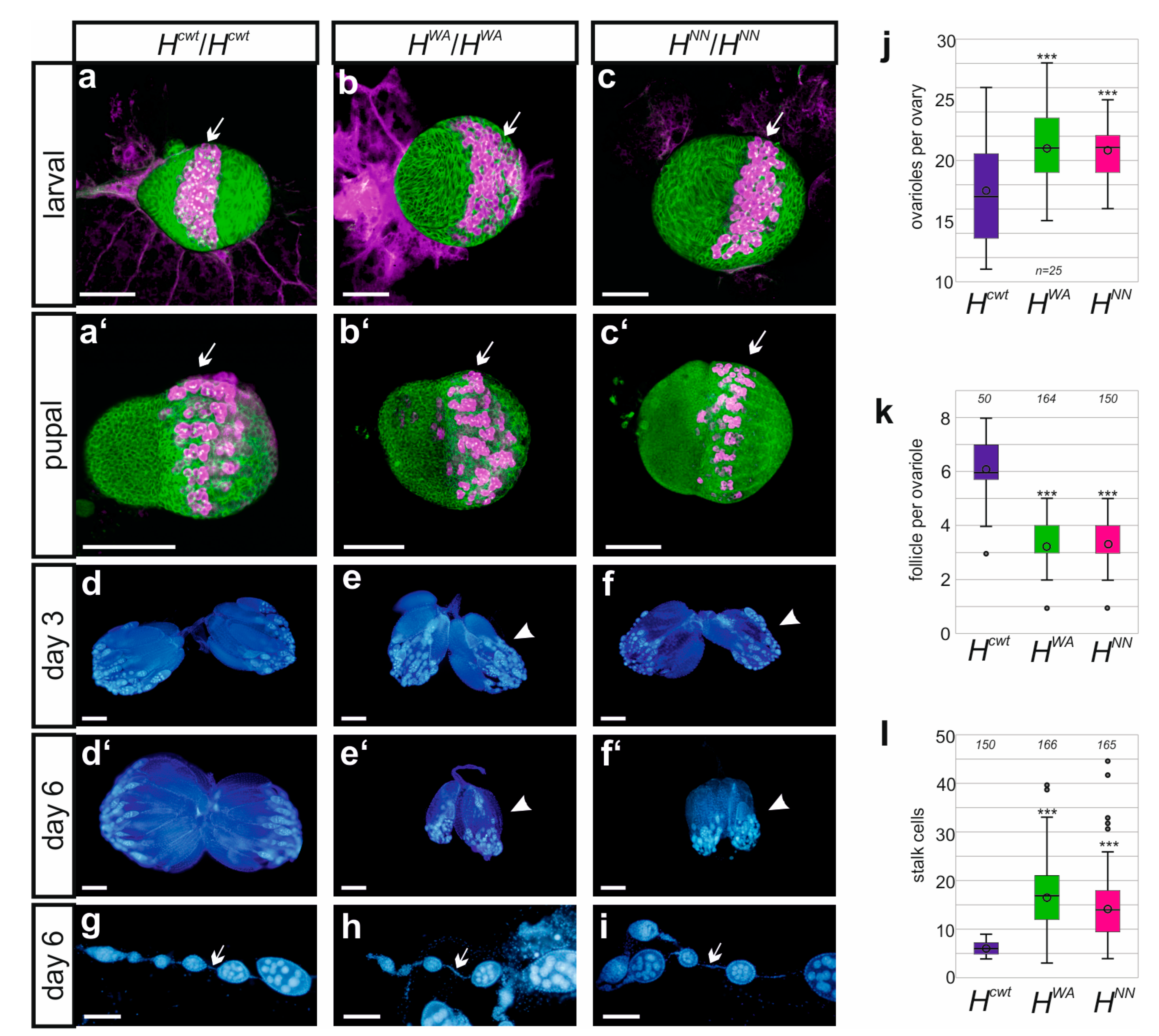
3.4. Fertility Is Impeded in the Homozygous HWA and HNN Alleles
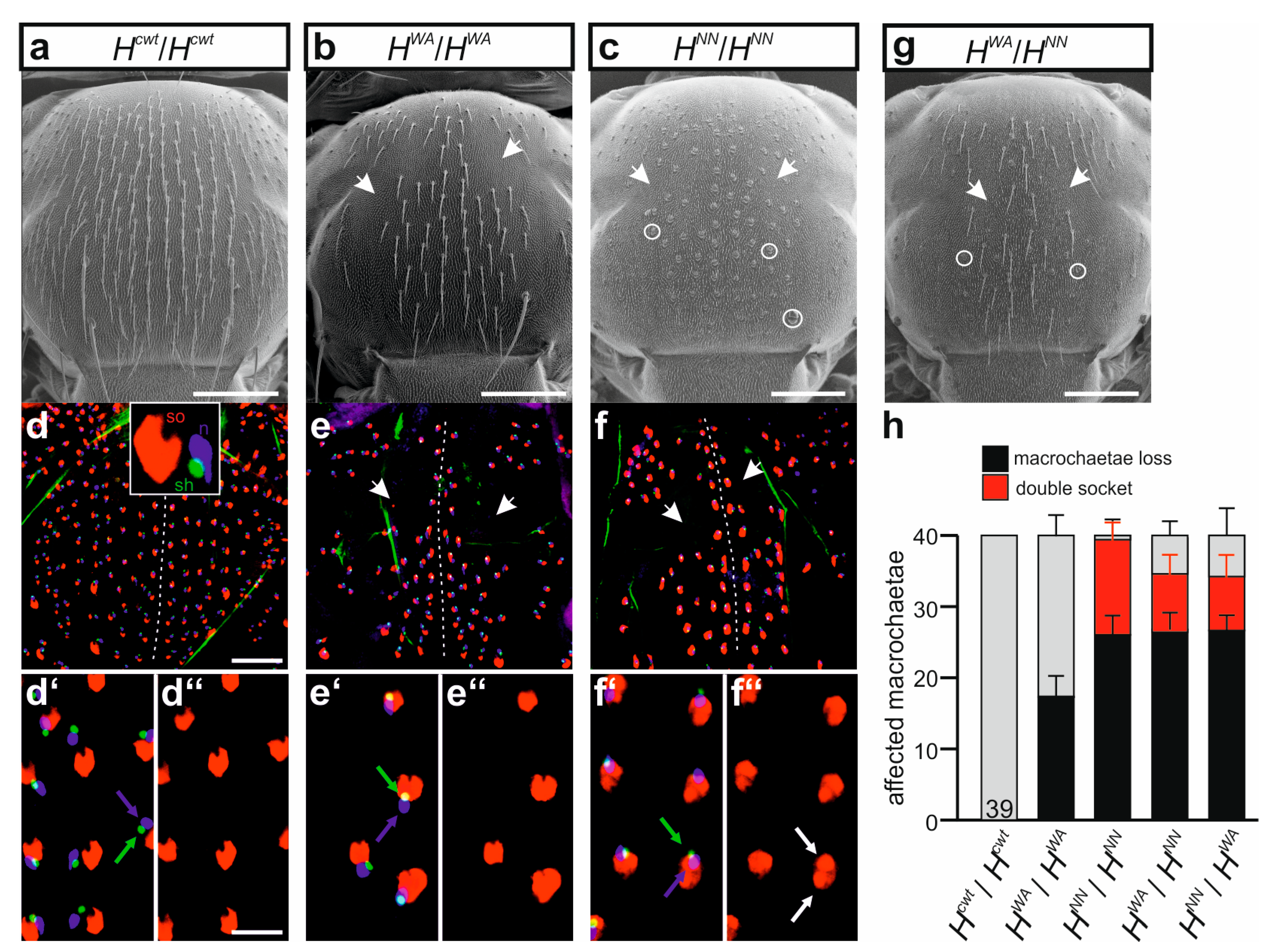
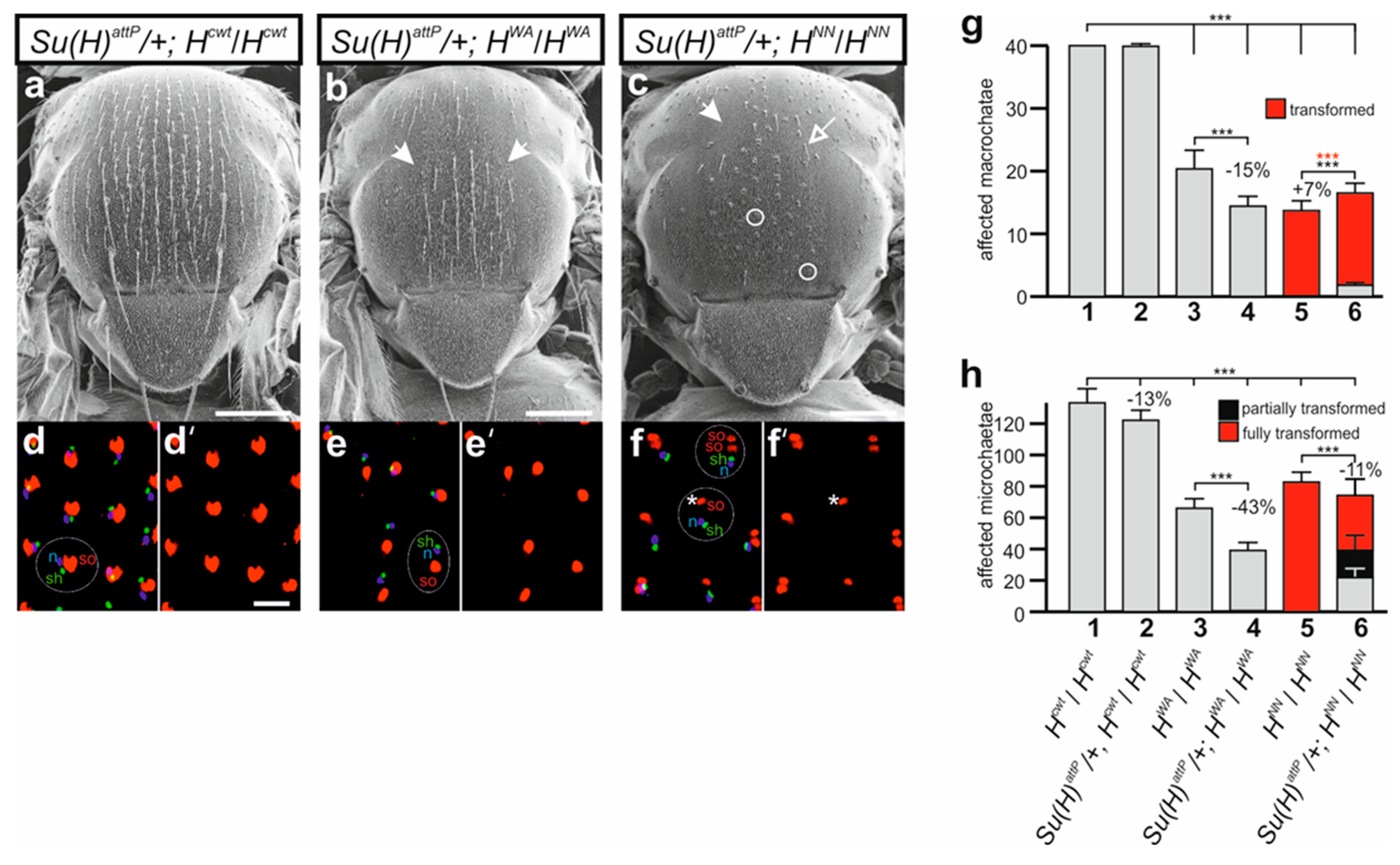
3.5. HWA Affects Lateral Inhibition during SOP Selection but Not the Asymmetric Cell Division of the SOP’s Daughters in Contrast to HNN
4. Discussion
4.1. The Three Novel H Alleles Form an Allelic Series According to Their Residual Su(H) Binding Capability
4.2. HWA and HNN Homozygotes Display Multi-Morbid Phenotypes
4.3. The Phenotypic Differences between HWA and HNN Homozygotes with Regard to Cell Type Specification Point to a Different Availability of Su(H)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kopan, R.; Ilagan, M.X. The canonical Notch signaling pathway: Unfolding the activation mechanism. Cell 2009, 137, 216–233. [Google Scholar] [CrossRef] [PubMed]
- Bray, S.J. Notch signalling: A simple pathway becomes complex. Nat. Rev. Mol. Biol. 2006, 7, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Lai, E.C. Notch signaling: Control of cell communication and cell fate. Development 2004, 131, 965–973. [Google Scholar] [CrossRef]
- Bray, S. Notch signalling in Drosophila: Three ways to use a pathway. Semin. Cell Dev. Biol. 1998, 9, 591–597. [Google Scholar] [CrossRef]
- Sjöqvist, M.; Andersson, E.R. Do as I say, Not(ch) as I do: Lateral control of cell fate. Dev. Biol. 2019, 447, 58–70. [Google Scholar] [CrossRef]
- Vasyutina, E.; Lenhard, D.C.; Birchmeier, C. Notch function in myogenesis. Cell Cycle 2007, 6, 1451–1454. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, U.; Girard, J.R.; Goins, L.M.; Spratford, C.M. Drosophila as a Genetic Model for Hematopoiesis. Genetics 2019, 211, 367–417. [Google Scholar] [CrossRef]
- Siebel, C.; Lendahl, U. Notch signaling in development, tissue homeostasis, and disease. Physiol. Rev. 2017, 97, 1235–1294. [Google Scholar] [CrossRef]
