
Genome-Wide Analysis of the Lateral Organ Boundaries Domain (LBD) Gene Family in Sweet Potato (Ipomoea batatas)

Abstract
1. Introduction
2. Materials and Methods
2.1. Identification and Physicochemical Properties of IbLBD Gene Family Members
2.2. Gene Structure and Conserved Motif Analysis
2.3. Phylogenetic Analysis of IbLBD Proteins
2.4. Chromosome Localization and Duplication Events
2.5. Cis-Acting Elements Analysis of IbLBD Genes
2.6. Plant Material and Treatments
2.7. Protein–Protein Interaction Network and GO Annotation Analysis of IbLBDs
3. Results
3.1. Identification of LBD Genes in Sweet Potato
3.2. Motif Compositions and Gene Structure of the LBD Gene Family in Sweet Potato
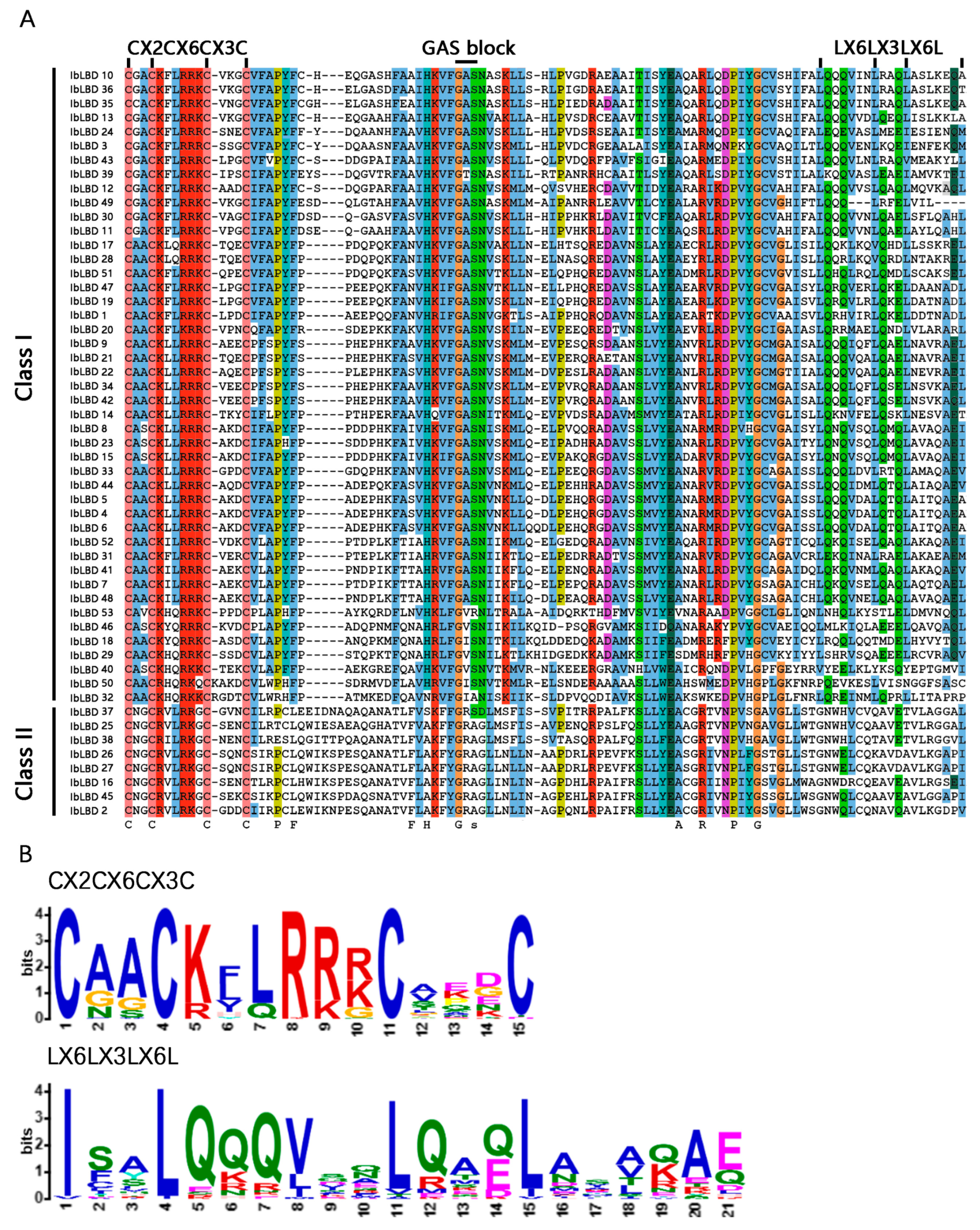
3.3. Conserved Sequence Alignment of LBD Gene Family in Sweet Potato
3.4. Phylogenetic Analysis
3.5. Chromosome Locations and Gene Duplication Analysis
3.6. Evolutionary Analysis of the IbLBD Genes
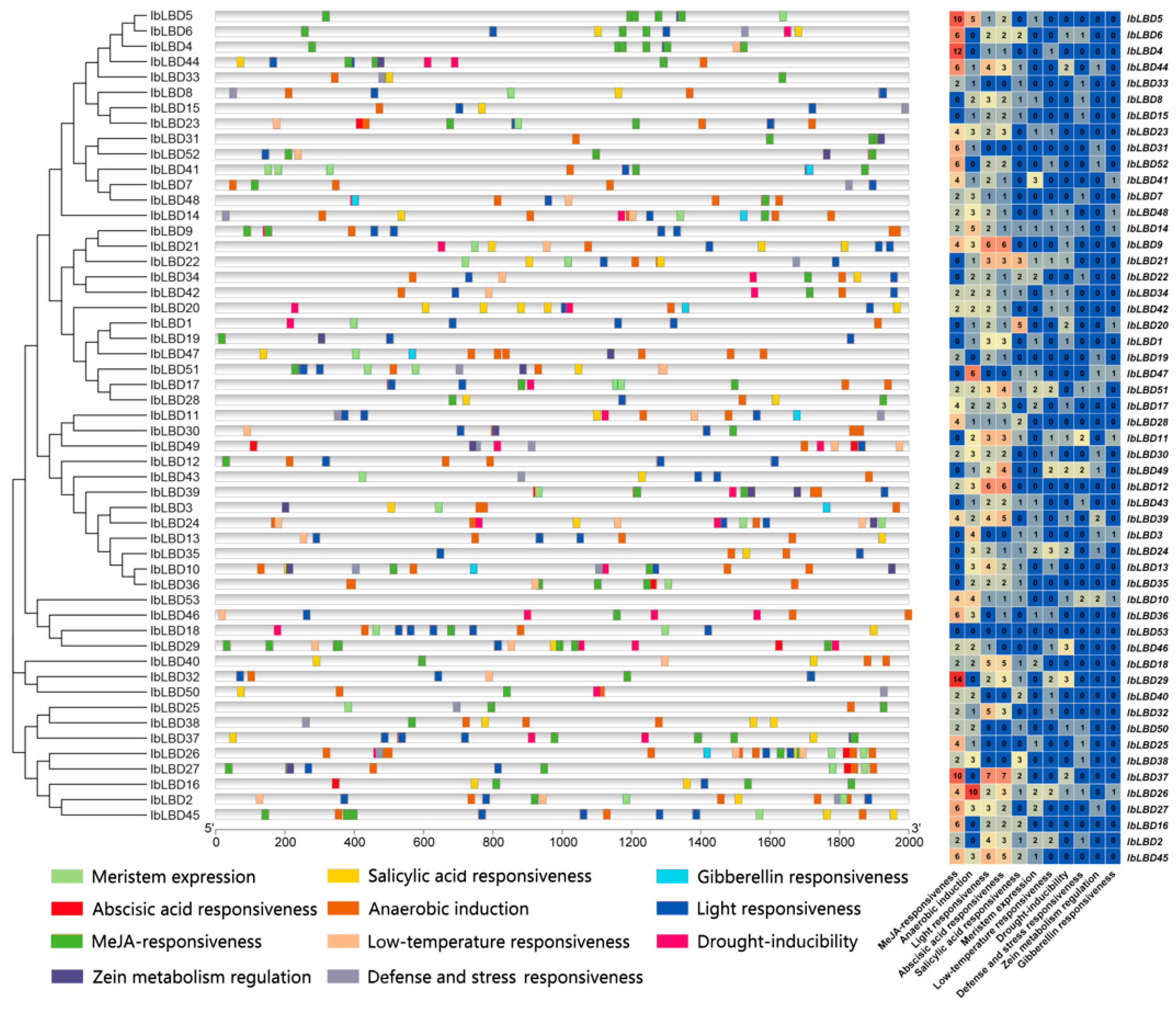
3.7. Analysis of Cis-Regulatory Element Distribution in IbLBD Promoters
3.8. Expression Patterns of IbLBD Gene Family in Sweet Potato
3.9. Quantitative qPCR Analysis of IbLBD Genes in Different Tissues
3.10. Quantitative qPCR Analysis of IbLBD Genes under Abiotic Stresses
3.11. Regulatory Network in Sweet Potato
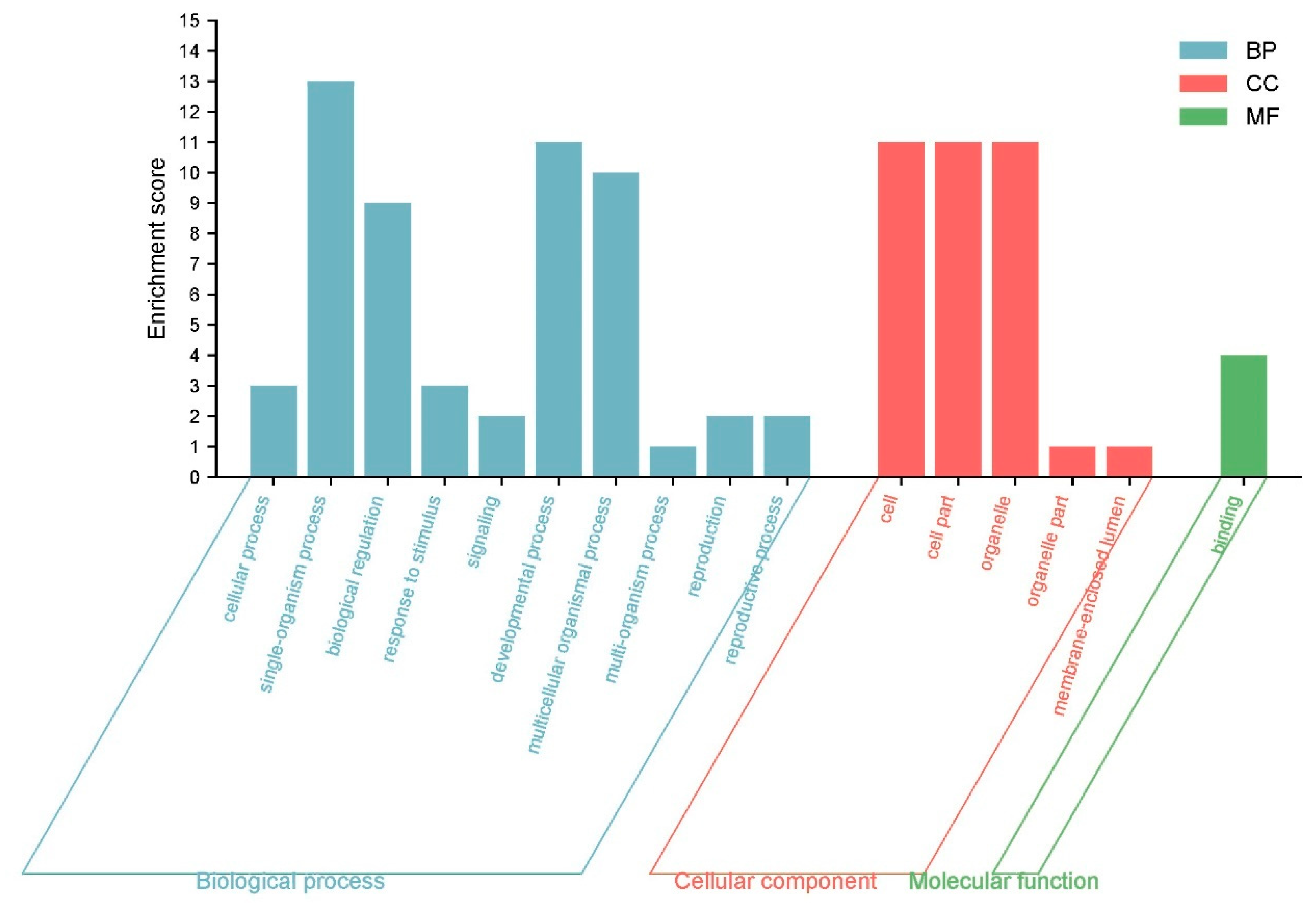
3.12. GO Annotation and Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Li, Z.; Ma, B.; Hou, Q.; Wan, X. Phylogeny and Functions of LOB Domain Proteins in Plants. Int. J. Mol. Sci. 2020, 21, 2278. [Google Scholar] [CrossRef]
- Liang, J.; Hou, Z.; Liao, J.; Qin, Y.; Wang, L.; Wang, X.; Su, W.; Cai, Z.; Fang, Y.; Aslam, M.; et al. Genome-Wide Identification and Expression Analysis of LBD Transcription Factor Genes in Passion Fruit (Passiflora edulis). Int. J. Mol. Sci. 2022, 23, 4700. [Google Scholar] [CrossRef] [PubMed]
- Majer, C.; Hochholdinger, F. Defining the boundaries: Structure and function of LOB domain proteins. Trends Plant Sci. 2011, 16, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Fang, G.Y.; He, H.; Chen, J. Genome-Wide Identification, Evolutionary Analysis and Expression Profiles of Lateral Organ Boundaries Domain Gene Family in Lotus japonicus and Medicago truncatula. PLoS ONE 2016, 11, e0161901. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Wang, J.; Lin, S.; Tian, Z.; Zhou, K.; Luan, H.; Lyu, C.; Zhang, X.; Xu, R. A genome-wide analysis of the Asymmetric Leaves2/Lateral Organ Boundaries (AS2/LOB) gene family in barley (Hordeum vulgare L.). J. Zhejiang Univ. Sci. B 2016, 17, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Chanderbali, A.S.; He, F.; Soltis, P.S.; Soltis, D.E. Out of the Water: Origin and Diversification of the LBD Gene Family. Mol. Biol. Evol. 2015, 32, 1996–2000. [Google Scholar] [CrossRef] [PubMed]
