
Exploring Multi-Tissue Alternative Splicing and Skeletal Muscle Metabolism Regulation in Obese- and Lean-Type Pigs
 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Identification and Quantification of Transcripts
2.3. Analysis of Differentially Expressed Genes, Transcripts and Alternative Splicing
2.4. RNA Extractison
2.5. Semi-Quantitative RT-PCR Analysis of Alternative Splicing Events
2.6. Statistical Analysis
3. Results
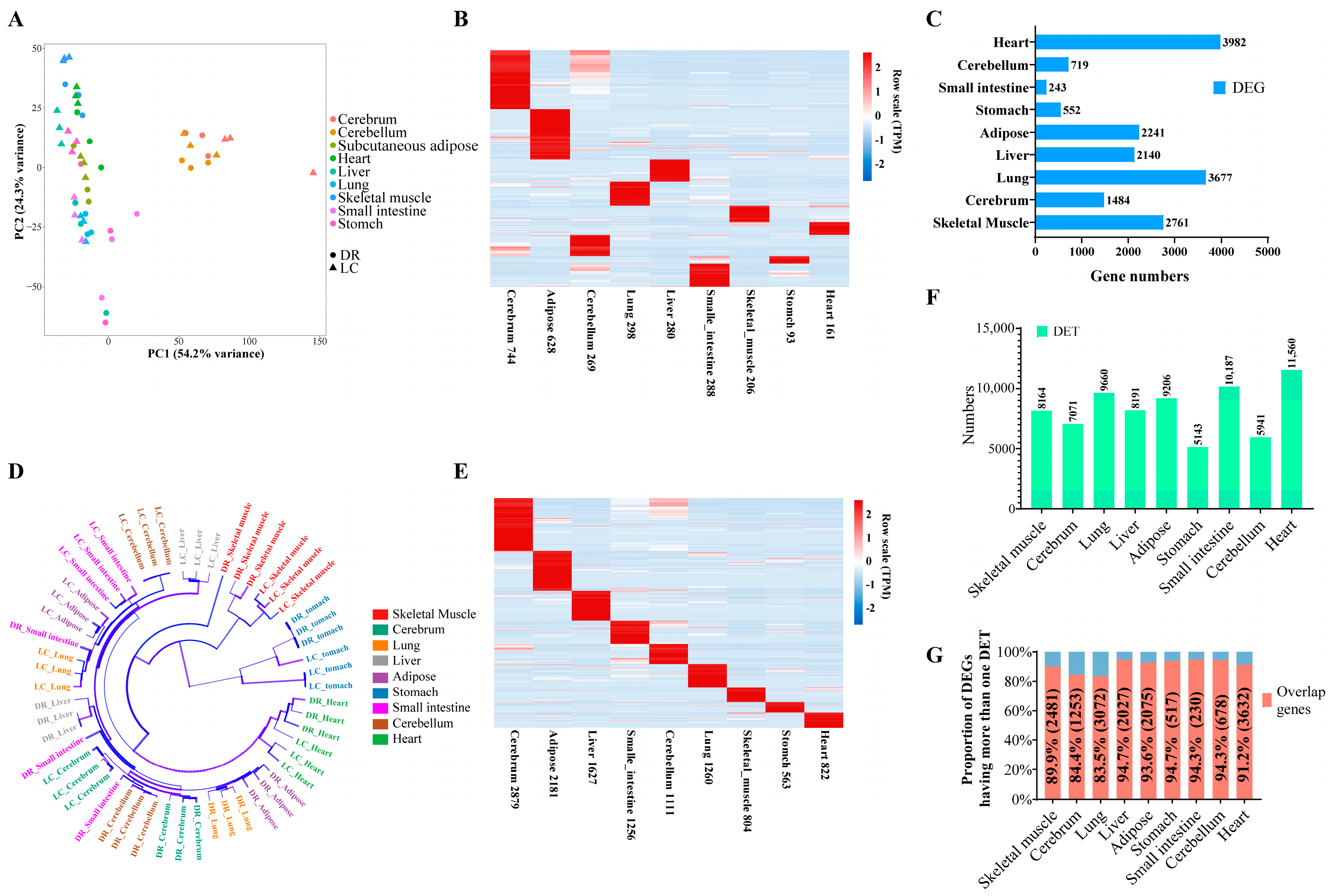
3.1. Tissue- and Breed-Specific Differential Expression Profiles of Lean- and Obese-Type Pigs
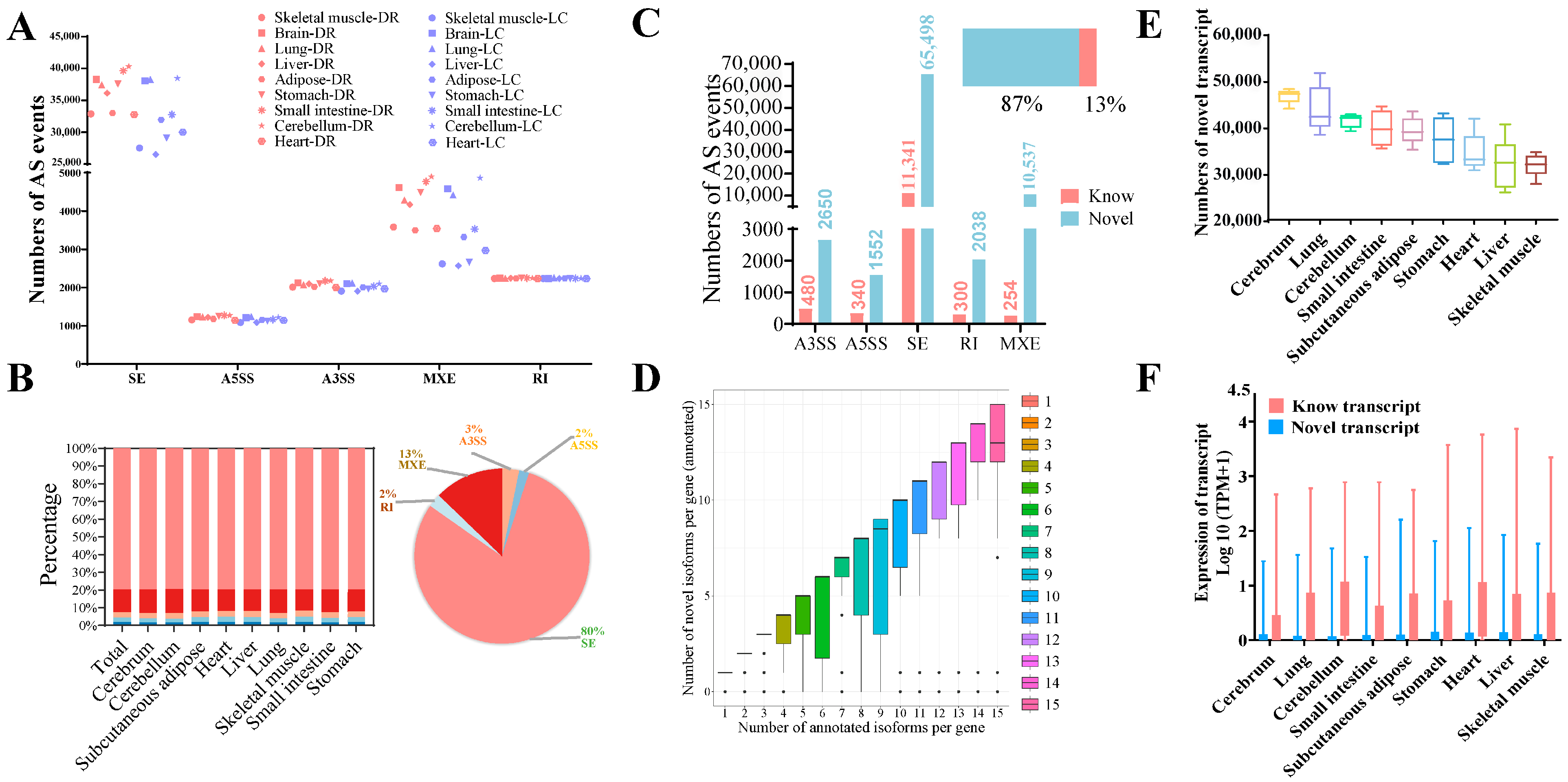
3.2. Five Types of Alternative Splicing Events Generate Novel Transcripts
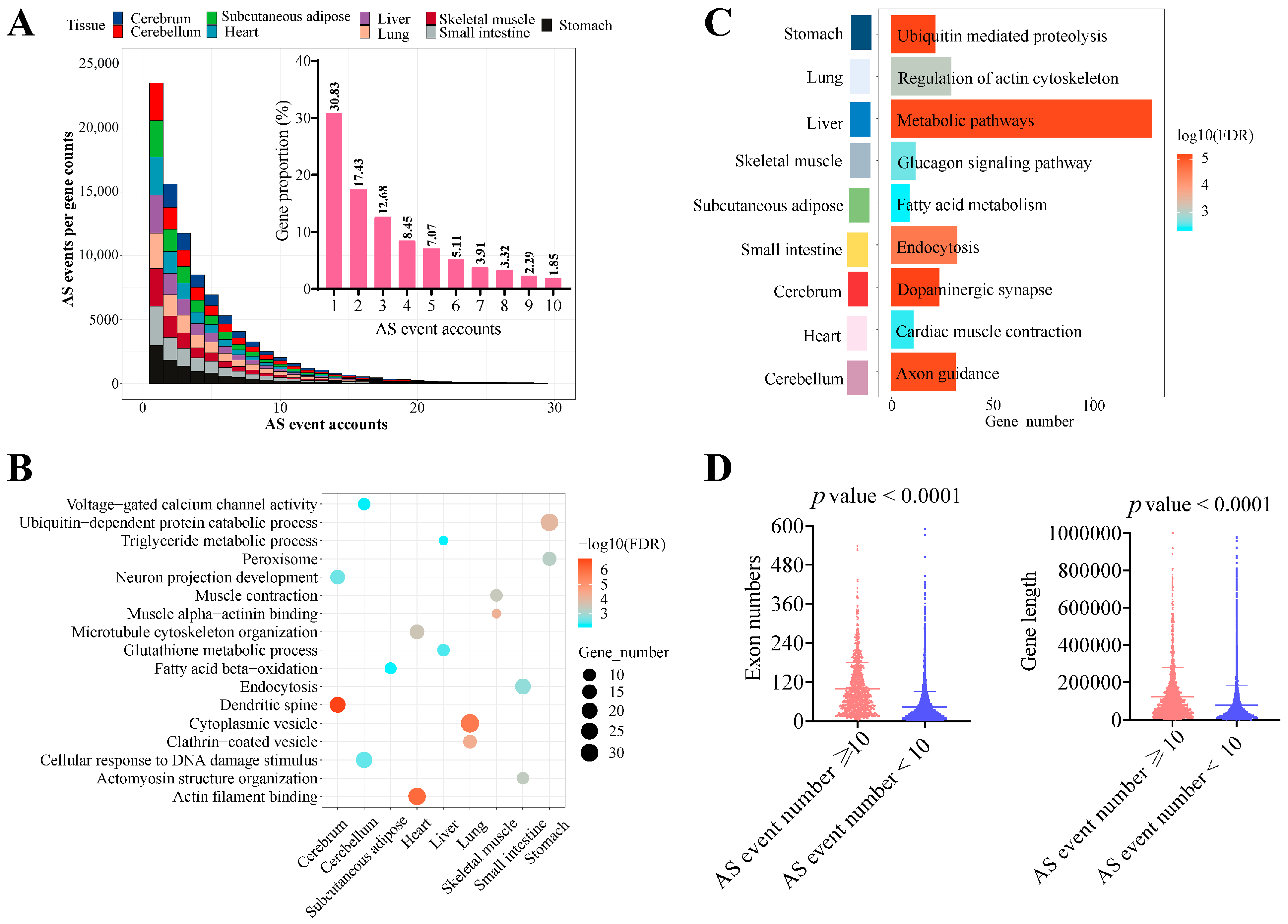
3.3. Tissue-Specific Alternative Splicing Events
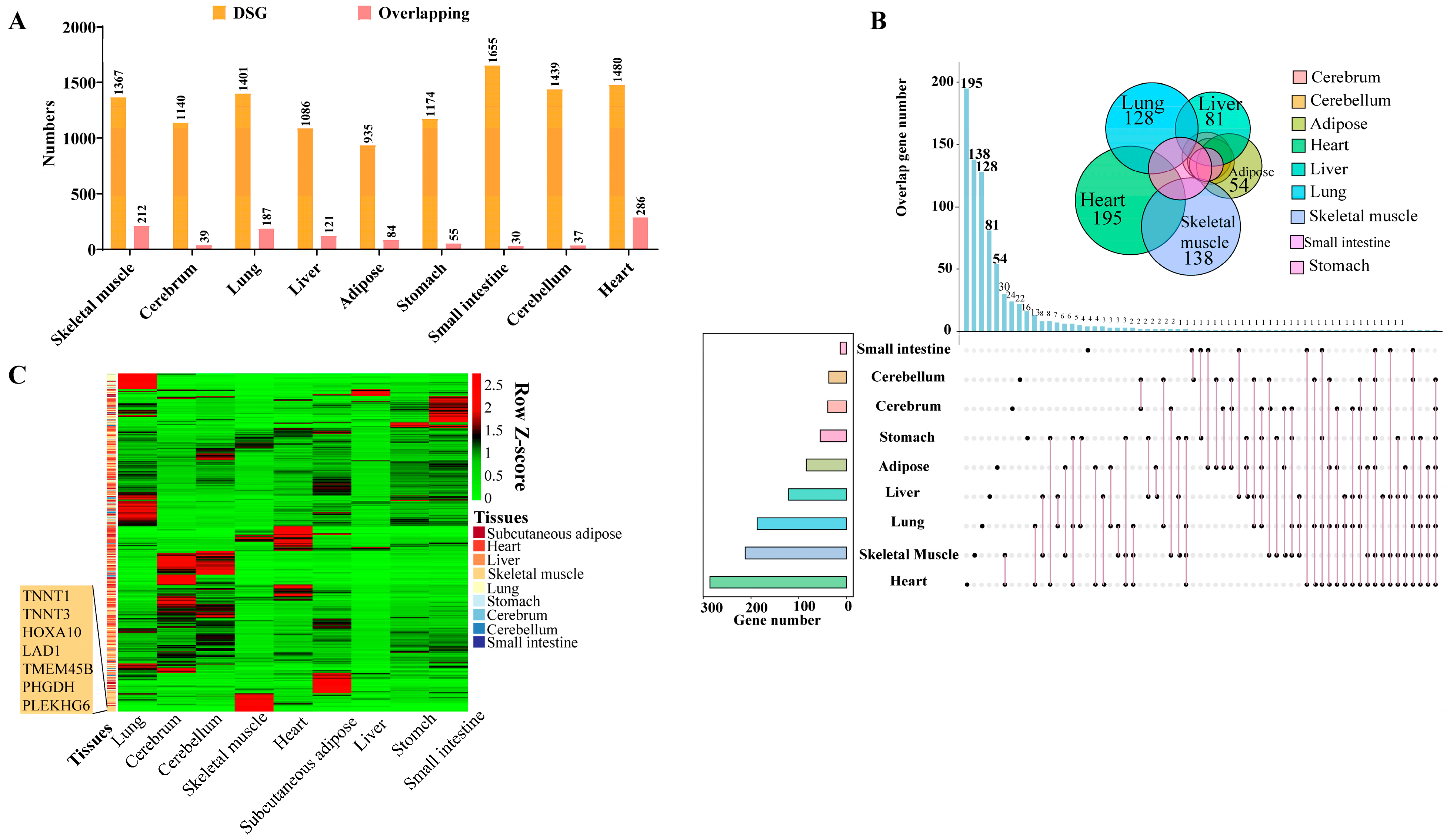
3.4. Comparison Analysis of Alternative Splicing between Duroc and Luchuan Pigs
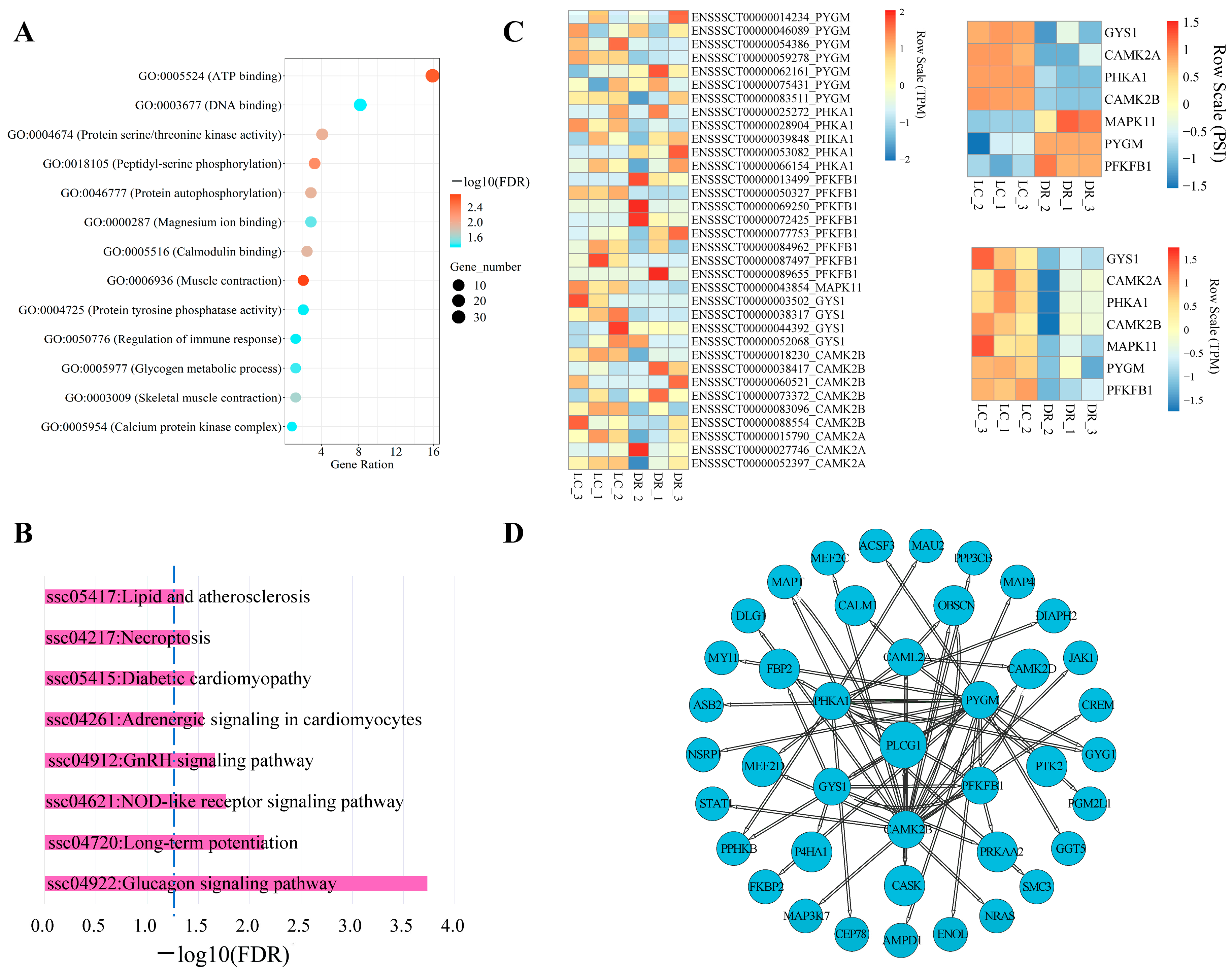
3.5. AS Analysis of Genes from Glucagon Signaling Pathway in Skeletal Muscle
3.6. Validation of AS Differences of Genes Related to the Glucagon Signaling Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Park, E.; Pan, Z.C.; Zhang, Z.J.; Lin, L.; Xing, Y. The Expanding Landscape of Alternative Splicing Variation in Human Populations. Am. J. Hum. Genet. 2018, 102, 11–26. [Google Scholar] [CrossRef]
- Marasco, L.E.; Kornblihtt, A.R. The physiology of alternative splicing. Nat. Rev. Mol. Cell Biol. 2023, 24, 242–254. [Google Scholar] [CrossRef]
- Zea, D.J.; Laskina, S.; Baudin, A.; Richard, H.; Laine, E. Assessing conservation of alternative splicing with evolutionary splicing graphs. Genome Res. 2021, 31, 1462–1473. [Google Scholar] [CrossRef]
- Baralle, F.E.; Giudice, J. Alternative splicing as a regulator of development and tissue identity. Nat. Rev. Mol. Cell Biol. 2017, 18, 437–451. [Google Scholar] [CrossRef]
- Pan, Q.; Shai, O.; Lee, L.J.; Frey, B.J.; Blencowe, B.J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 2008, 40, 1413–1415. [Google Scholar] [CrossRef]
- Cieply, B.; Carstens, R.P. Functional roles of alternative splicing factors in human disease. Wiley Interdiscip. Rev. RNA 2015, 6, 311–326. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.Y.; Alipanahi, B.; Lee, L.J.; Bretschneider, H.; Merico, D.; Yuen, R.K.C.; Hua, Y.; Gueroussov, S.; Najafabadi, H.S.; Hughes, T.R.; et al. The human splicing code reveals new insights into the genetic determinants of disease. Science 2015, 347, 1254806. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Si, J.; Yue, J.; Zhao, M.; Qi, W.; Zhu, S.; Mo, J.; Wang, L.; Lan, G.; Liang, J. The Landscape of Accessible Chromatin and Developmental Transcriptome Maps Reveal a Genetic Mechanism of Skeletal Muscle Development in Pigs. Int. J. Mol. Sci. 2023, 24, 6413. [Google Scholar] [CrossRef]
- Zhang, Y.; Xue, L.Y.; Xu, H.; Liang, W.; Wu, Q.Q.; Zhang, Q.D.; Zhou, X.; Liu, B. Global Analysis of Alternative Splicing Difference in Peripheral Immune Organs between Tongcheng Pigs and Large White Pigs Artificially Infected with PRRSV. Biomed. Res. Int. 2020, 2020, 4045204. [Google Scholar] [CrossRef] [PubMed]
- Giuffra, E.; Kijas, J.M.H.; Amarger, V.; Carlborg, Ö.; Jeon, J.T.; Andersson, L. The origin of the domestic pig: Independent domestication and subsequent introgression. Genetics 2000, 154, 1785–1791. [Google Scholar] [CrossRef]
- Groenen, M.A.M.; Archibald, A.L.; Uenishi, H.; Tuggle, C.K.; Takeuchi, Y.; Rothschild, M.F.; Rogel-Gaillard, C.; Park, C.; Milan, D.; Megens, H.J.; et al. Analyses of pig genomes provide insight into porcine demography and evolution. Nature 2012, 491, 393–398. [Google Scholar] [CrossRef]
- Koopmans, S.J.; Mroz, Z.; Dekker, R.; Corbijn, H.; Ackermans, M.; Sauerwein, H. Association of insulin resistance with hyperglycemia in streptozotocin-diabetic pigs: Effects of metformin at isoenergetic feeding in a type 2-like diabetic pig model. Metabolism 2006, 55, 960–971. [Google Scholar] [CrossRef] [PubMed]
- Lunney, J.K.; Van Goor, A.; Walker, K.E.; Hailstock, T.; Franklin, J.; Dai, C.H. Importance of the pig as a human biomedical model. Sci. Transl. Med. 2021, 13, eabd5758. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.G.; Wang, L.J.; Xu, Z.R.; Huang, J.F.; Wang, Y.R. Developmental changes of carcass composition, meat quality and organs in the Jinhua pig and Landrace. Animal 2009, 3, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.H.; Rodrigues, G.D.; Valente, D.T.; da Silva, C.B.; Costa, T.C.; Duarte, M.D.; Saraiva, A. Performance, Carcass Traits, Pork Quality and Expression of Genes Related to Intramuscular Fat Metabolism of Two Diverse Genetic Lines of Pigs. Foods 2022, 11, 2280. [Google Scholar] [CrossRef] [PubMed]