- Zamfirescu, A.M.; Yatsenko, A.S.; Shcherbata, H.R. Notch signaling sculpts the stem cell niche. Front. Cell Dev. Biol. 2022, 10, 1027222. [Google Scholar] [CrossRef]
- Lindsley, D.L.; Zimm, G.G. The Genome of Drosophila Melanogaster; Academic Press: San Diego, CA, USA, 1992. [Google Scholar]
- Simpson, P. Lateral inhibition and the development of the sensory bristles of the adult peripheral nervous system of Drosophila. Development 1990, 109, 509–519. [Google Scholar] [CrossRef]
- Gho, M.; Bellaïche, Y.; Schweisguth, F. Revisiting the Drosophila microchaete lineage: A novel intrinsically asymmetric cell division generates a glial cell. Development 1999, 126, 3573–3584. [Google Scholar] [CrossRef] [PubMed]
- Bardin, A.J.; Le Borgne, R.; Schweisguth, F. Asymmetric localization and function of cell-fate determinants: A fly’s view. Curr. Opin. Neurobiol. 2004, 14, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Hartenstein, V.; Posakony, J.W. Development of adult sensilla on the wing and notum of Drosophila melanogaster. Development 1989, 107, 389–405. [Google Scholar] [CrossRef] [PubMed]
- Posakony, J.W. Nature versus nurture: Asymmetric cell divisions in Drosophila bristle development. Cell 1994, 76, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Mohr, O.L. A genetic and cytological analysis of a section deficiency involving four units of the X-chromosome in Drosophila melanogaster. Z. Indukt. Abstamm. Vererbungsl. 1923, 32, 108–232. [Google Scholar] [CrossRef]
- de Celis, J.F.; Bray, S.; Garcia-Bellido, A. Notch signalling regulates veinlet expression and establishes boundaries between veins and interveins in the Drosophila wing. Development 1997, 124, 1919–1928. [Google Scholar] [CrossRef] [PubMed]