- Shuai, B.; Reynaga-Pena, C.G.; Springer, P.S. The lateral organ boundaries gene defines a novel, plant-specific gene family. Plant Physiol. 2002, 129, 747–761. [Google Scholar] [CrossRef]
- Zhao, D.; Chen, P.; Chen, Z.; Zhang, L.; Wang, Y.; Xu, L. Genome-wide analysis of the LBD family in rice: Gene functions, structure and evolution. Comput. Biol. Med. 2023, 153, 106452. [Google Scholar] [CrossRef]
- Taramino, G.; Sauer, M.; Stauffer, J.L., Jr.; Multani, D.; Niu, X.; Sakai, H.; Hochholdinger, F. The maize (Zea mays L.) RTCS gene encodes a LOB domain protein that is a key regulator of embryonic seminal and post-embryonic shoot-borne root initiation. Plant J. 2007, 50, 649–659. [Google Scholar] [CrossRef]
- Zhang, X.; He, Y.; He, W.; Su, H.; Wang, Y.; Hong, G.; Xu, P. Structural and functional insights into the LBD family involved in abiotic stress and flavonoid synthases in Camellia sinensis. Sci. Rep. 2019, 9, 15651. [Google Scholar] [CrossRef]
- Yu, Q.; Hu, S.; Du, J.; Yang, Y.; Sun, X. Genome-wide identification and characterization of the lateral organ boundaries domain gene family in Brassica rapa var. rapa. Plant Divers. 2019, 42, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Shao, F.; Macmillan, C.; Wilson, I.W.; van der Merwe, K.; Hussey, S.G.; Myburg, A.A.; Dong, X.; Qiu, D. Genomewide analysis of the lateral organ boundaries domain gene family in Eucalyptus grandis reveals members that differentially impact secondary growth. Plant Biotechnol. J. 2018, 16, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Shi, G.; Du, H.; Wang, H.; Zhang, Z.; Hu, D.; Wang, J.; Huang, F.; Yu, D. Genome-Wide Analysis of Soybean Lateral Organ Boundaries Domain-Containing Genes: A Functional Investigation of GmLBD12. Plant Genome 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, R.; Cheng, Y.; Lei, P.; Song, W.; Zheng, W.; Nie, X. Genome-wide Identification Evolution and Expression Analysis of LBD Transcription Factor Family in Bread Wheat (Triticum aestivum L.). Front. Plant Sci. 2021, 12, 721253. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.; Jeon, E.; Pandey, S.K.; Ha, S.H.; Kim, J. LBD13 positively regulates lateral root formation in Arabidopsis. Planta 2019, 249, 1251–1258. [Google Scholar] [CrossRef] [PubMed]