- Miao, W.W.; Ma, Z.Q.; Tang, Z.Y.; Yu, L.; Liu, S.Q.; Huang, T.D.; Wang, P.; Wu, T.; Song, Z.Y.; Zhang, H.J.; et al. Integrative ATAC-seq and RNA-seq Analysis of the Longissimus Muscle of Luchuan and Duroc Pigs. Front. Nutr. 2021, 8, 742672. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.Y.; Jung, S.; Kook, T.L.; Rouchka, E.C.; Bok, J.; Park, J.W. rMAPS2: An update of the RNA map analysis and plotting server for alternative splicing regulation. Nucleic Acids Res. 2020, 48, W300–W306. [Google Scholar] [CrossRef]
- Chen, M.Y.; Yao, Y.L.; Yang, Y.L.; Zhu, M.; Tang, Y.J.; Liu, S.Y.; Li, K.; Tang, Z.L. Comprehensive Profiles of mRNAs and miRNAs Reveal Molecular Characteristics of Multiple Organ Physiologies and Development in Pigs. Front. Genet. 2019, 10, 756. [Google Scholar] [CrossRef]
- Nygård, A.-B.; Cirera, S.; Gilchrist, M.J.; Gorodkin, J.; Jørgensen, C.B.; Fredholm, M. A study of alternative splicing in the pig. BMC Res. Notes 2010, 3, 123. [Google Scholar] [CrossRef]
- Hao, W.; Yang, Z.; Sun, Y.; Li, J.; Zhang, D.; Liu, D.; Yang, X. Characterization of Alternative Splicing Events in Porcine Skeletal Muscles with Different Intramuscular Fat Contents. Biomolecules 2022, 12, 154. [Google Scholar] [CrossRef]
- Liu, Y.W.; Fu, Y.; Yang, Y.L.; Yi, G.Q.; Lian, J.M.; Xie, B.K.; Yao, Y.L.; Chen, M.Y.; Niu, Y.C.; Liu, L.; et al. Integration of multi-omics data reveals cis-regulatory variants that are associated with phenotypic differentiation of eastern from western pigs. Genet. Sel. Evol. 2022, 54, 62. [Google Scholar] [CrossRef]
- Chen, S.F.; Zhou, Y.Q.; Chen, Y.R.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Landmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Melé, M.; Ferreira, P.G.; Reverter, F.; DeLuca, D.S.; Monlong, J.; Sammeth, M.; Young, T.R.; Goldmann, J.M.; Pervouchine, D.D.; Sullivan, T.J.; et al. The human transcriptome across tissues and individuals. Science 2015, 348, 660–665. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Tan, Q.; Kir, J.; Liu, D.; Bryant, D.; Guo, Y.; Stephens, R.; Baseler, M.W.; Lane, H.C.; et al. DAVID Bioinformatics Resources: Expanded annotation database and novel algorithms to better extract biology from large gene lists. Nucleic Acids Res. 2007, 35, W169–W175. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Feng, S.L.; Li, J.M.; Wen, H.; Liu, K.; Gui, Y.Q.; Wen, Y.J.; Wang, X.L.; Yuan, S.Q. hnRNPH1 recruits PTBP2 and SRSF3 to modulate alternative splicing in germ cells. Nat. Commun. 2022, 13, 3588. [Google Scholar] [CrossRef]