- Krejčí, A.; Bray, S. Notch activation stimulates transient and selective binding of Su(H)/CSL to target enhancers. Genes Dev. 2007, 21, 1322–1327. [Google Scholar] [CrossRef]
- Kovall, R.A.; Blacklow, S.C. Mechanistic insights into Notch receptor signaling from structural and biochemical studies. Curr. Top. Dev. Biol. 2010, 92, 31–71. [Google Scholar] [CrossRef] [PubMed]
- Oswald, F.; Kovall, R.A. CSL-Associated corepressor and coactivator complexes. Adv. Exp. Med. Biol. 2018, 1066, 279–295. [Google Scholar] [CrossRef]
- Maier, D. The evolution of transcriptional repressors in the Notch signaling pathway: A computational analysis. Hereditas 2019, 156, 5. [Google Scholar] [CrossRef]
- Hall, D.P.; Kovall, R.A. Structurally conserved binding motifs of transcriptional regulators to notch nuclear effector CSL. Exp. Biol. Med. 2019, 244, 1520–1529. [Google Scholar] [CrossRef] [PubMed]
- Maier, D. Hairless, the ignored antagonist of the Notch signalling pathway. Hereditas 2006, 143, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Maier, D.; Kurth, P.; Schulz, A.; Russell, A.; Yuan, Z.; Gruber, K.; Kovall, R.A.; Preiss, A. Structural and functional analysis of the repressor complex in the Notch signaling pathway of Drosophila melanogaster. Mol. Cell Biol. 2011, 22, 3242–3252. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Praxenthaler, H.; Tabaja, N.; Torella, R.; Preiss, A.; Maier, D.; Kovall, R.A. Structure and function of the Su(H)-Hairless repressor complex, the major antagonist of Notch signalling in Drosophila melanogaster. PLoS Biol. 2016, 14, e1002509. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Hayward, S.D. Nuclear localization of CBF1 is regulated by interactions with the SMRT corepressor complex. Mol. Cell Biol. 2001, 21, 6222–6232. [Google Scholar] [CrossRef] [PubMed]
