
Assessment of the Effect of Leonurine Hydrochloride in a Mouse Model of PCOS by Gene Expression Profiling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. PCOS Model Construction and Leo Treatment
2.3. Detection of the Oestrus Cycle
2.4. Determination of Serum Hormones
2.5. Testing of Ovarian Tissue
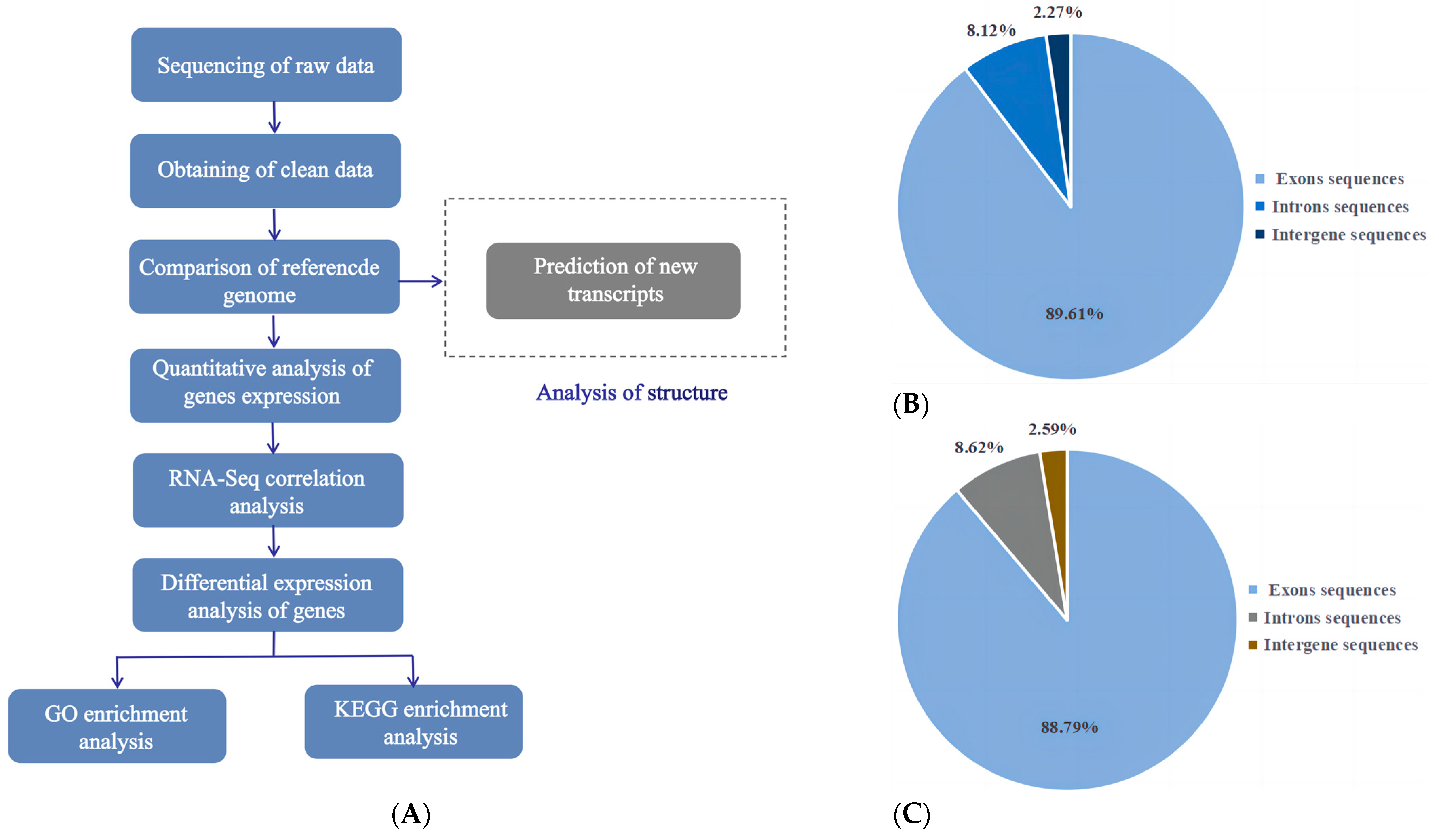
2.6. Library Construction and Sequencing
2.7. Sequence Processing and Analysis
2.8. Identification of Differentially Expressed Genes
2.9. Functional Enrichment Analysis of the DEGs
2.10. Differential Gene Expression Analysis and RT-qPCR Validation
2.11. Statistical Analysis
3. Results
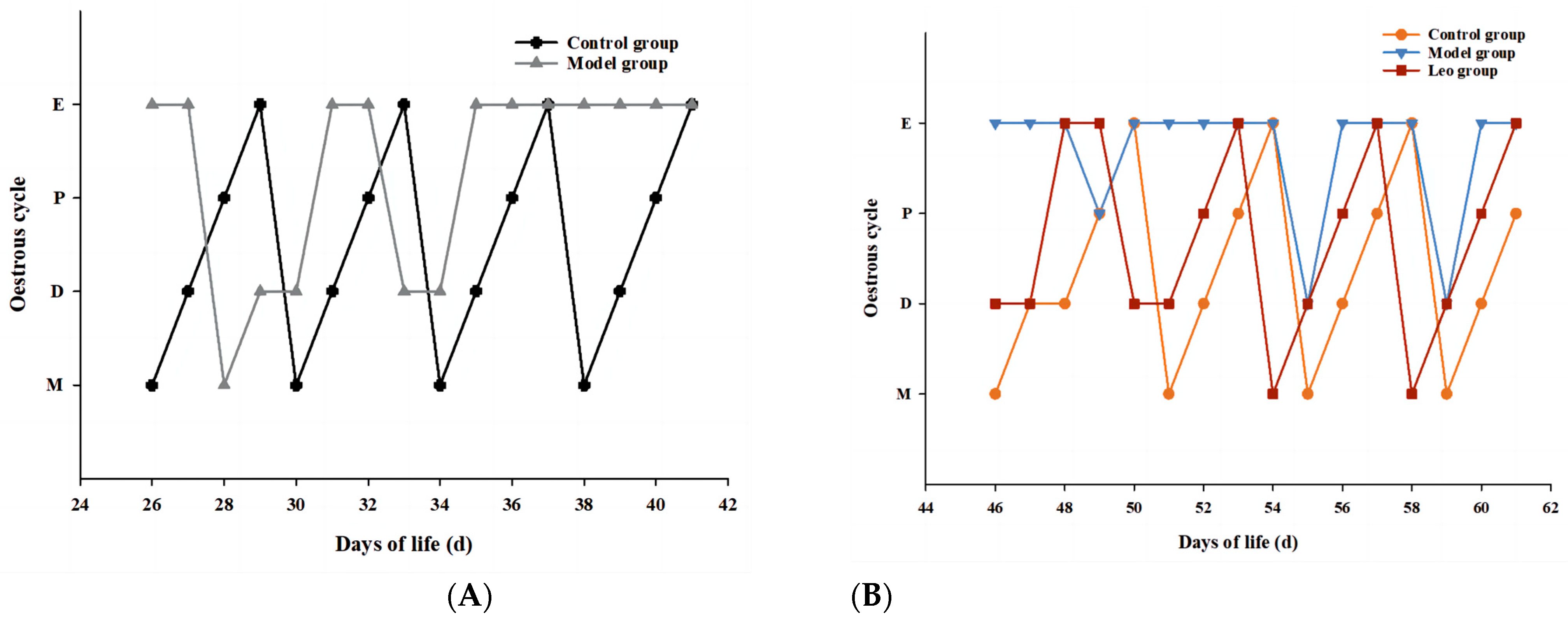
3.1. Effects of Leo on the Oestrous Cycle in Mice
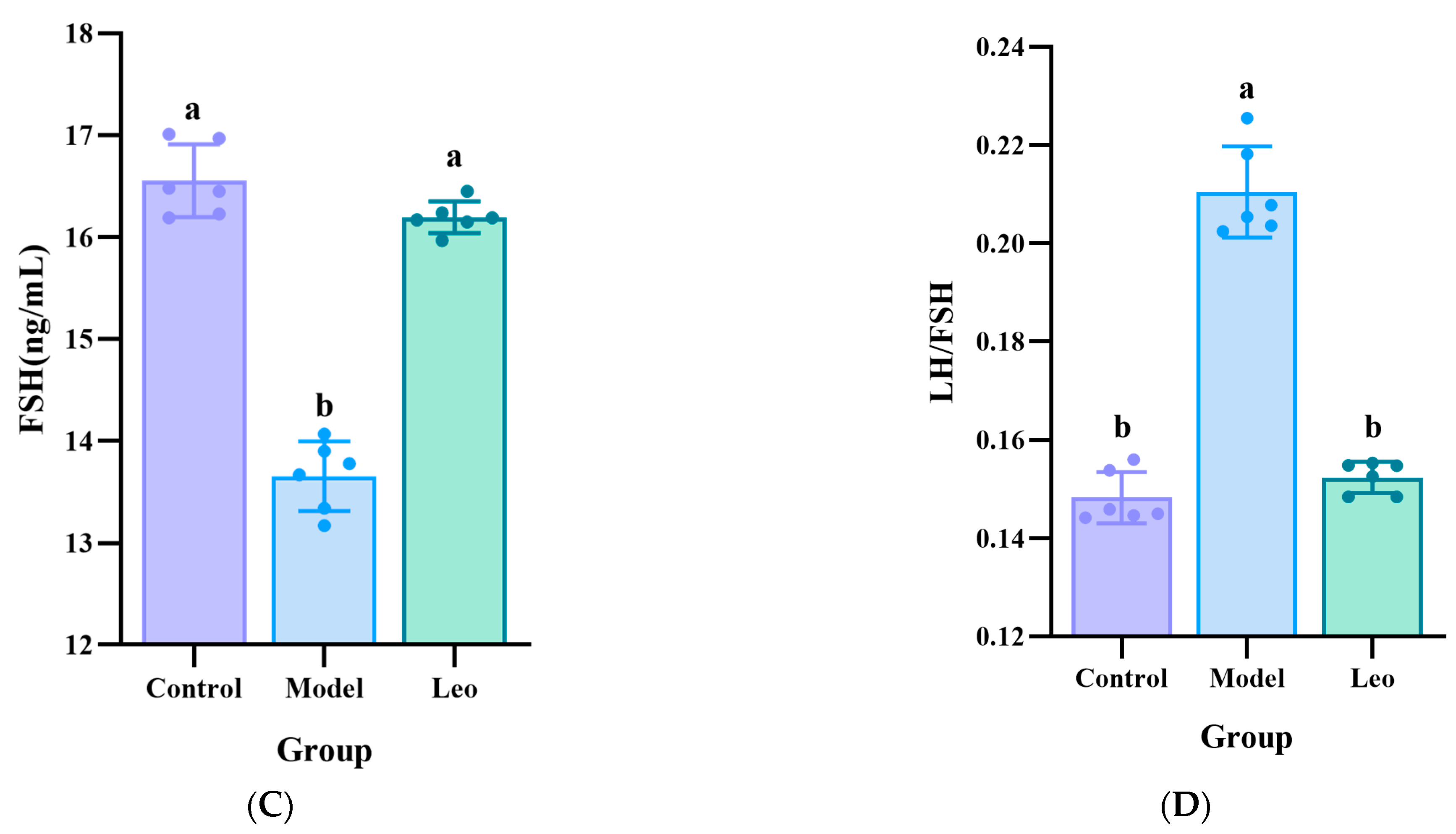
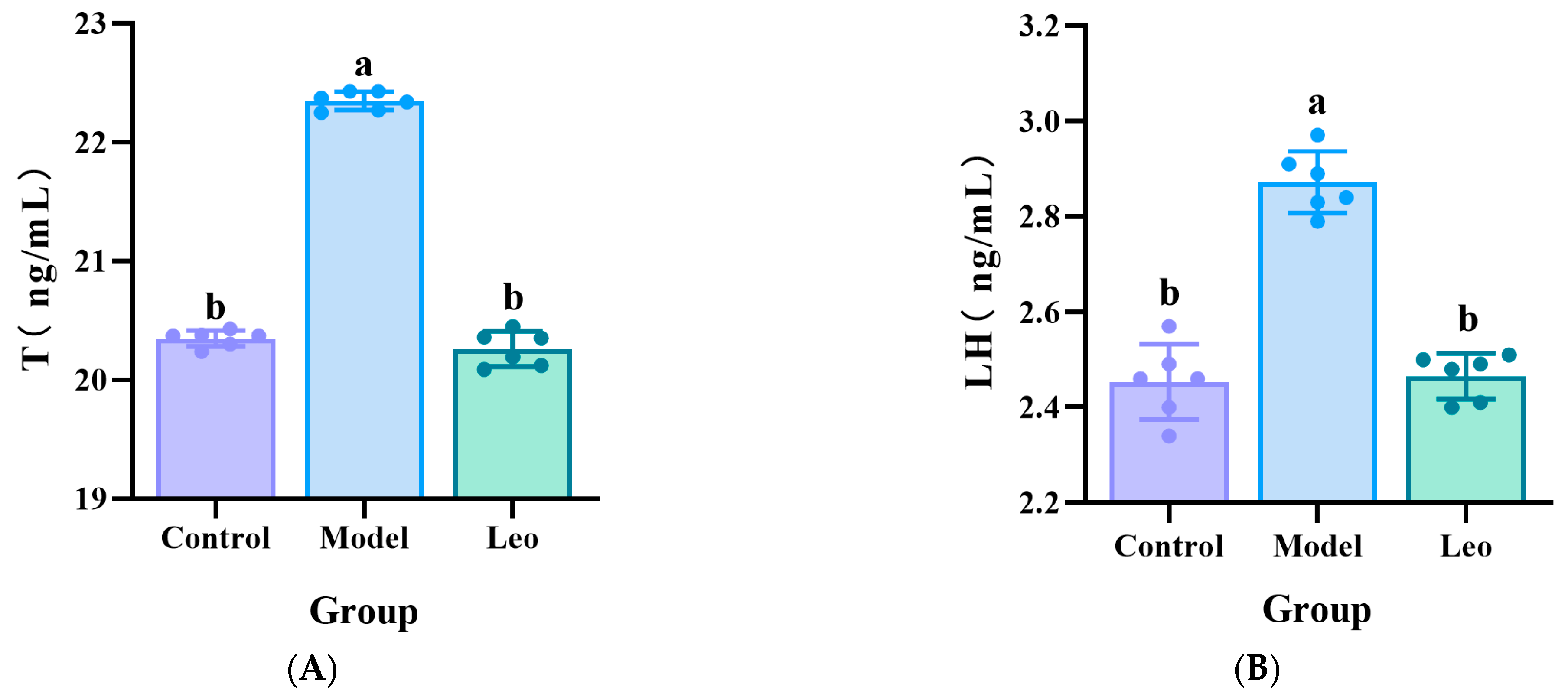
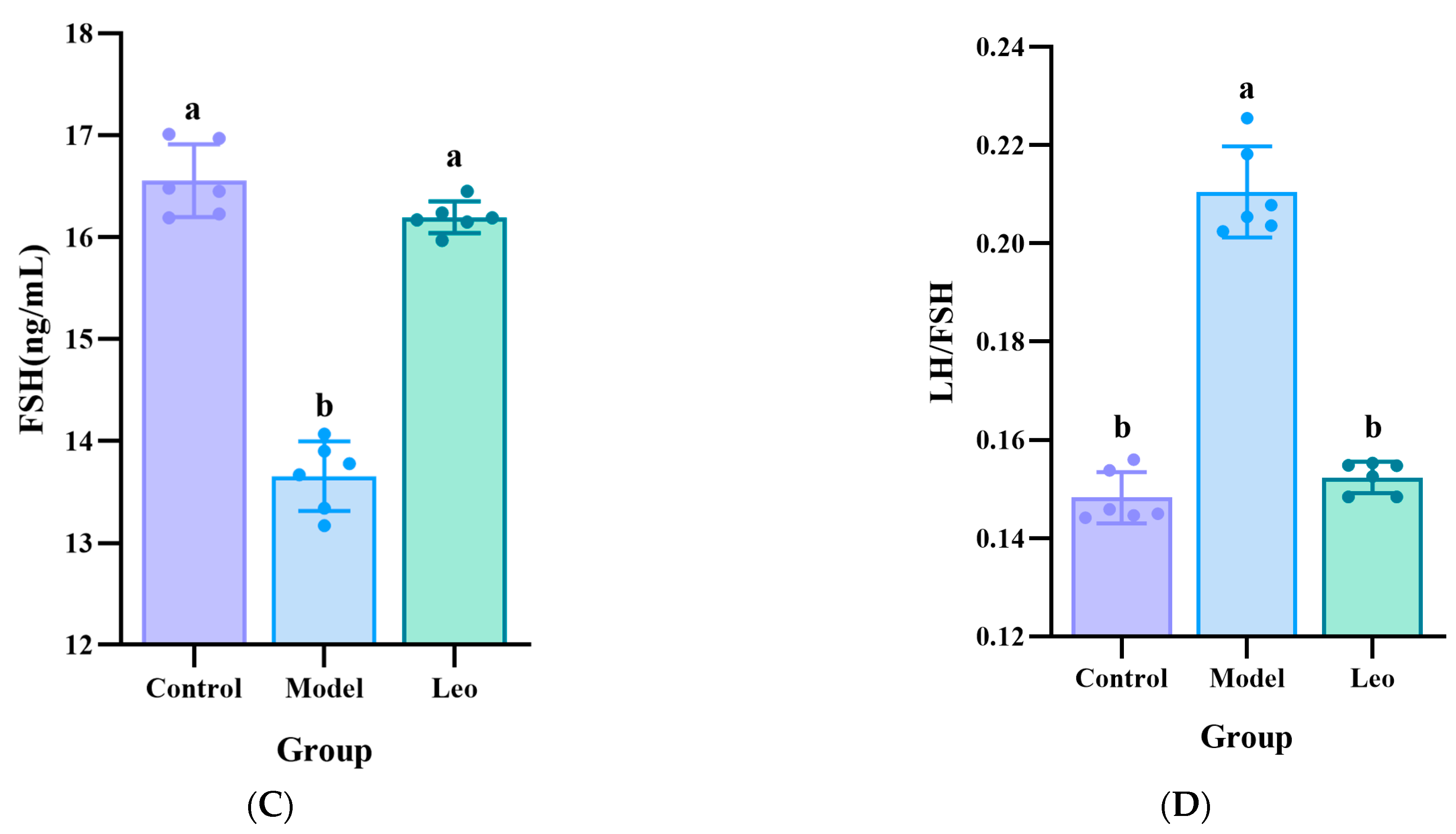
3.2. Effects of Leo on the Serum Hormone Levels in the PCOS Mouse Model
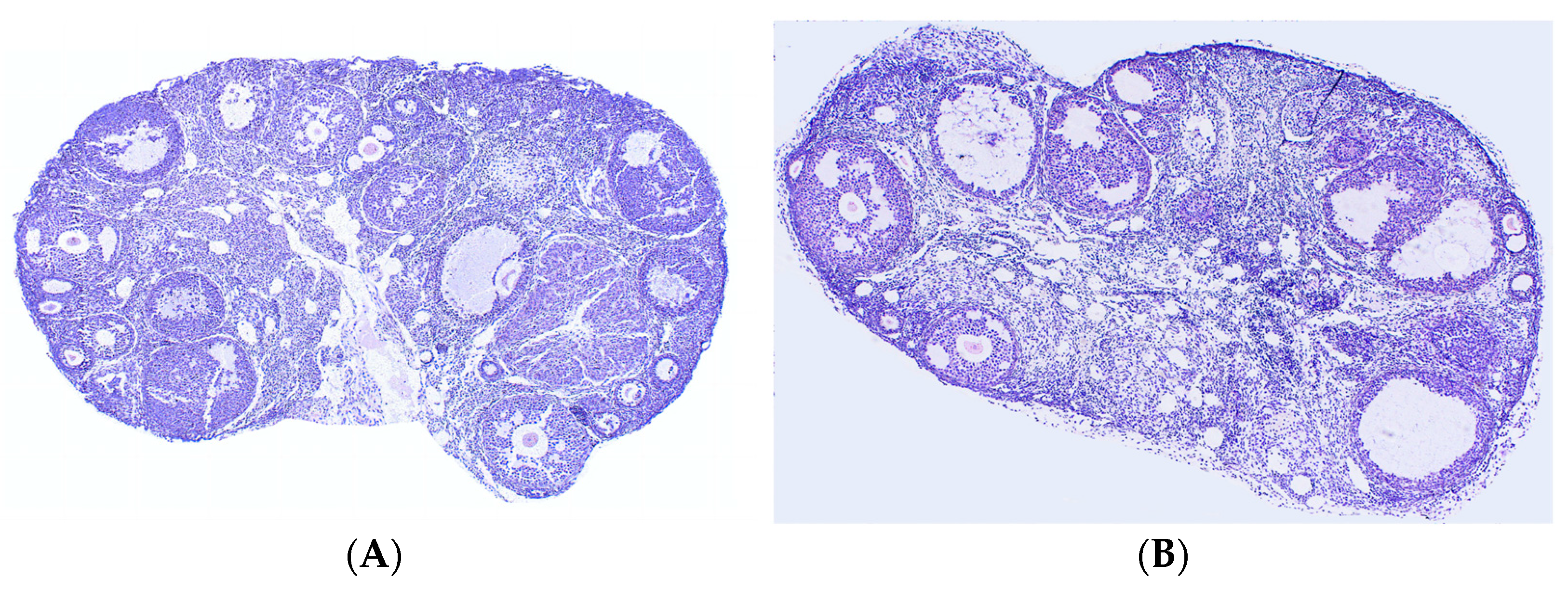
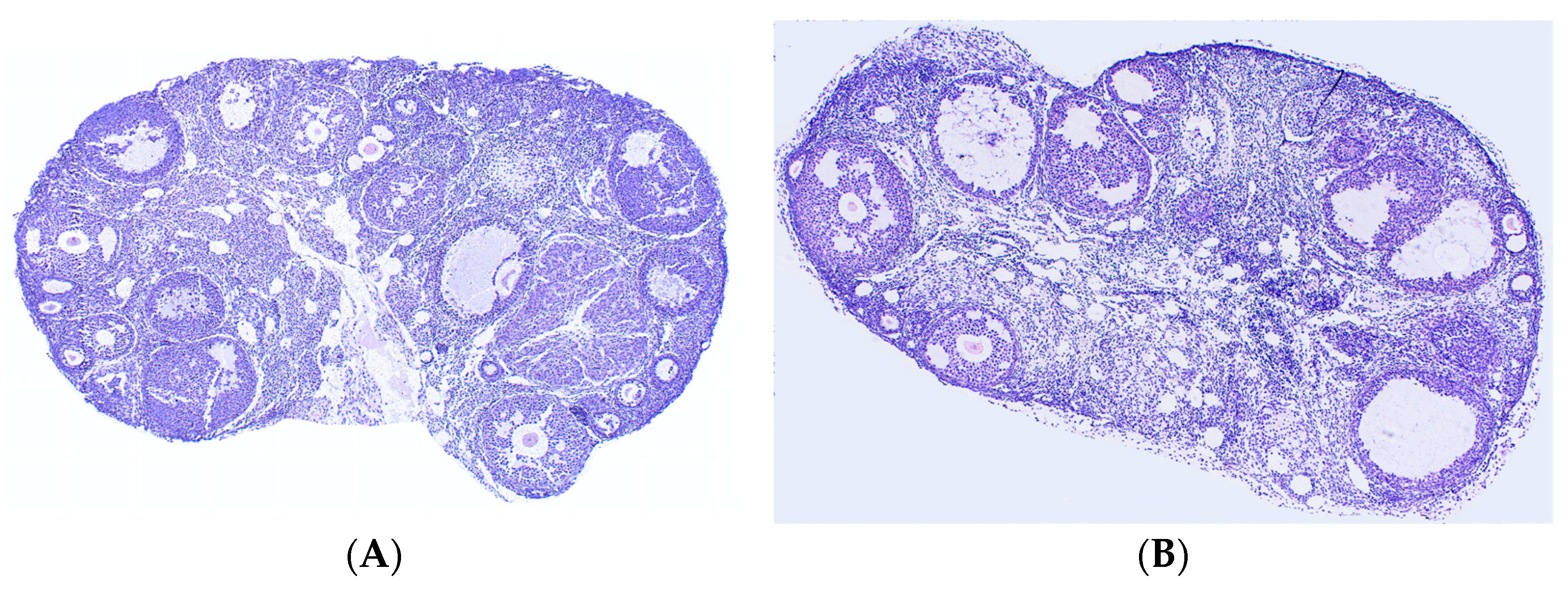
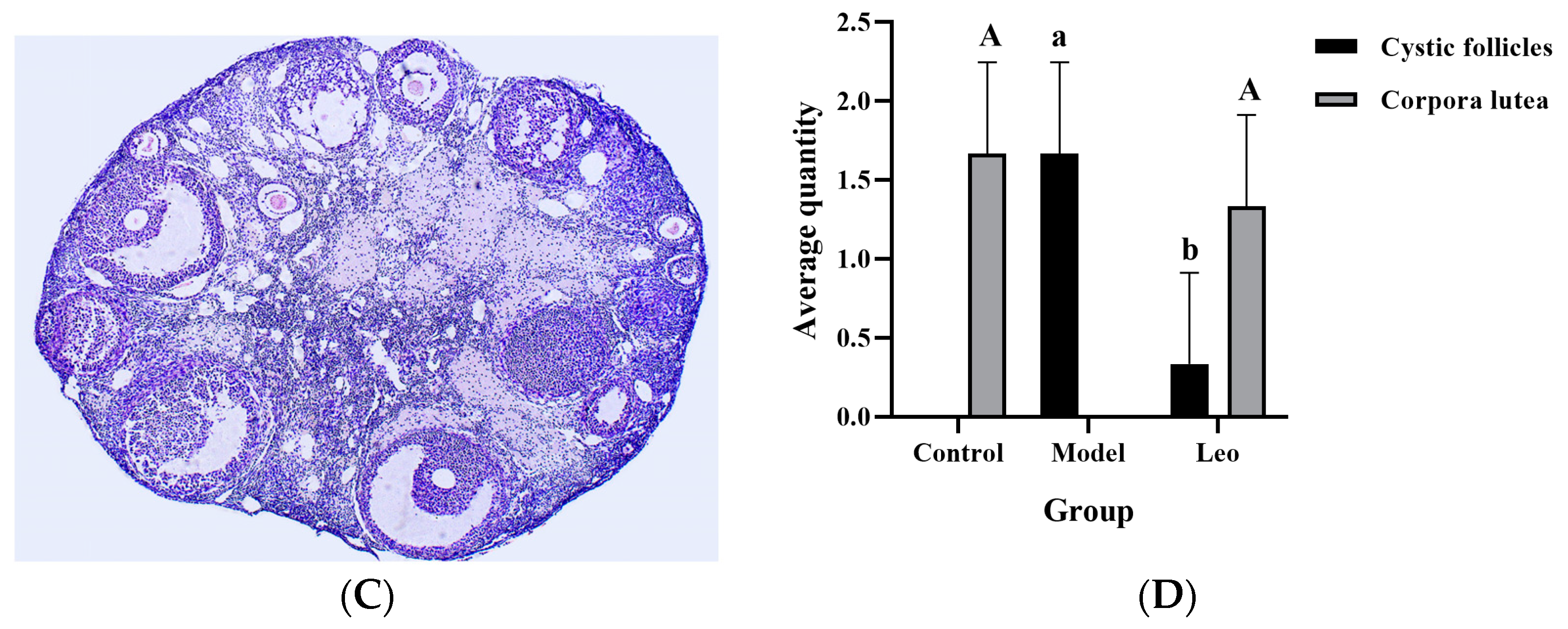
3.3. Effects of Leo on the Ovaries of PCOS Model Mice
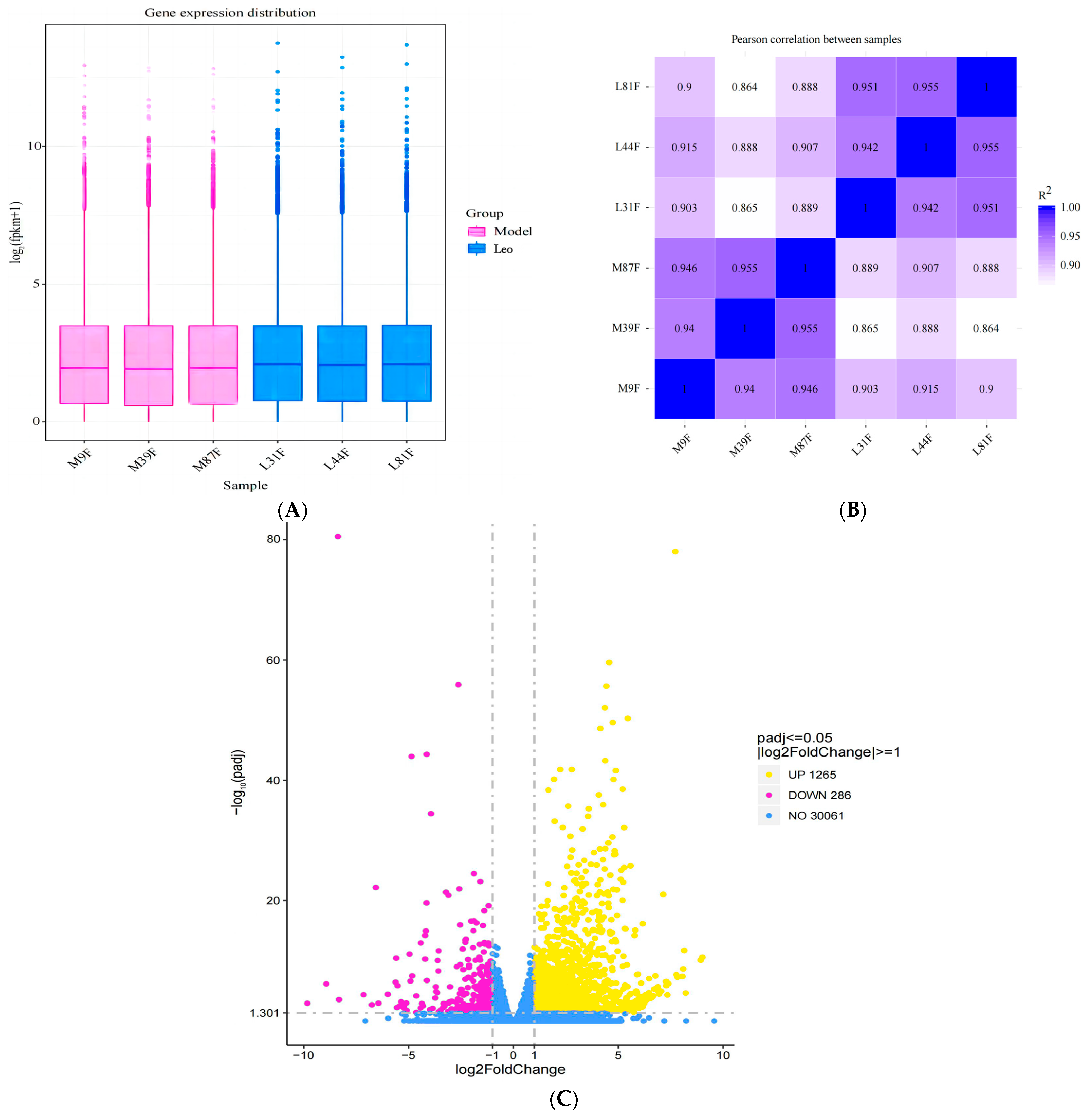
3.4. Basic RNA-Seq Data Analysis
3.5. Identification of DEGs
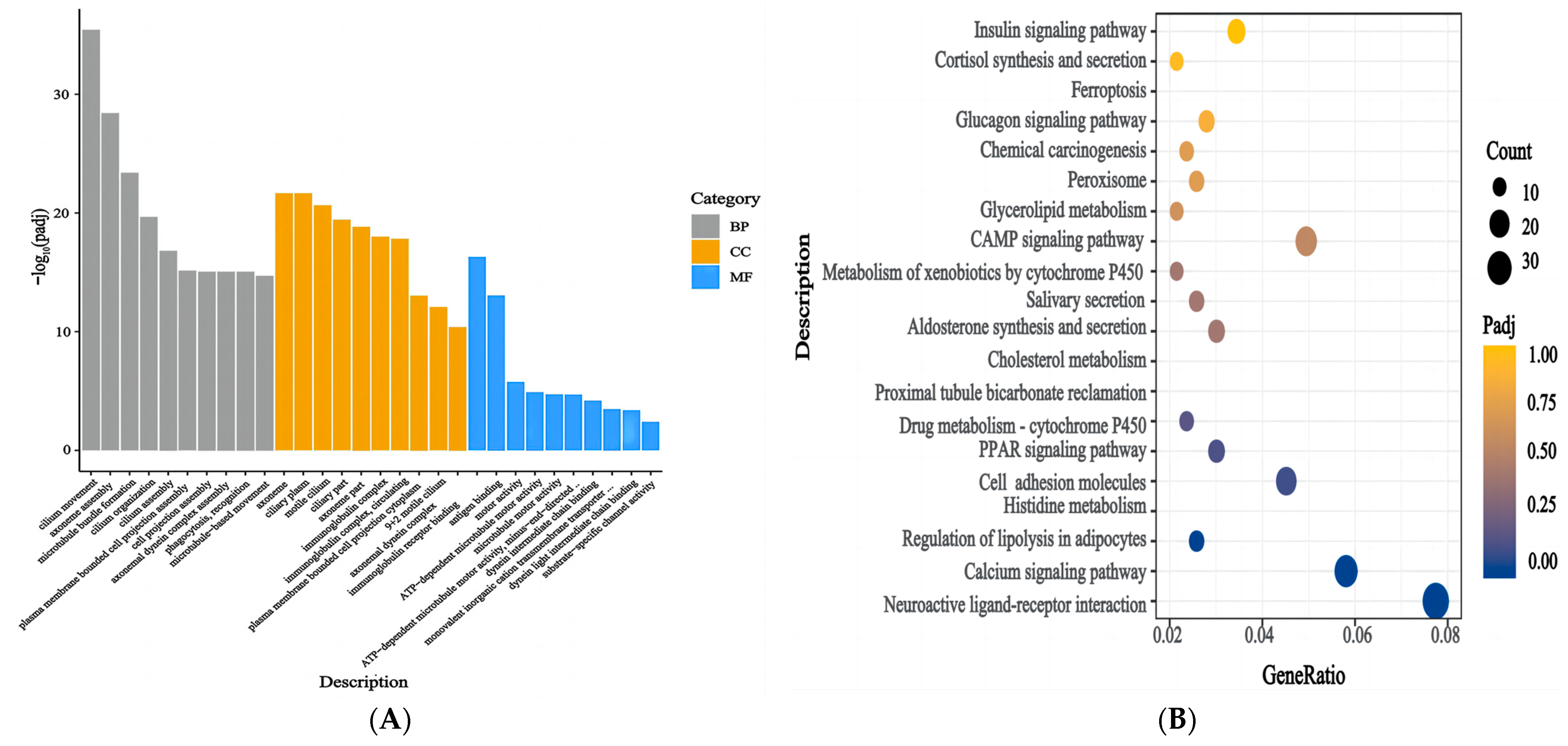
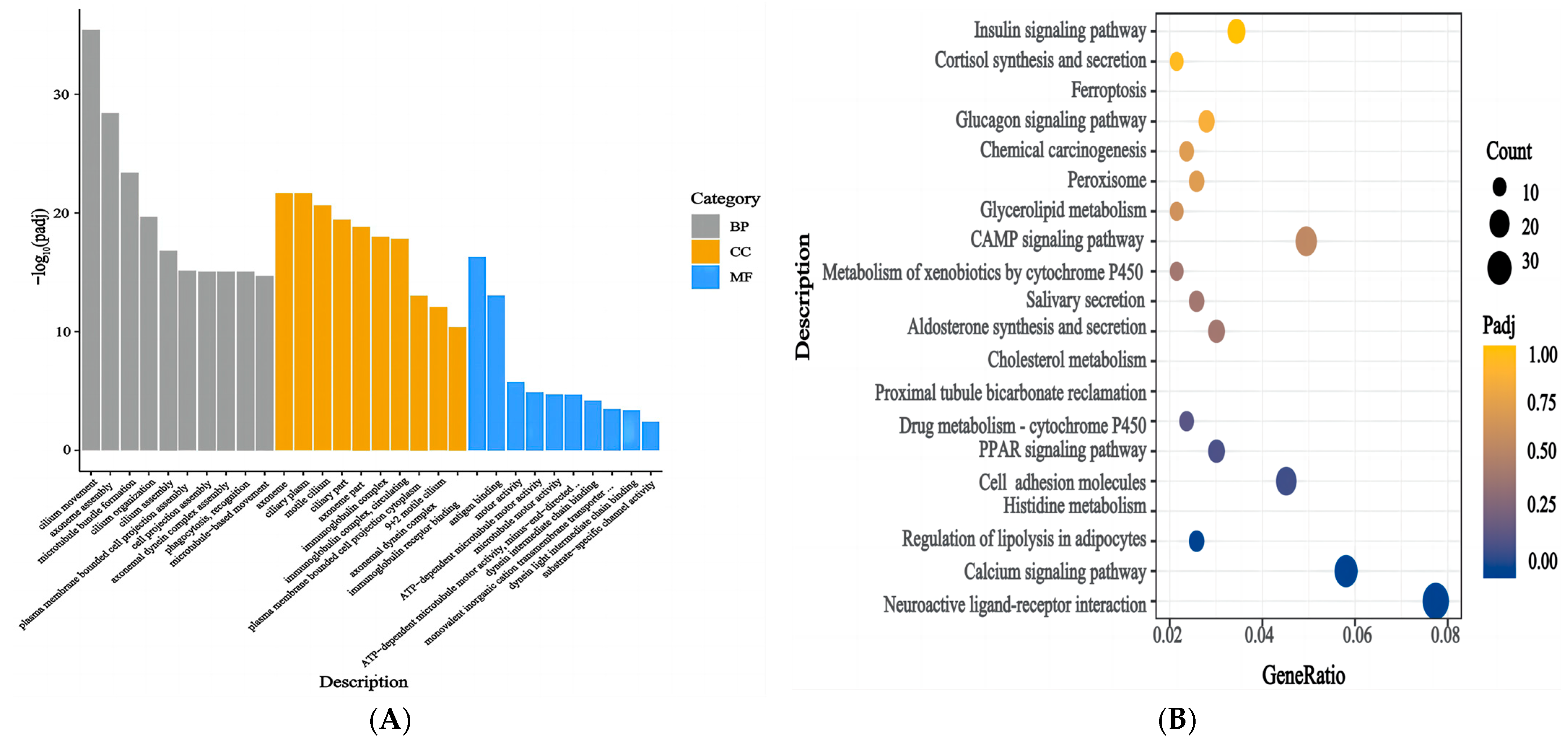
3.6. Functional Enrichment Analysis of the DEGs
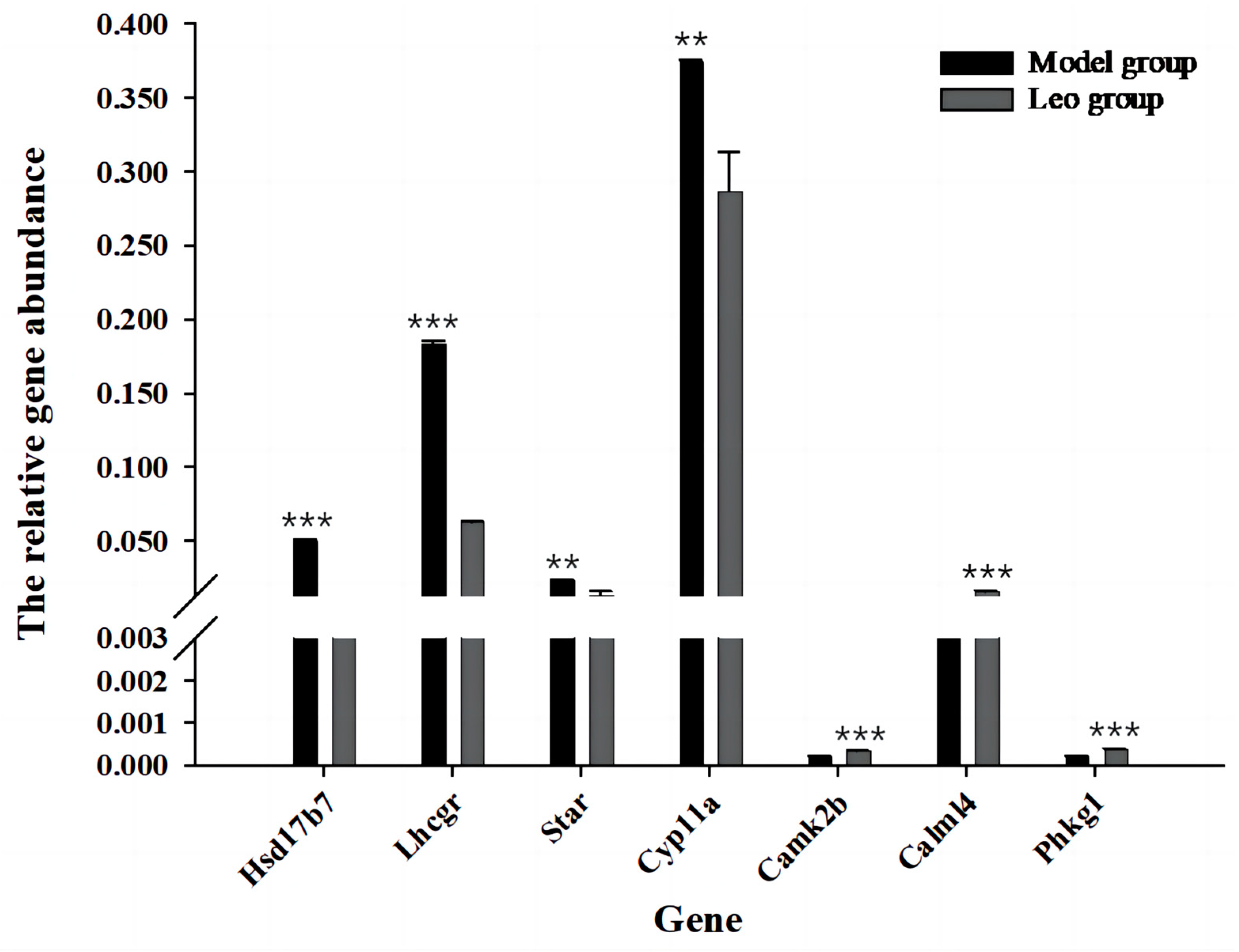
3.7. Differential Gene Expression Analysis and RT-qPCR Validation
3.8. Analysis of DEGs Most Relevant to Ovary and Validation by RT-qPCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cirillo, F.; Catellani, C.; Lazzeroni, P.; Sartori, C.; Nicoli, A.; Amarri, S.; La Sala, G.B.; Street, M.E. MiRNAs Regulating Insulin Sensitivity Are Dysregulated in Polycystic Ovary Syndrome (PCOS) Ovaries and Are Associated with Markers of Inflammation and Insulin Sensitivity. Front. Endocrinol. 2019, 10, 879. [Google Scholar] [CrossRef] [PubMed]