- Li, Q.Q.; Zhang, Y.P.; Li, W.C.; Yan, K.; Liu, Y.R.; Xu, H.Y.; Lu, Y.Q.; Liang, X.W.; Yang, X.G. Allicin protects porcine oocytes against LPS-induced defects during maturation in vitro. Theriogenology 2022, 182, 138–147. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef]
- Feng, W.; Zhao, P.; Zheng, X.; Hu, Z.; Liu, J. Profiling Novel Alternative Splicing within Multiple Tissues Provides Useful Insights into Porcine Genome Annotation. Genes 2020, 11, 1405. [Google Scholar] [CrossRef]
- Shen, S.H.; Park, J.W.; Lu, Z.X.; Lin, L.; Henry, M.D.; Wu, Y.N.; Zhou, Q.; Xing, Y. rMATS: Robust and flexible detection of differential alternative splicing from replicate RNA-Seq data. Proc. Natl. Acad. Sci. USA 2014, 111, E5593–E5601. [Google Scholar] [CrossRef]
- Eghbali, A.; Dukes, A.; Toischer, K.; Hasenfuss, G.; Field, L.J. Cell Cycle-Mediated Cardiac Regeneration in the Mouse Heart. Curr. Cardiol. Rep. 2019, 21, 131. [Google Scholar] [CrossRef]
- Mastej, E.J.; DeBoer, E.M.; Humphries, S.M.; Cook, M.C.; Hunter, K.S.; Liptzin, D.R.; Weinman, J.P.; Deterding, R.R. Lung and airway shape in neuroendocrine cell hyperplasia of infancy. Pediatr. Radiol. 2018, 48, 1745–1754. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Chen, N.; Chen, Y.H.; Sun, L.; Li, L.; Liu, H. Elevated GPC3 level promotes cell proliferation in liver cancer. Oncol. Lett. 2018, 16, 970–976. [Google Scholar] [CrossRef] [PubMed]
- Pant, M.; Bal, N.C.; Periasamy, M. Sarcolipin: A Key Thermogenic and Metabolic Regulator in Skeletal Muscle. Trends Endocrinol. Metab. 2016, 27, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Zha, C.W.; Jiang, A.W.; Chao, Z.; Hou, L.M.; Liu, H.L.; Huang, R.H.; Wu, W.J. A Combined Differential Proteome and Transcriptome Profiling of Fast- and Slow-Twitch Skeletal Muscle in Pigs. Foods 2022, 11, 2842. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yan, J.; Fan, X.; Chen, J.; Wang, Z.; Liu, X.; Yi, G.; Liu, Y.; Niu, Y.; Zhang, L.; et al. The genome variation and developmental transcriptome maps reveal genetic differentiation of skeletal muscle in pigs. PLoS Genet. 2021, 17, e1009910. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Fan, X.; Yan, J.; Chen, M.; Zhu, M.; Tang, Y.; Liu, S.; Tang, Z. A comprehensive epigenome atlas reveals DNA methylation regulating skeletal muscle development. Nucleic Acids Res. 2021, 49, 1313–1329. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.L.; Zhou, R.; Mu, Y.L.; Hou, X.H.; Tang, Z.L.; Li, K. Genome-wide analysis of DNA methylation in obese, lean, and miniature pig breeds. Sci. Rep. 2016, 6, 30160. [Google Scholar] [CrossRef]