- Wolf, D.; Smylla, T.K.; Reichmuth, J.; Hoffmeister, P.; Kober, L.; Zimmermann, M.; Turkiewicz, A.; Borggrefe, T.; Nagel, A.C.; Oswald, F.; et al. Nucleo-cytoplasmic shuttling of Drosophila Hairless/Su(H) heterodimer as a means of regulating Notch dependent transcription. Biochim. Biophys. Acta-Mol. Cell Res. 2019, 1866, 1520–1532. [Google Scholar] [CrossRef] [PubMed]
- Praxenthaler, H.; Nagel, A.C.; Schulz, A.; Zimmermann, M.; Meier, M.; Schmid, H.; Preiss, A.; Maier, D. Hairless-binding deficient Suppressor of Hairless alleles reveal Su(H) protein levels are dependent on complex formation with Hairless. PLoS Genet. 2017, 13, e1006774. [Google Scholar] [CrossRef] [PubMed]
- Fechner, J.; Ketelhut, M.; Maier, D.; Preiss, A.; Nagel, A.C. The binding of CSL proteins to either co-activators or co-repressors protects from proteasomal degradation induced by MAPK-dependent phosphorylation. Int. J. Mol. Sci. 2022, 23, 12336. [Google Scholar] [CrossRef] [PubMed]
- Praxenthaler, H.; Smylla, T.K.; Nagel, A.C.; Preiss, A.; Maier, D. Generation of new Hairless alleles by genomic engineering at the Hairless locus in Drosophila melanogaster. PLoS ONE 2015, 10, e0140007. [Google Scholar] [CrossRef] [PubMed]
- Smylla, T.K.; Preiss, A.; Maier, D. In vivo analysis of internal ribosome entry at the Hairless locus by genome engineering in Drosophila. Sci. Rep. 2016, 6, 34881. [Google Scholar] [CrossRef]
- Smylla, T.K.; Meier, M.; Preiss, A.; Maier, D. The Notch repressor complex in Drosophila: In vivo analysis of Hairless mutants using overexpression experiments. Dev. Genes Evol. 2019, 229, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Bang, A.G.; Hartenstein, V.; Posakony, J.W. Hairless is required for the development of adult sensory organ precursor cells in Drosophila. Development 1991, 111, 89–104. [Google Scholar] [CrossRef] [PubMed]
- Flybase. Available online: http://flybase.org/ (accessed on 15 February 2024).