- Okushima, Y.; Fukaki, H.; Onoda, M.; Theologis, A.; Tasaka, M. Arf7 and arf19 regulate lateral root formation via direct activation of LBD/ASL genes in Arabidopsis. Plant Cell 2007, 19, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Iwakawa, H.; Iwasaki, M.; Kojima, S.; Ueno, Y.; Soma, T.; Tanaka, H.; Semiarti, E.; Machida, Y.; Machida, C. Expression of the asymmetric leaves 2 gene in the adaxial domain of Arabidopsis leaves represses cell proliferation in this domain and is critical for the development of properly expanded leaves. Plant J. 2007, 51, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Soyano, T.; Thitamadee, S.; Machida, Y.; Chua, N.H. Asymmetric Leaves2-Like19/Lateral Organ Boundaries DOMAIN30 and ASL20/LBD18 Regulate Tracheary Element Differentiation in Arabidopsis. Plant Cell 2008, 20, 3359–3373. [Google Scholar] [CrossRef]
- Lee, K.H.; Du, Q.; Zhuo, C.; Qi, L.; Wang, H. LBD29-involved auxin signaling represses nac master regulators and fiber wall biosynthesis. Plant Physiol. 2019, 181, 595–608. [Google Scholar] [CrossRef]
- Thatcher, L.F.; Powell, J.J.; Aitken, E.A.B.; Kazan, K.; Manners, J.M. The lateral organ boundaries domain transcription factor LBD20 functions in fusarium wilt susceptibility and jasmonate signaling in Arabidopsis. Plant Physiol. 2012, 160, 407–418. [Google Scholar] [CrossRef]
- Montano, M.M.; Muller, V.; Trobaugh, A.; Katzenellenbogen, B.S. The carboxy-terminal F domain of the human estrogen receptor: Role in the transcriptional activity of the receptor and the effectiveness of antiestrogens as estrogen antagonists. Mol. Endocrinol. 1995, 9, 814–825. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Patel, S.R.; Skafar, D.F. Modulation of nuclear receptor activity by the F domain. Mol. Cell. Endocrinol. 2015, 418, 298–305. [Google Scholar] [CrossRef]
- Rubin, G.; Tohge, T.; Matsuda, F.; Saito, K.; Scheible, W.R. Members of the LBD Family of Transcription Factors Repress Anthocyanin Synthesis and Affect Additional Nitrogen Responses in Arabidopsis. Plant Cell 2009, 21, 3567–3584. [Google Scholar] [CrossRef] [PubMed]