- Fauser, B.C.; Tarlatzis, B.C.; Rebar, R.W.; Legro, R.S.; Balen, A.H.; Lobo, R.; Carmina, E.; Chang, J.; Yildiz, B.O.; Laven, J.S. Consensus on women’s health aspects of polycystic ovary syndrome (PCOS): The Amsterdam ESHRE/ASRM-sponsored 3rd PCOS consensus workshop group. Fertil. Steril. 2012, 97, 28–38.e25. [Google Scholar] [CrossRef] [PubMed]
- Abbott, D.H.; Dumesic, D.A.; Franks, S. Developmental origin of polycystic ovary syndrome—A hypothesis. J. Endocrinol. 2002, 174, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, M.O.; Dumesic, D.A.; Chazenbalk, G.; Azziz, R. Polycystic ovary syndrome: Etiology, pathogenesis and diagnosis. Nat. Rev. Endocrinol. 2011, 7, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Dumesic, D.A.; Oberfield, S.E.; Stener-Victorin, E.; Marshall, J.C.; Laven, J.S.; Legro, R.S. Scientific statement on the diagnostic criteria, epidemiology, pathophysiology, and molecular genetics of polycystic ovary syndrome. Endocr. Rev. 2015, 36, 487–525. [Google Scholar] [CrossRef] [PubMed]
- Diamanti-Kandarakis, E.; Dunaif, A. Insulin resistance and the polycystic ovary syndrome revisited: An update on mechanisms and implications. Endocr. Rev. 2012, 33, 981–1030. [Google Scholar] [CrossRef] [PubMed]
- Tsilchorozidou, T.; Overton, C.; Conway, G.S. The pathophysiology of polycystic ovary syndrome. Clin. Endocrinol. 2004, 60, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Gersak, K.; Ferk, P. Genetics of polycystic ovary syndrome. Gynaecol. J. Perinatol. 2007, 16, 53–57. [Google Scholar]
- Escobar-Morreale, H.F.; Luque-Ramírez, M.; San Millán, J.L. The molecular-genetic basis of functional hyperandrogenism and the polycystic ovary syndrome. Endocr. Rev. 2005, 26, 251–282. [Google Scholar] [CrossRef]
- Chen, C.; Zhu, Z.; Hu, N.; Liang, X.; Huang, W. Leonurine Hydrochloride Suppresses Inflammatory Responses and Ameliorates Cartilage Degradation in Osteoarthritis via NF-κB Signaling Pathway. Inflammation 2020, 43, 146–154. [Google Scholar] [CrossRef]