- Huang, J.; Zhou, W.; Dong, W.; Watson, A.M.; Hong, Y. Directed, efficient, and versatile modifications of the Drosophila genome by genomic engineering. Proc. Natl. Acad. Sci. USA 2009, 106, 8284–8289. [Google Scholar] [CrossRef] [PubMed]
- Preiss, A.; Nagel, A.C.; Praxenthaler, H.; Maier, D. Complex genetic interactions of novel Suppressor of Hairless alleles deficient in co-repressor binding. PLoS ONE 2018, 13, e0193956. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Rubin, G.M. Analysis of genetic mosaics in developing and adult Drosophila tissues. Development 1993, 117, 1223–1237. [Google Scholar] [CrossRef]
- Gho, M.; Lecourtois, M.; Géraud, G.; Posakony, J.W.; Schweisguth, F. Subcellular localization of Suppressor of Hairless in Drosophila sense organ cells during Notch signalling. Development 1996, 122, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Nagel, A.C.; Maier, D.; Preiss, A. Su(H)-independent activity of Hairless during mechano-sensory organ formation in Drosophila. Mech. Dev. 2000, 94, 3–12. [Google Scholar] [CrossRef]
- Kober, L.; Zimmermann, M.; Kurz, M.; Bayer, M.; Nagel, A.C. Loss of putzig in the germline impedes germ cell development by inducing cell death and new niche like microenvironments. Sci. Rep. 2019, 9, 9108. [Google Scholar] [CrossRef]
- Lee, F.K.; Wong, A.K.; Lee, Y.W.; Wan, O.W.; Chan, H.Y.; Chung, K.K. The role of ubiquitin linkages on alpha-synuclein induced-toxicity in a Drosophila model of Parkinson’s disease. J. Neurochem. 2009, 110, 208–219. [Google Scholar] [CrossRef]
- Mensah, L.B.; Davison, C.; Fan, S.J.; Morris, J.F.; Goberdhan, D.C.; Wilson, C. Fine-tuning of PI3K/AKT signalling by the tumour suppressor PTEN is required for maintenance of flight muscle function and mitochondrial integrity in ageing adult Drosophila melanogaster. PLoS ONE 2015, 10, e0143818. [Google Scholar] [CrossRef]
- Villella, A.; Hall, J.C. Neurogenetics of courtship and mating in Drosophila. Adv. Genet. 2008, 62, 67–184. [Google Scholar] [CrossRef]
- Fields, S.; Song, O. A novel genetic system to detect protein–protein interactions. Nature 1989, 340, 245–246. [Google Scholar] [CrossRef] [PubMed]
- Gyuris, J.; Golemis, E.; Chertkov, H.; Brent, R. Cdi1, a human G1 and S phase protein phosphatase that associates with Cdk2. Cell 1993, 75, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Nagel, A.C.; Krejci, A.; Tenin, G.; Bravo-Patiño, A.; Bray, S.; Maier, D.; Preiss, A. Hairless-mediated repression of Notch target genes requires the combined activity of Groucho and CtBP corepressors. Mol. Cell Biol. 2005, 25, 10433–10441. [Google Scholar] [CrossRef]
- Nagel, A.C.; Maier, D.; Scharpf, J.; Ketelhut, M.; Preiss, A. Limited availability of general co-repressors uncovered in an overexpression context during wing venation in Drosophila melanogaster. Genes 2020, 11, 1141. [Google Scholar] [CrossRef] [PubMed]
- Praxenthaler, H. Molekulare und Genetische Analyse des Notch-Repressorkomplexes in D. melanogaster. In Dissertation Universität Hohenheim, Stuttgart-Hohenheim; University of Hohenheim: Stuttgart, Germany, 2015; ISBN 978-3-8439-2196-1. [Google Scholar]
- Maier, D.; Nagel, A.C.; Johannes, B.; Preiss, A. Subcellular localization of Hairless protein shows a major focus of activity within the nucleus. Mech. Dev. 1999, 89, 195–199. [Google Scholar] [CrossRef]
- Hu, Y.; Sopko, R.; Foos, M.; Kelley, C.; Flockhart, I.; Ammeux, N.; Wang, X.; Perkins, L.; Perrimon, N.; Mohr, S.E. FlyPrimerBank: An online database for Drosophila melanogaster gene expression analysis and knockdown evaluation of RNAi reagents. G3 Genes Genomes Genet. 2013, 3, 1607–1616. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for a group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Bridges, C.B.; Morgan, T.H. The third-chromosome group of mutant characters of Drosophila melanogaster. Carnegie Inst. Wash. 1923, 327, 1–251. [Google Scholar]
- Lees, A.D.; Waddington, C.H. Development of bristle mutants. Drosoph. Inf. Serv. 1942, 16, 70–70a. [Google Scholar]
- Bang, A.G.; Posakony, J.W. The Drosophila gene Hairless encodes a novel basic protein that controls alternative cell fates in adult sensory organ development. Genes Dev. 1992, 6, 1752–1769, Erratum in Genes Dev. 1992, 6, 2233. [Google Scholar] [CrossRef]
- Maier, D.; Stumm, D.; Kuhn, K.; Preiss, A. Hairless, a Drosophila gene involved in neural development, encodes a novel, serine rich protein. Mech. Dev. 1992, 38, 143–156. [Google Scholar] [CrossRef]