- Goh, T.; Toyokura, K.; Yamaguchi, N.; Okamoto, Y.; Uehara, T.; Kaneko, S.; Takebayashi, Y.; Kasahara, H.; Ikeyama, Y.; Okushima, Y.; et al. Lateral root initiation requires the sequential induction of transcription factors LBD16 and PUCHI in Arabidopsis thaliana. New Phytol. 2019, 224, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Kim, M.J.; Kim, N.Y.; Lee, S.H.; Kim, J. LBD18 acts as a transcriptional activator that directly binds to the expansin14 promoter in promoting lateral root emergence of Arabidopsis. Plant J. 2013, 73, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, L.F.; Kazan, K.; Manners, J.M. Lateral organ boundaries domain transcription factors: New roles in plant defense. Plant Signal Behav. 2012, 7, 1702–1704. [Google Scholar] [CrossRef] [PubMed]
- Mangeon, A.; Bell, E.M.; Lin, W.C.; Jablonska, B.; Springer, P.S. Misregulation of the LOB domain gene DDA1 suggests possible functions in auxin signalling and photomorphogenesis. J. Exp. Bot. 2011, 62, 221–233. [Google Scholar] [CrossRef]
- Liu, H.; Cao, M.; Chen, X.; Ye, M.; Zhao, P.; Nan, Y.; Li, W.; Zhang, C.; Kong, L.; Kong, N.; et al. Genome-wide analysis of the lateral organ boundaries domain (LBD) gene family in Solanum tuberosum. Int. J. Mol. Sci. 2019, 20, 5360. [Google Scholar] [CrossRef]
- Yu, J.; Xie, Q.; Li, C.; Dong, Y.; Zhu, S.; Chen, J. Comprehensive characterization and gene expression patterns of LBD gene family in Gossypium. Planta 2020, 251, 81. [Google Scholar] [CrossRef]
- Huang, X.; Liu, G.; Zhang, W. Genome-wide analysis of LBD (LATERAL ORGAN BOUNDARIES Domain) gene family in Brassica rapa. Braz. Arch. Biol. Technol. 2018, 61. [Google Scholar] [CrossRef]
- Xiong, J.; Zhang, W.; Zheng, D.; Xiong, H.; Feng, X.; Zhang, X.; Wang, Q.; Wu, F.; Xu, J.; Lu, Y. ZmLBD5 Increases drought sensitivity by suppressing ROS accumulation in Arabidopsis. Plants 2022, 11, 1382. [Google Scholar] [CrossRef] [PubMed]
- Teng, R.; Yang, N.; Liu, C.; Chen, Y.; Wang, Y.; Zhuang, J. CsLBD37, a LBD/ASL transcription factor, affects nitrate response and flowering of tea plant. Sci. Hortic. 2022, 306, 111457. [Google Scholar] [CrossRef]
- Mao, Y.; Abdoulaye, A.H.; Song, J.; Yao, X.; Zhang, Y.; Gao, Y.; Ye, Y.; Luo, K.; Xia, W.; Chen, Y. Genome-wide analyses of LATERAL ORGAN BOUNDARIES in cassava reveal the role of LBD47 in defence against bacterial blight. PLoS ONE 2023, 18, e0282100. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; He, W.; Wang, L.; Zhang, X.; Wang, K.; Xiang, Y. PheLBD29, an LBD transcription factor from Moso bamboo, causes leaf curvature and enhances tolerance to drought stress in transgenic Arabidopsis. J. Plant Physiol. 2023, 280, 153865. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Shi, J.; Hu, Y.; Li, D.; Guo, L.; Zhao, Z.; Lee, G.S.; Qiao, Y. Genome-Wide Analysis of Soybean Lateral Organ Boundaries Domain Gene Family Reveals the Role in Phytophthora Root and Stem Rot. Front. Plant Sci. 2022, 13, 865165. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhang, P.; Zhu, Y.; Lou, Q.; He, S. Antioxidant and prebiotic activity of five peonidin-based anthocyanins extracted from purple sweet potato (Ipomoea batatas (L.) Lam.). Sci. Rep. 2018, 8, 5018. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q. Improvement for agronomically important traits by gene engineering in sweetpotato. Breed Sci. 2017, 67, 15–26. [Google Scholar] [CrossRef]
- Hou, F.; Du, T.; Qin, Z.; Xu, T.; Li, A.; Dong, S.; Ma, D.; Li, Z.; Wang, Q.; Zhang, L. Genome-wide in silico identification and expression analysis of β-galactosidase family members in sweetpotato [Ipomoea batatas (L.) Lam]. BMC Genom. 2021, 22, 140. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Krejci, A.; Hupp, T.R.; Lexa, M.; Vojtesek, B.; Muller, P. Hammock: A hidden Markov model-based peptide clustering algorithm to identify protein-interaction consensus motifs in large datasets. Bioinformatics 2016, 32, 9–16. [Google Scholar] [CrossRef]
- Schultz, J.; Copley, R.R.; Doerks, T.; Ponting, C.P.; Bork, P. SMART: A web-based tool for the study of genetically mobile domains. Nucleic Acids Res. 2000, 28, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. Meme Suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Yang, X.; Zhou, T.; Wang, M.; Li, T.; Wang, G.; Fu, F.F.; Cao, F. Systematic investigation and expression profiles of the GbR2R3-MYB transcription factor family in ginkgo (Ginkgo biloba L.). Int. J. Biol. Macromol. 2021, 172, 250–262. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Ding, N.; Wang, A.; Zhang, X.; Wu, Y.; Wang, R.; Cui, H.; Huang, R.; Luo, Y. Identification and analysis of glutathione S-transferase gene family in sweet potato reveal divergent GST-mediated networks in aboveground and underground tissues in response to abiotic stresses. BMC Plant Biol. 2017, 17, 225. [Google Scholar] [CrossRef]
- Otasek, D.; Morris, J.H.; Boucas, J.; Pico, A.R.; Demchak, B. Cytoscape Automation: Empowering workflow-based network analysis. Genome Biol. 2019, 20, 185. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, X.; Su, L.; Sun, Y.; Zhang, S.; Hao, Y.; You, C. Identification, evolution and expression analysis of the LBD gene family in tomato. Sci. Agric. Sin. 2013, 46, 2501–2513. [Google Scholar] [CrossRef]