- Braunschweig, U.; Gueroussov, S.; Plocik, A.M.; Graveley, B.R.; Blencowe, B.J. Dynamic Integration of Splicing within Gene Regulatory Pathways. Cell 2013, 152, 1252–1269. [Google Scholar] [CrossRef]
- Tang, L.T.; Ran, X.Q.; Mao, N.; Zhang, F.P.; Niu, X.; Ruan, Y.Q.; Yi, F.L.; Li, S.; Wang, J.F. Analysis of alternative splicing events by RNA sequencing in the ovaries of Xiang pig at estrous and diestrous. Theriogenology 2018, 119, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Sammeth, M.; Foissac, S.; Guigó, R. A General Definition and Nomenclature for Alternative Splicing Events. PLoS Comput. Biol. 2008, 4, e1000147. [Google Scholar] [CrossRef]
- Han, J.; An, O.; Ren, X.; Song, Y.; Tang, S.J.; Shen, H.; Ke, X.; Ng, V.H.E.; Tay, D.J.T.; Tan, H.Q.; et al. Multilayered control of splicing regulatory networks by DAP3 leads to widespread alternative splicing changes in cancer. Nat. Commun. 2022, 13, 1793. [Google Scholar] [CrossRef] [PubMed]
- Petrillo, E. Do not panic: An intron-centric guide to alternative splicing. Plant Cell 2023, 35, 1752–1761. [Google Scholar] [CrossRef] [PubMed]
- Leung, S.K.; Jeffries, A.R.; Castanho, I.; Jordan, B.T.; Moore, K.; Davies, J.P.; Dempster, E.L.; Bray, N.J.; O’Neill, P.; Tseng, E.; et al. Full-length transcript sequencing of human and mouse cerebral cortex identifies widespread isoform diversity and alternative splicing. Cell Rep. 2021, 37, 110022. [Google Scholar] [CrossRef]
- Brogna, S.; McLeod, T.; Petric, M. The Meaning of NMD: Translate or Perish. Trends Genet. 2016, 32, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Supek, F.; Lehner, B.; Lindeboom, R.G.H. To NMD or Not To NMD: Nonsense-Mediated mRNA Decay in Cancer and Other Genetic Diseases. Trends Genet. 2021, 37, 657–668. [Google Scholar] [CrossRef]
- Yang, J.; Hung, L.H.; Licht, T.; Kostin, S.; Looso, M.; Khrameeva, E.; Bindereif, A.; Schneider, A.; Braun, T. RBM24 is a major regulator of muscle-specific alternative splicing. Dev. Cell 2014, 31, 87–99. [Google Scholar] [CrossRef]