- Blair, S.S. Wing vein patterning in Drosophila and the analysis of intercellular signaling. Annu. Rev. Cell Dev. Biol. 2007, 23, 293–319. [Google Scholar] [CrossRef] [PubMed]
- Brou, C.; Logeat, F.; Lecourtois, M.; Vandekerckhove, J.; Kourilsky, P.; Schweisguth, F.; Israël, A. Inhibition of the DNA-binding activity of Drosophila Suppressor of Hairless and of its human homolog, KBF2/RBP-J kappa, by direct protein-protein interaction with Drosophila Hairless. Genes Dev. 1994, 8, 2491–2503. [Google Scholar] [CrossRef] [PubMed]
- Barolo, S.; Stone, T.; Bang, A.G.; Posakony, J.W. Default repression and Notch signaling: Hairless acts as an adaptor to recruit the corepressors Groucho and dCtBP to Suppressor of Hairless. Genes Dev. 2002, 16, 1964–1976. [Google Scholar] [CrossRef] [PubMed]
- Bray, S.; Furriols, M. Notch pathway: Making sense of Suppressor of Hairless. Curr. Biol. 2001, 11, R217–R221. [Google Scholar] [CrossRef]
- San Juan, B.P.; Andrade-Zapata, I.; Baonza, A. The bHLH factors Dpn and members of the E(spl) complex mediate the function of Notch signalling regulating cell proliferation during wing disc development. Biol. Open 2012, 1, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Babaoğlan, A.B.; Housden, B.E.; Furriols, M.; Bray, S.J. Deadpan contributes to the robustness of the Notch response. PLoS ONE 2013, 8, e75632. [Google Scholar] [CrossRef]
- Bartholomew, N.R.; Burdett, J.M.; VandenBrooks, J.M.; Quinlan, M.C.; Call, G.B. Impaired climbing and flight behaviour in Drosophila melanogaster following carbon dioxide anaesthesia. Sci. Rep. 2015, 5, 15298. [Google Scholar] [CrossRef]
- Rai, M.; Katti, P.; Nongthomba, U. Drosophila Erect wing (Ewg) controls mitochondrial fusion during muscle growth and maintenance by regulation of the Opa1-like gene. J. Cell Sci. 2014, 127 Pt 1, 191–203. [Google Scholar] [CrossRef]
- Matsushima, Y.; Garesse, R.; Kaguni, L.S. Drosophila mitochondrial transcription factor B2 regulates mitochondrial DNA copy number and transcription in Schneider cells. J. Biol. Chem. 2004, 279, 26900–26905. [Google Scholar] [CrossRef]
- Sawicki, R.; Singh, S.P.; Mondal, A.K.; Benes, H.; Zimniak, P. Cloning, expression and biochemical characterization of one Epsilon-class (GST-3) and ten Delta-class (GST-1) glutathione S-transferases from Drosophila melanogaster, and identification of additional nine members of the Epsilon class. Biochem. J. 2003, 370 Pt 2, 661–669. [Google Scholar] [CrossRef]
- Sykiotis, G.P.; Bohmann, D. Keap1/Nrf2 signaling regulates oxidative stress tolerance and lifespan in Drosophila. Dev. Cell 2008, 14, 76–85. [Google Scholar] [CrossRef]
- Johnstone, K.; Wells, R.E.; Strutt, D.; Zeidler, M.P. Localised JAK/STAT pathway activation is required for Drosophila wing hinge development. PLoS ONE 2013, 8, e65076. [Google Scholar] [CrossRef] [PubMed]
- Haussmann, I.U.; White, K.; Soller, M. Erect wing regulates synaptic growth in Drosophila by integration of multiple signaling pathways. Genome Biol. 2008, 9, R73. [Google Scholar] [CrossRef]
- Ruohola, H.; Bremer, K.A.; Baker, D.; Swedlow, J.R.; Jan, L.Y.; Jan, Y.N. Role of neurogenic genes in establishment of follicle cell fate and oocyte polarity during oogenesis in Drosophila. Cell 1991, 66, 433–449. [Google Scholar] [CrossRef]
- Larkin, M.K.; Holder, K.; Yost, C.; Giniger, E.; Ruohola-Baker, H. Expression of constitutively active Notch arrests follicle cells at a precursor stage during Drosophila oogenesis and disrupts the anterior-posterior axis of the oocyte. Development 1996, 122, 3639–3650. [Google Scholar] [CrossRef]
- López-Schier, H.; St Johnston, D. Delta signaling from the germ line controls the proliferation and differentiation of the somatic follicle cells during Drosophila oogenesis. Genes Dev. 2001, 15, 1393–1405. [Google Scholar] [CrossRef] [PubMed]
- Nash, D. The expression of ‘Hairless’ in Drosophila and the role of two closely linked modifiers of opposite effect. Genet. Res. 1965, 6, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M. The genetics of a small autosomal region of Drosophila melanogaster containing the structural gene for Alcohol Dehydrogenase. III. Hypomorphic and hypermorphic mutations affecting the expression of Hairless. Genetics 1982, 101, 447–459. [Google Scholar] [CrossRef]
- Schweisguth, F.; Posakony, J.W. Suppressor of Hairless, the Drosophila homolog of the mouse recombination signal-binding protein gene, controls sensory organ cell fates. Cell 1992, 69, 1199–1212. [Google Scholar] [CrossRef]