- Yang, J.; Moeinzadeh, M.H.; Kuhl, H.; Helmuth, J.; Xiao, P.; Haas, S.; Liu, G.; Zheng, J.; Sun, Z.; Fan, W.; et al. Haplotype-resolved sweet potato genome traces back its hexaploidization history. Nat. Plants 2017, 3, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Huang, Z.; Ma, R.; Ramakrishnan, M.; Chen, J.; Zhang, Z.; Yrjala, K. Genome-wide identification and expression analysis of LBD transcription factor genes in Moso bamboo (Phyllostachys edulis). BMC Plant Biol. 2021, 21, 296. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Tang, Z.; Li, X.; Li, J.; Zhang, M.; Zhao, K.; Liu, H.; Zhang, S.; Wu, J. Mining and evolution analysis of lateral organ boundaries domain (LBD) genes in Chinese white pear (Pyrus bretschneideri). BMC Genom. 2020, 21, 644. [Google Scholar] [CrossRef]
- Du, Y.; Zhao, Q.; Li, W.; Geng, J.; Li, S.; Yuan, X.; Gu, Y.; Zhong, J.; Zhang, Y.; Du, J. Genome-wide identification of the LBD transcription factor genes in common bean (Phaseolus vulgaris L.) and expression analysis under different abiotic stresses. J. Plant Interact. 2022, 17, 731–743. [Google Scholar] [CrossRef]
- Chalfun, A.; Franken, J.; Mes, J.; Marsch-Martinez, N.; Pereira, A.; Angenent, G. ASYMMETRIC LEAVES2-LIKE1 gene, a member of the AS2/LOB family, controls proximal-distal patterning in Arabidopsis petals. Plant Mol. Biol. 2005, 57, 559–575. [Google Scholar] [CrossRef]
- Mangeon, A.; Lin, W.; Springer, P. Functional divergence in the Arabidopsis LOB domain gene family. Plant Signal. Behav. 2012, 7, 1544–1547. [Google Scholar] [CrossRef]
- Guo, Z.; Xu, H.; Lei, Q.; Du, J.; Li, C.; Wang, C.; Yang, Y.; Yang, Y.; Sun, X. The Arabidopsis transcription factor LBD15 mediates ABA signaling and tolerance of water-deficit stress by regulating ABI4 expression. Plant J. 2020, 104, 510–521. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Chromosome Location | Size (aa) | Mw (kD) | pI | GRAVY | Predicted Location | |
|---|---|---|---|---|---|---|---|---|
| IbLBD1 | OR133645 | 1687306 | 1688873 | 188 | 20.72 | 6.5 | −0.311 | Nucleus |
| IbLBD2 | OR133646 | 8494464 | 8496120 | 249 | 27.22 | 6.6 | −0.447 | Nucleus |
| IbLBD3 | OR133647 | 27014760 | 27015755 | 209 | 23.87 | 5.21 | −0.431 | Nucleus |
| IbLBD4 | OR133648 | 271534 | 275058 | 174 | 19.07 | 8.97 | −0.411 | Nucleus |
| IbLBD5 | OR133649 | 1039868 | 1043206 | 167 | 18.27 | 7.72 | −0.411 | Nucleus |
| IbLBD6 | OR133650 | 1069879 | 1074143 | 191 | 21.44 | 10.08 | −0.717 | Nucleus |
| IbLBD7 | OR133651 | 3938571 | 3939920 | 215 | 23.25 | 6.14 | −0.039 | Nucleus |