- Singh, R.K.; Xia, Z.; Bland, C.S.; Kalsotra, A.; Scavuzzo, M.A.; Curk, T.; Ule, J.; Li, W.; Cooper, T.A. Rbfox2-coordinated alternative splicing of Mef2d and Rock2 controls myoblast fusion during myogenesis. Mol. Cell 2014, 55, 592–603. [Google Scholar] [CrossRef]
- Montanes-Agudo, P.; Aufiero, S.; Schepers, E.N.; van der Made, I.; Cocera-Ortega, L.; Ernault, A.C.; Richard, S.; Kuster, D.W.D.; Christoffels, V.M.; Pinto, Y.M.; et al. The RNA-binding protein QKI governs a muscle-specific alternative splicing program that shapes the contractile function of cardiomyocytes. Cardiovasc. Res. 2023, 119, 1161–1174. [Google Scholar] [CrossRef]
- Shu, Z.; Wang, L.G.; Wang, J.B.; Zhang, L.C.; Hou, X.H.; Yan, H.; Wang, L.X. Integrative Analysis of Nanopore and Illumina Sequencing Reveals Alternative Splicing Complexity in Pig Longissimus Dorsi Muscle. Front. Genet. 2022, 13, 877646. [Google Scholar] [CrossRef]
- Black, A.J.; Schilder, R.J.; Kimball, S.R. Palmitate- and C6 ceramide-induced pre-mRNA alternative splicing occurs in a PP2A dependent manner. Nutr. Metab. 2018, 15, 87. [Google Scholar] [CrossRef]
- Chen, H.L.; Zhang, J.H.; Yu, B.; Li, L.; Shang, Y.S. Molecular cloning, structural analysis, and tissue expression of the TNNT3 gene in Guizhou black goat. Gene 2015, 573, 123–128. [Google Scholar] [CrossRef]
- Géraud, J.; Dieterich, K.; Rendu, J.; Coste, E.U.; Dobrzynski, M.; Marcorelle, P.; Ioos, C.; Romero, N.B.; Baudou, E.; Brocard, J.; et al. Clinical phenotype and loss of the slow skeletal muscle troponin T in three new patients with recessive TNNT1 nemaline myopathy. J. Med. Genet. 2021, 58, 602–608. [Google Scholar] [CrossRef]
- Novelli, G.; Gennarelli, M.; Zelano, G.; Sangiuolo, F.; Lo Cicero, S.; Samson, F.; Dallapiccola, B. Polymerase chain reaction in the detection of mRNA transcripts from the slow skeletal troponin T (TNNT1) gene in myotonic dystrophy and normal muscle. Cell Biochem. Funct. 1992, 10, 251–256. [Google Scholar] [CrossRef]
- Ravi, S.; Schilder, R.J.; Berg, A.S.; Kimball, S.R. Effects of age and hindlimb immobilization and remobilization on fast troponin T precursor mRNA alternative splicing in rat gastrocnemius muscle. Appl. Physiol. Nutr. Metab. 