- Schweisguth, F.; Posakony, J.W. Antagonistic activities of Suppressor of Hairless and Hairless control alternative cell fates in the Drosophila adult epidermis. Development 1994, 120, 1433–1441. [Google Scholar] [CrossRef] [PubMed]
- Borggrefe, T.; Oswald, F. The Notch signaling pathway: Transcriptional regulation at Notch target genes. Cell. Mol. Life Sci. 2009, 66, 1631–1646. [Google Scholar] [CrossRef] [PubMed]
- Maier, D.; Marquart, J.; Thompson-Fontaine, A.; Beck, I.; Wurmbach, E.; Preiss, A. In vivo structure-function analysis of Drosophila Hairless. Mech. Dev. 1997, 67, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Edgar, B.A. Intestinal stem cells in the adult Drosophila midgut. Exp. Cell Res. 2011, 317, 2780–2788. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Lucchetta, E.; Rafel, N.; Ohlstein, B. Maintenance of the adult Drosophila intestine: All roads lead to homeostasis. Curr. Opin. Genet. Dev. 2016, 40, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Miguel-Aliaga, I.; Jasper, H.; Lemaitre, B. Anatomy and physiology of the digestive tract of Drosophila melanogaster. Genetics 2018, 210, 357–396. [Google Scholar] [CrossRef] [PubMed]
- Slaninova, V.; Krafcikova, M.; Perez-Gomez, R.; Steffal, P.; Trantirek, L.; Bray, S.J.; Krejci, A. Notch stimulates growth by direct regulation of genes involved in the control of glycolysis and the tricarboxylic acid cycle. Open Biol. 2016, 6, 150155. [Google Scholar] [CrossRef] [PubMed]
- Skeath, J.B.; Thor, S. Genetic control of Drosophila nerve cord development. Curr. Opin. Neurobiol. 2003, 13, 8–15. [Google Scholar] [CrossRef]
- Egger, B.; Gold, K.S.; Brand, A.H. Regulating the balance between symmetric and asymmetric stem cell division in the developing brain. Fly 2011, 5, 237–241. [Google Scholar] [CrossRef]
- Meng, J.L.; Heckscher, E.S. Development of motor circuits: From neuronal stem cells and neuronal diversity to motor circuit assembly. Curr. Top. Dev. Biol. 2021, 142, 409–442. [Google Scholar] [CrossRef]
- Poulson, D.F. Chromosomal deficiencies and the embryonic development of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1937, 23, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, R.; Jimenez, F.; Dietrich, U.; Campos-Ortega, J.A. On the phenotype and development of mutants of early neurogenesis in Drosophila melanogaster. Wilhelm Roux’s Arch. Dev. Biol. 1983, 192, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Truman, J.W.; Moats, W.; Altman, J.; Marin, E.C.; Williams, D.W. Role of Notch signaling in establishing the hemilineages of secondary neurons in Drosophila melanogaster. Development 2010, 137, 53–61. [Google Scholar] [CrossRef]
- Bivik, C.; MacDonald, R.B.; Gunnar, E.; Mazouni, K.; Schweisguth, F.; Thor, S. Control of neural daughter cell proliferation by multi-level Notch/Su(H)/E(spl)-HLH signaling. PLoS Genet. 2016, 12, e1005984. [Google Scholar] [CrossRef] [PubMed]
- Konstantinides, N.; Holguera, I.; Rossi, A.M.; Escobar, A.; Dudragne, L.; Chen, Y.C.; Tran, T.N.; Martínez Jaimes, A.M.; Özel, M.N.; Simon, F.; et al. A complete temporal transcription factor series in the fly visual system. Nature 2022, 604, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Presente, A.; Boyles, R.S.; Serway, C.N.; de Belle, J.S.; Andres, A.J. Notch is required for long-term memory in Drosophila. Proc. Natl. Acad. Sci. USA 2004, 101, 1764–1768. [Google Scholar] [CrossRef]
- Salazar, J.L.; Yang, S.A.; Yamamoto, S. Post-developmental roles of Notch signaling in the nervous system. Biomolecules 2020, 10, 985. [Google Scholar] [CrossRef]
- Nellesen, D.T.; Lai, E.C.; Posakony, J.W. Discrete enhancer elements mediate selective responsiveness of Enhancer of split complex genes to common transcriptional activators. Dev. Biol. 1999, 213, 33–53. [Google Scholar] [CrossRef]
- Bang, A.G.; Bailey, A.M.; Posakony, J.W. Hairless promotes stable commitment to the sensory organ precursor cell fate by negatively regulating the activity of the Notch signaling pathway. Dev. Biol. 1995, 172, 479–494. [Google Scholar] [CrossRef]
- Castro, B.; Barolo, S.; Bailey, A.M.; Posakony, J.W. Lateral inhibition in proneural clusters: Cis-regulatory logic and default repression by Suppressor of Hairless. Development 2005, 132, 3333–3344. [Google Scholar] [CrossRef]
- Troost, T.; Schneider, M.; Klein, T. A re-examination of the selection of the sensory organ precursor of the bristle sensilla of Drosophila melanogaster. PLoS Genet. 2015, 11, e1004911. [Google Scholar] [CrossRef] [PubMed]
- Usui, K.; Kimura, K.I. Sequential emergence of the evenly spaced microchaetes on the notum of Drosophila. Rouxs Arch. Dev. Biol. 1993, 203, 151–158. [Google Scholar] [CrossRef]