| IbLBD8 | OR133652 | 5639554 | 5640802 | 191 | 20.68 | 6.99 | −0.117 | Nucleus |
| IbLBD9 | OR133653 | 5829105 | 5830345 | 242 | 25.65 | 7.66 | 0.173 | Nucleus |
| IbLBD10 | OR133654 | 7327214 | 7328603 | 236 | 26.03 | 6.2 | −0.534 | Nucleus |
| IbLBD11 | OR133655 | 36568489 | 36573862 | 334 | 35.22 | 9.28 | −0.256 | Nucleus |
| IbLBD12 | OR133656 | 1456672 | 1458605 | 214 | 23.11 | 8.36 | −0.817 | Nucleus |
| IbLBD13 | OR133657 | 1466969 | 1469187 | 225 | 24.32 | 6.48 | −0.534 | Nucleus |
| IbLBD14 | OR133658 | 3005283 | 3007545 | 160 | 17.77 | 6.71 | 0.322 | Nucleus |
| IbLBD15 | OR133659 | 3278659 | 3280029 | 172 | 18.94 | 7.63 | −0.211 | Nucleus |
| IbLBD16 | OR133660 | 22684760 | 22686086 | 224 | 24.46 | 8.6 | −0.628 | Nucleus |
| IbLBD17 | OR133661 | 4128151 | 4129743 | 277 | 31.01 | 6.16 | 0.000 | Nucleus |
| IbLBD18 | OR133662 | 5451750 | 5452974 | 226 | 25.48 | 4.68 | −0.700 | Nucleus |
| IbLBD19 | OR133663 | 5662166 | 5664439 | 197 | 22.23 | 8.15 | −0.278 | Nucleus |
| IbLBD20 | OR133664 | 27801512 | 27802278 | 180 | 19.99 | 8.28 | −0.445 | Nucleus |
| IbLBD21 | OR133665 | 27967462 | 27968594 | 214 | 23.51 | 8.6 | −0.445 | Nucleus |
| IbLBD22 | OR133666 | 1121586 | 1126383 | 246 | 26.19 | 8.67 | 0.122 | Nucleus |
| IbLBD23 | OR133667 | 1261129 | 1262521 | 212 | 23.44 | 5.79 | −0.584 | Nucleus |
| IbLBD24 | OR133668 | 2016696 | 2019488 | 307 | 34.00 | 4.58 | −0.080 | Nucleus |
| IbLBD25 | OR133669 | 27558292 | 27560598 | 187 | 20.18 | 9.27 | −0.284 | Nucleus |
| IbLBD26 | OR133670 | 28842217 | 28843398 | 204 | 21.82 | 8.17 | −0.417 | Nucleus |
| IbLBD27 | OR133671 | 29266026 | 29267217 | 236 | 25.74 | 8.52 | −0.528 | Nucleus |
| IbLBD28 | OR133672 | 33791108 | 33797053 | 326 | 36.47 | 6.66 | −0.462 | Nucleus |
| IbLBD29 | OR133673 | 34877261 | 34878276 | 210 | 23.58 | 6.65 | −0.745 | Nucleus |
| IbLBD30 | OR133674 | 850688 | 852583 | 218 | 22.77 | 8.47 | −0.250 | Nucleus |
| IbLBD31 | OR133675 | 4610655 | 4611562 | 149 | 16.16 | 8.75 | −0.017 | Nucleus |
| IbLBD32 | OR133676 | 3472142 | 3475757 | 322 | 36.23 | 9.58 | −0.739 | Nucleus |
| IbLBD33 | OR133677 | 6217307 | 6221397 | 366 | 39.29 | 4.59 | −0.306 | Nucleus |
| IbLBD34 | OR133678 | 14820911 | 14822874 | 214 | 23.46 | 9.11 | −0.261 | Nucleus |
| IbLBD35 | OR133679 | 4501947 | 4503162 | 226 | 24.89 | 5.39 | −0.273 | Nucleus |
| IbLBD36 | OR133680 | 4532985 | 4534460 | 245 | 27.27 | 4.97 | −0.233 | Nucleus |
| IbLBD37 | OR133681 | 3579351 | 3580312 | 209 | 22.47 | 8.67 | −0.101 | Nucleus |