2016, 41, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wu, Y.L.; Ding, H.; Xie, K.Z.; Zhang, T.; Zhang, G.X.; Wang, J.Y. promotes the proliferation and differentiation of primary chicken myoblasts. Br. Poult. Sci. 2022, 63, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Altea-Manzano, P.; Demicco, M.; Doglioni, G.; Bornes, L.; Fukano, M.; Vandekeere, A.; Cuadros, A.M.; Fernández-García, J.; Riera-Domingo, C.; et al. PHGDH heterogeneity potentiates cancer cell dissemination and metastasis. Nature 2022, 605, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.P.; Zhou, X.; Gong, Y.Q.; Gou, K.; Luo, Y.F.; Jia, D.; Dai, L.Z.; Zhao, Y.L.; Sun, Q.X. Biophysical and biochemical properties of PHGDH revealed by studies on PHGDH inhibitors. Cell Mol. Life Sci. 2022, 79, 27. [Google Scholar] [CrossRef] [PubMed]
- Braun, T.; Gautel, M. Transcriptional mechanisms regulating skeletal muscle differentiation, growth and homeostasis. Nat. Rev. Mol. Cell Biol. 2011, 12, 349–361. [Google Scholar] [CrossRef]
- Bentzinger, C.F.; Wang, Y.X.; Rudnicki, M.A. Building muscle: Molecular regulation of myogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008342. [Google Scholar] [CrossRef] [PubMed]
- McNamara, E.L.; Taylor, R.L.; Clayton, J.S.; Goullee, H.; Dilworth, K.L.; Pinós, T.; Brull, A.; Alexander, I.E.; Lisowski, L.; Ravenscroft, G.; et al. Systemic A AV8-mediated delivery of a functional copy of muscle glycogen phosphorylase ameliorates disease in a murine model of McArdle disease. Hum. Mol. Genet. 2020, 29, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Lacham-Kaplan, O.; Camera, D.M.; Hawley, J.A. Divergent Regulation of Myotube Formation and Gene Expression by E2 and EPA during In-Vitro Differentiation of C2C12 Myoblasts. Int. J. Mol. Sci. 2020, 21, 745. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wu, M.L.; Liu, X.Q.; Wang, F.W.; Li, M.; An, X.Y.; Bai, F.X.; Lei, C.Z.; Dang, R.H. MiR-24-3p Conservatively Regulates Muscle Cell Proliferation and Apoptosis by Targeting Common Gene in Rat and Cattle. Animals 2022, 12, 505. [Google Scholar] [CrossRef] [PubMed]
- Park, C.Y.; Pierce, S.A.; von Drehle, M.; Ivey, K.N.; Morgan, J.; Blau, H.M.; Srivastava, D. skNAC, a Smyd1-interacting transcription factor, is involved in cardiac development and skeletal muscle growth and regeneration. Proc. Natl. Acad. Sci. USA 2010, 107, 20750–20755. [Google Scholar] [CrossRef]
- Sebastian, S.; Faralli, H.; Yao, Z.; Rakopoulos, P.; Palii, C.; Cao, Y.; Singh, K.; Liu, Q.C.; Chu, A.; Aziz, A.; et al. Tissue-specific splicing of a ubiquitously expressed transcription factor is essential for muscle differentiation. Genes Dev. 2013, 27, 1247–1259. [Google Scholar] [CrossRef]