- Yan, K.; Hu, J.; Hou, T.; Ci, X.; Peng, L. Leonurine inhibits the TXNIP/NLRP3 and NF-κB pathways via Nrf2 activation to alleviate carrageenan-induced pleurisy in mice. Phytother. Res. 2022, 36, 2161–2172. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Li, D.; Ouyang, J.; Tian, X.; Zhao, Y.; Peng, X.; Li, S.; Yu, G.; Yang, J. Leonurine alleviates LPS-induced myocarditis through suppressing the NF-кB signaling pathway. Toxicology 2019, 422, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.; Gong, L.; Zhou, M.; Xue, X.; Li, Y.; Peng, C. Leonurine Ameliorates Oxidative Stress and Insufficient Angiogenesis by Regulating the PI3K/Akt-eNOS Signaling Pathway in H2O2-Induced HUVECs. Oxid. Med. Cell. Longev. 2021, 2021, 9919466. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Wu, H.; Zhou, Z.; Tao, Z.; Gao, W.; Jia, T. The Effect of Leonurine on Multiterritory Perforator Flap Survival in Rats. J. Surg. Res. 2020, 245, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Xu, D.Q.; Chen, Y.Y.; Yue, S.J.; Tang, Y.P. Leonurine, a potential drug for the treatment of cardiovascular system and central nervous system diseases. Brain Behav. 2021, 11, e01995. [Google Scholar] [CrossRef] [PubMed]
- Rong, W.; Li, J.; Pan, D.; Zhou, Q.; Zhang, Y.; Lu, Q.; Wang, L.; Wang, A.; Zhu, Y.; Zhu, Q. Cardioprotective Mechanism of Leonurine against Myocardial Ischemia through a Liver-Cardiac Crosstalk Metabolomics Study. Biomolecules 2022, 12, 1512. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.M.; Guo, C.L.; Zhang, Y.F.; Chen, J.F.; Liang, Z.P.; Yang, L.H.; Ma, Y.P. Leonurine-Repressed miR-18a-5p/SOCS5/JAK2/STAT3 Axis Activity Disrupts CML malignancy. Front. Pharmacol. 2021, 12, 657724. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wei, J.; Zhou, Y.; Chen, G.; Ren, L. Leonurine hydrochloride—A new drug for the treatment of menopausal syndrome: Synthesis, estrogen-like effects and pharmacokinetics. Fitoterapia 2022, 157, 105108. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Zhang, J.B.; Liu, W.; Yao, X.R.; Guo, H.; Jin, Z.L.; Zhang, M.J.; Yuan, B.; Jiang, H.; Kim, N.H. Leonurine improves in vitro porcine embryo development competence by reducing reactive oxygen species production and protecting mitochondrial function. Theriogenology 2020, 156, 116–123. [Google Scholar] [CrossRef]