| IbLBD38 | OR133682 | 13130843 | 13131707 | 201 | 22.17 | 7.64 | −0.336 | Nucleus |
| IbLBD39 | OR133683 | 22497885 | 22500062 | 240 | 26.20 | 5.47 | −0.062 | Nucleus |
| IbLBD40 | OR133684 | 39950925 | 39952093 | 223 | 25.26 | 8.77 | −0.966 | Nucleus |
| IbLBD41 | OR133685 | 8697591 | 8699166 | 202 | 21.89 | 6.11 | −0.084 | Nucleus |
| IbLBD42 | OR133686 | 9909480 | 9911649 | 214 | 23.43 | 9.25 | −0.284 | Nucleus |
| IbLBD43 | OR133687 | 28316625 | 28318692 | 175 | 18.91 | 6.71 | −0.007 | Nucleus |
| IbLBD44 | OR133688 | 29434436 | 29438687 | 183 | 19.95 | 8.58 | −0.578 | Nucleus |
| IbLBD45 | OR133689 | 11950627 | 11951867 | 280 | 30.17 | 5.64 | −1.033 | Nucleus |
| IbLBD46 | OR133690 | 18216174 | 18217557 | 314 | 34.94 | 5.46 | −0.580 | Nucleus |
| IbLBD47 | OR133691 | 19821743 | 19824082 | 185 | 20.18 | 6.88 | −0.728 | Nucleus |
| IbLBD48 | OR133692 | 291788 | 295234 | 475 | 52.48 | 6.25 | 0.084 | Nucleus |
| IbLBD49 | OR133693 | 8713728 | 8714718 | 157 | 17.29 | 9.97 | 0.361 | Nucleus |
| IbLBD50 | OR133694 | 26782324 | 26785743 | 520 | 57.51 | 8.81 | −0.730 | Nucleus |
| IbLBD51 | OR133695 | 5492804 | 5494838 | 227 | 24.66 | 8.56 | −0.495 | Nucleus |
| IbLBD52 | OR133696 | 5850533 | 5851542 | 213 | 22.89 | 5.47 | −0.056 | Nucleus |
| IbLBD53 | OR133697 | 18062560 | 18063739 | 277 | 31.14 | 9.69 | −0.403 | Nucleus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, L.; Lin, X.; Tang, B.; Zhao, R.; Wang, Y.; Lin, Y.; Wu, L.; Zheng, C.; Zhu, H. Genome-Wide Analysis of the Lateral Organ Boundaries Domain (LBD) Gene Family in Sweet Potato (Ipomoea batatas). Genes 2024, 15, 237. https://doi.org/10.3390/genes15020237
Shi L, Lin X, Tang B, Zhao R, Wang Y, Lin Y, Wu L, Zheng C, Zhu H. Genome-Wide Analysis of the Lateral Organ Boundaries Domain (LBD) Gene Family in Sweet Potato (Ipomoea batatas). Genes. 2024; 15(2):237. https://doi.org/10.3390/genes15020237
Chicago/Turabian StyleShi, Lei, Xiongjian Lin, Binquan Tang, Rong Zhao, Yichi Wang, Yingyi Lin, Liangliang Wu, Chao Zheng, and Hongbo Zhu. 2024. "Genome-Wide Analysis of the Lateral Organ Boundaries Domain (LBD) Gene Family in Sweet Potato (Ipomoea batatas)" Genes 15, no. 2: 237. https://doi.org/10.3390/genes15020237
APA StyleShi, L., Lin, X., Tang, B., Zhao, R., Wang, Y., Lin, Y., Wu, L., Zheng, C., & Zhu, H. (2024). Genome-Wide Analysis of the Lateral Organ Boundaries Domain (LBD) Gene Family in Sweet Potato (Ipomoea batatas). Genes, 15(2), 237. https://doi.org/10.3390/genes15020237