- Corson, F.; Couturier, L.; Rouault, H.; Mazouni, K.; Schweisguth, F. Self-organized Notch dynamics generate stereotyped sensory organ patterns in Drosophila. Science 2017, 356, eaai7407. [Google Scholar] [CrossRef] [PubMed]
- Couturier, L.; Mazouni, K.; Corson, F.; Schweisguth, F. Regulation of Notch output dynamics via specific E(spl)-HLH factors during bristle patterning in Drosophila. Nat. Commun. 2019, 10, 3486. [Google Scholar] [CrossRef]
- Ayeni, J.O.; Audibert, A.; Fichelson, P.; Srayko, M.; Gho, M.; Campbell, S.D. G2 phase arrest prevents bristle progenitor self-renewal and synchronizes cell division with cell fate differentiation. Development 2016, 143, 1160–1169. [Google Scholar] [CrossRef]
- Ramat, A.; Audibert, A.; Louvet-Vallée, S.; Simon, F.; Fichelson, P.; Gho, M. Escargot and Scratch regulate neural commitment by antagonizing Notch activity in Drosophila sensory organs. Development 2016, 143, 3024–3034. [Google Scholar] [CrossRef] [PubMed]
- Schweisguth, F. Asymmetric cell division in the Drosophila bristle lineage: From the polarization of sensory organ precursor cells to Notch-mediated binary fate decision. WIREs Dev. Biol. 2015, 4, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Younger-Shepherd, S.; Jan, L.Y.; Jan, Y.N. Delta and Serrate are redundant Notch ligands required for asymmetric cell divisions within the Drosophila sensory organ lineage. Genes Dev. 1998, 12, 1086–1091. [Google Scholar] [CrossRef]
- Kandachar, V.; Roegiers, F. Endocytosis and control of Notch signaling. Curr. Opin. Cell Biol. 2012, 24, 534–554. [Google Scholar] [CrossRef]
- Miller, S.W.; Avidor-Reiss, T.; Polyanovsky, A.; Posakony, J.W. Complex interplay of three transcription factors in controlling the tormogen differentiation program of Drosophila mechanoreceptors. Dev. Biol. 2009, 329, 386–399. [Google Scholar] [CrossRef]
- Kavaler, J.; Fu, W.; Duan, H.; Noll, M.; Posakony, J.W. An essential role for the Drosophila Pax2 homolog in the differentiation of adult sensory organs. Development 1999, 126, 2261–2272. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.A.; Harmon, K.J.; Smiley, S.G.; Still, F.M.; Kavaler, J. Discrete regulatory regions control early and late expression of D-Pax2 during external sensory organ development. Dev. Dyn. 2011, 240, 1769–1778. [Google Scholar] [CrossRef] [PubMed]
- Barolo, S.; Walker, R.G.; Polyanovsky, A.D.; Freschi, G.; Keil, R.; Posakony, J.W. A Notch-independent activity of Suppressor of Hairless is required for normal mechanoreceptor physiology. Cell 2000, 103, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Ohnuki, H.; Inoue, H.; Takemori, N.; Nakayama, H.; Sakaue, T.; Fukuda, S.; Miwa, D.; Nishiwaki, E.; Hatano, M.; Tokuhisa, T. BAZF, a novel component of cullin3-based E3 ligase complex, mediates VEGFR and Notch cross-signaling in angiogenesis. Blood 2012, 119, 2688–2698. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Kim, M.Y.; Ann, E.J.; Mo, J.S.; Yoon, J.H.; Park, H.S. Presenilin-2 regulates the degradation of RBP-Jk protein through p38 mitogen-activated protein kinase. J. Cell Sci. 2012, 125, 1296–1308. [Google Scholar] [CrossRef][Green Version]
- Deshmukh, R.S.; Sharma, S.; Das, S. Cyclin F-Dependent Degradation of RBPJ Inhibits IDH1R132H-Mediated Tumorigenesis. Cancer Res. 2018, 78, 6386–6398. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mönch, T.C.; Smylla, T.K.; Brändle, F.; Preiss, A.; Nagel, A.C. Novel Genome-Engineered H Alleles Differentially Affect Lateral Inhibition and Cell Dichotomy Processes during Bristle Organ Development. Genes 2024, 15, 552. https://doi.org/10.3390/genes15050552
Mönch TC, Smylla TK, Brändle F, Preiss A, Nagel AC. Novel Genome-Engineered H Alleles Differentially Affect Lateral Inhibition and Cell Dichotomy Processes during Bristle Organ Development. Genes. 2024; 15(5):552. https://doi.org/10.3390/genes15050552
Chicago/Turabian StyleMönch, Tanja C., Thomas K. Smylla, Franziska Brändle, Anette Preiss, and Anja C. Nagel. 2024. "Novel Genome-Engineered H Alleles Differentially Affect Lateral Inhibition and Cell Dichotomy Processes during Bristle Organ Development" Genes 15, no. 5: 552. https://doi.org/10.3390/genes15050552
APA StyleMönch, T. C., Smylla, T. K., Brändle, F., Preiss, A., & Nagel, A. C. (2024). Novel Genome-Engineered H Alleles Differentially Affect Lateral Inhibition and Cell Dichotomy Processes during Bristle Organ Development. Genes, 15(5), 552. https://doi.org/10.3390/genes15050552