- Zheng, L.; Luo, Y.; Zhou, D.; Liu, H.; Zhou, G.; Meng, L.; Hou, Y.; Liu, C.; Li, J.; Fu, X. Leonurine improves bovine oocyte maturation and subsequent embryonic development by reducing oxidative stress and improving mitochondrial function. Theriogenology 2023, 199, 11–18. [Google Scholar] [CrossRef]
- Chi, Y.N.; Hai, D.M.; Ma, L.; Cui, Y.H.; Hu, H.T.; Liu, N.; Lan, X.B.; Yu, J.Q.; Yang, J.M. Protective effects of leonurine hydrochloride on pyroptosis in premature ovarian insufficiency via regulating NLRP3/GSDMD pathway. Int. Immunopharmacol. 2023, 114, 109520. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.Z.; Wu, W.; Zhu, Q.; Liu, X. Discovery of Leonuri and therapeutical applications: From bench to bedside. Pharmacol. Ther. 2018, 188, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Kyei, G.; Sobhani, A.; Nekonam, S.; Shabani, M.; Ebrahimi, F.; Qasemi, M.; Salahi, E.; Fardin, A. Assessing the effect of MitoQ10 and Vitamin D3 on ovarian oxidative stress, steroidogenesis and histomorphology in DHEA induced PCOS mouse model. Heliyon 2020, 6, e04279. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Yan, C.; Zhou, H.; Zhang, C.; Huang, Y.; Dong, M.; Zhang, H.; Lin, J.; Jiang, X.; Yuan, S. Brown adipose tissue activation with ginsenoside compound K ameliorates polycystic ovary syndrome. Br. J. Pharmacol. 2022, 179, 4563–4574. [Google Scholar] [CrossRef] [PubMed]
- Dou, L.; Zheng, Y.; Li, L.; Gui, X.; Chen, Y.; Yu, M.; Guo, Y. The effect of cinnamon on polycystic ovary syndrome in a mouse model. Reprod. Biol. Endocrinol. 2018, 16, 99. [Google Scholar] [CrossRef] [PubMed]
- Kamoshita, K.; Okamoto, N.; Nakajima, M.; Haino, T.; Sugimoto, K.; Okamoto, A.; Sugishita, Y.; Suzuki, N. Investigation of in vitro parameters and fertility of mouse ovary after storage at an optimal temperature and duration for transportation. Hum. Reprod. 2016, 31, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Goff, L.A.; Trapnell, C.; Kelley, D. CummeRbund: Visualization and exploration of Cufflinks high-throughput sequencing data. R Package Version 2012, 2. [Google Scholar]
- Li, A.; Zhang, J.; Zhou, Z.; Wang, L.; Liu, Y.; Liu, Y. ALDB: A domestic-animal long noncoding RNA database. PLoS ONE 2015, 10, e0124003. [Google Scholar] [CrossRef]
- Trapnell, C.; Hendrickson, D.G.; Sauvageau, M.; Goff, L.; Rinn, J.L.; Pachter, L. Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat. Biotechnol. 2013, 31, 46–53. [Google Scholar] [CrossRef]
- Zheng, J.; Wang, Z.; Yang, H.; Yao, X.; Yang, P.; Ren, C.; Wang, F.; Zhang, Y. Pituitary Transcriptomic Study Reveals the Differential Regulation of lncRNAs and mRNAs Related to Prolificacy in Different FecB Genotyping Sheep. Genes 2019, 10, 157. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Shen, H.; Jia, B.; Zhang, Y.S.; Wang, X.H.; Zeng, X.C. Differential gene expression in ovaries of Qira black sheep and Hetian sheep using RNA-Seq technique. PLoS ONE 2015, 10, e0120170. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Pearson, K. Mathematical contributions to the theory of evolution—On a form of spurious correlation which may arise when indices are used in the measurement of organs. Proc. R. Soc. Lond. 1897, 60, 489–498. [Google Scholar]
- Azziz, R.; Carmina, E.; Dewailly, D.; Diamanti-Kandarakis, E.; Escobar-Morreale, H.F.; Futterweit, W.; Janssen, O.E.; Legro, R.S.; Norman, R.J.; Taylor, A.E.; et al. Positions statement: Criteria for defining polycystic ovary syndrome as a predominantly hyperandrogenic syndrome: An Androgen Excess Society guideline. J. Clin. Endocrinol. Metab. 2006, 91, 4237–4245. [Google Scholar] [CrossRef] [PubMed]
- Rotterdam ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome (PCOS). Hum. Reprod. 2004, 19, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Lucis, O.J.; Hobkirk, R.; Hollenberg, C.H.; MacDonald, S.A.; Blahey, P. Polycystic ovaries associated with congenital adrenal hyperplasia. Can. Med. Assoc. J. 1966, 94, 1–7. [Google Scholar] [PubMed]
- Laven, J.S.; Imani, B.; Eijkemans, M.J.; Fauser, B.C. New approach to polycystic ovary syndrome and other forms of anovulatory infertility. Obstet. Gynecol. Surv. 2002, 57, 755–767. [Google Scholar] [CrossRef]
- Xue, J.; Li, X.; Liu, P.; Li, K.; Sha, L.; Yang, X.; Zhu, L.; Wang, Z.; Dong, Y.; Zhang, L. Inulin and metformin ameliorate polycystic ovary syndrome via anti-inflammation and modulating gut microbiota in mice. Endocr. J. 2019, 66, 859–870. [Google Scholar] [CrossRef]
- Li, S.Y.; Song, Z.; Song, M.J.; Qin, J.W.; Zhao, M.L.; Yang, Z.M. Impaired receptivity and decidualization in DHEA-induced PCOS mice. Sci. Rep. 2016, 6, 38134. [Google Scholar] [CrossRef] [PubMed]
- Noroozzadeh, M.; Behboudi-Gandevani, S.; Zadeh-Vakili, A.; Ramezani Tehrani, F. Hormone-induced rat model of polycystic ovary syndrome: A systematic review. Life Sci. 2017, 191, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Ji, T.; Chen, X.; Zhang, Y.; Fu, K.; Zou, Y.; Wang, W.; Zhao, J. Effects of N-Acetylcysteine on the Proliferation, Hormone Secretion Level, and Gene Expression Profiles of Goat Ovarian Granulosa Cells. Genes 2022, 13, 2306. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhou, W.; Cao, H.; Zhang, H.; Xiang, X.; Yin, Z. Ovarian Transcriptomic Analysis of Ninghai Indigenous Chickens at Different Egg-Laying Periods. Genes 2022, 13, 595. [Google Scholar] [CrossRef]
- He, D.; Liu, L.; Wang, Y.; Sheng, M. A novel genes signature associated with the progression of polycystic ovary syndrome. Pathol. Oncol. Res. 2020, 26, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; He, D.; Wang, Y.; Sheng, M. Integrated analysis of DNA methylation and transcriptome profiling of polycystic ovary syndrome. Mol. Med. Rep. 2020, 21, 2138–2150. [Google Scholar] [CrossRef]
- Ellinger, I. The Calcium-Sensing Receptor and the Reproductive System. Front. Physiol. 2016, 7, 371. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, M. Calcium at fertilization and in early development. Physiol. Rev. 2006, 86, 25–88. [Google Scholar] [CrossRef] [PubMed]
- Horner, V.L.; Wolfner, M.F. Transitioning from egg to embryo: Triggers and mechanisms of egg activation. Dev. Dyn. 2008, 237, 527–544. [Google Scholar] [CrossRef]
- Yao, J.; Ma, Y.; Zhou, S.; Bao, T.; Mi, Y.; Zeng, W.; Li, J.; Zhang, C. Metformin Prevents Follicular Atresia in Aging Laying Chickens through Activation of PI3K/AKT and Calcium Signaling Pathways. Oxid. Med. Cell. Longev. 2020, 2020, 3648040. [Google Scholar] [CrossRef]
- Richards, J.S.; Ascoli, M. Endocrine, Paracrine, and Autocrine Signaling Pathways That Regulate Ovulation. Trends Endocrinol. Metab. 2018, 29, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Jin, D.; An, X.; Zhang, Y.; Zhao, S.; Zhou, R.; Duan, Y.; Zhang, Y.; Liu, X.; Lian, F. The Potential Effect of Rhizoma coptidis on Polycystic Ovary Syndrome Based on Network Pharmacology and Molecular Docking. Evid. Based Complement. Alternat. Med. 2021, 2021, 5577610. [Google Scholar] [CrossRef] [PubMed]
- Gharani, N.; Waterworth, D.M.; Batty, S.; White, D.; Gilling-Smith, C.; Conway, G.S.; McCarthy, M.; Franks, S.; Williamson, R. Association of the steroid synthesis gene CYP11a with polycystic ovary syndrome and hyperandrogenism. Hum. Mol. Genet. 1997, 6, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Jiang, Y.; Zhao, S.; Zhao, H. Multiomics Analysis Reveals Molecular Abnormalities in Granulosa Cells of Women with Polycystic Ovary Syndrome. Front. Genet. 2021, 12, 648701. [Google Scholar] [CrossRef]
- Sagvekar, P.; Kumar, P.; Mangoli, V.; Desai, S.; Mukherjee, S. DNA methylome profiling of granulosa cells reveals altered methylation in genes regulating vital ovarian functions in polycystic ovary syndrome. Clin. Epigenet. 2019, 11, 61. [Google Scholar] [CrossRef]
- Strauss, J.F., 3rd. Some new thoughts on the pathophysiology and genetics of polycystic ovary syndrome. Ann. N. Y. Acad. Sci. 2003, 997, 42–48. [Google Scholar] [CrossRef]
- Wang, W.; Zheng, J.; Cui, N.; Jiang, L.; Zhou, H.; Zhang, D.; Hao, G. Baicalin ameliorates polycystic ovary syndrome through AMP-activated protein kinase. J. Ovarian Res. 2019, 12, 109. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhao, H.; Li, T.; Zhang, W.; Wu, K.; Li, M.; Bian, Y.; Liu, H.; Ning, Y.; Li, G.; et al. Hypomethylation of the LH/choriogonadotropin receptor promoter region is a potential mechanism underlying susceptibility to polycystic ovary syn-drome. Endocrinology 2014, 155, 1445–1452. [Google Scholar] [CrossRef] [PubMed]
- Kanamarlapudi, V.; Gordon, U.D.; López Bernal, A. Luteinizing hormone/chorionic gonadotrophin receptor overexpressed in granulosa cells from polycystic ovarysyndrome ovaries is functionally active. Reprod. Biomed. Online 2016, 32, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.R.; Brower, M.A.; Xu, N.; Cui, J.; Mengesha, E.; Chen, Y.D.; Taylor, K.D.; Azziz, R.; Goodarzi, M.O. Systems Genetics Reveals the Functional Context of PCOS Loci and Identifies Genetic and Molecular Mechanisms of Disease Heterogeneity. PLoS Genet. 2015, 11, e1005455. [Google Scholar] [CrossRef]
- Sutton-McDowall, M.L.; Gilchrist, R.B.; Mpson, J.G. The pivotal role of glucose metabolism in determining oocyte developmental competence. Reproduction 2010, 139, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Yang, L.; Sun, G.; Shao, Y.; Liu, Y.; Yang, H.; Wang, Y.; Zhang, M.; Shang, Y.; Gu, X. Assessment of the Effect of Leonurine Hydrochloride in a Mouse Model of PCOS by Gene Expression Profiling. Genes 2024, 15, 507. https://doi.org/10.3390/genes15040507
Wang M, Yang L, Sun G, Shao Y, Liu Y, Yang H, Wang Y, Zhang M, Shang Y, Gu X. Assessment of the Effect of Leonurine Hydrochloride in a Mouse Model of PCOS by Gene Expression Profiling. Genes. 2024; 15(4):507. https://doi.org/10.3390/genes15040507
Chicago/Turabian StyleWang, Mengmeng, Li Yang, Guojie Sun, Yongbin Shao, Yuran Liu, Huiying Yang, Yan Wang, Mengyuan Zhang, Yunxia Shang, and Xinli Gu. 2024. "Assessment of the Effect of Leonurine Hydrochloride in a Mouse Model of PCOS by Gene Expression Profiling" Genes 15, no. 4: 507. https://doi.org/10.3390/genes15040507
APA StyleWang, M., Yang, L., Sun, G., Shao, Y., Liu, Y., Yang, H., Wang, Y., Zhang, M., Shang, Y., & Gu, X. (2024). Assessment of the Effect of Leonurine Hydrochloride in a Mouse Model of PCOS by Gene Expression Profiling. Genes, 15(4), 507. https://doi.org/10.3390/genes15040507