- Blondelle, J.; Ohno, Y.; Gache, V.; Guyot, S.; Storck, S.; Blanchard-Gutton, N.; Barthélémy, I.; Walmsley, G.; Rahier, A.; Gadin, S.; et al. HACD1, a regulator of membrane composition and fluidity, promotes myoblast fusion and skeletal muscle growth. J. Mol. Cell Biol. 2015, 7, 429–440. [Google Scholar] [CrossRef]
- Gonczi, M.; Teixeira, J.M.C.; Barrera-Vilarmau, S.; Mediani, L.; Antoniani, F.; Nagy, T.M.; Feher, K.; Raduly, Z.; Ambrus, V.; Tozser, J.; et al. Alternatively spliced exon regulates context-dependent MEF2D higher-order assembly during myogenesis. Nat. Commun. 2023, 14, 1329. [Google Scholar] [CrossRef]
- Yuan, J.; Ma, Y.; Huang, T.; Chen, Y.; Peng, Y.; Li, B.; Li, J.; Zhang, Y.; Song, B.; Sun, X.; et al. Genetic Modulation of RNA Splicing with a CRISPR-Guided Cytidine Deaminase. Mol. Cell 2018, 72, 380–394. [Google Scholar] [CrossRef]
- Huang, S.; Liao, Z.; Li, X.; Liu, Z.; Li, G.; Li, J.; Lu, Z.; Zhang, Y.; Li, X.; Ma, X.; et al. Developing ABEmax-NG with Precise Targeting and Expanded Editing Scope to Model Pathogenic Splice Site Mutations In Vivo. iScience 2019, 15, 640–648. [Google Scholar] [CrossRef]
- Long, C.; Li, H.; Tiburcy, M.; Rodriguez-Caycedo, C.; Kyrychenko, V.; Zhou, H.; Zhang, Y.; Min, Y.-L.; Shelton, J.M.; Mammen, P.P.A.; et al. Correction of diverse muscular dystrophy mutations in human engineered heart muscle by single-site genome editing. Sci. Adv. 2018, 4, eaap9004. [Google Scholar] [CrossRef]
- Yang, S.-P.; Zhu, X.-X.; Qu, Z.-X.; Chen, C.-Y.; Wu, Y.-B.; Wu, Y.; Luo, Z.-D.; Wang, X.-Y.; He, C.-Y.; Fang, J.-W.; et al. Production of MSTN knockout porcine cells using adenine base-editing-mediated exon skipping. Vitr. Cell. Dev. Biol. Anim. 2023, 59, 241–255. [Google Scholar] [CrossRef]
- Bhadra, M.; Howell, P.; Dutta, S.; Heintz, C.; Mair, W.B. Alternative splicing in aging and longevity. Hum. Genet. 2019, 139, 357–369. [Google Scholar] [CrossRef]
- Li, D.; Yu, W.; Lai, M. Towards understandings of serine/arginine-rich splicing factors. Acta Pharm. Sin. B 2023, 13, 3181–3207. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, X.; Huang, L.; Huo, H.; Wang, P.; Li, W.; Dai, H.; Yang, F.; Fu, G.; Zhao, G.; et al. Long- and short-read RNA sequencing from five reproductive organs of boar. Sci. Data 2023, 10, 678. [Google Scholar] [CrossRef] [PubMed]
- Tien, J.F.; Mazloomian, A.; Cheng, S.G.; Hughes, C.S.; Chow, C.C.T.; Canapi, L.T.; Oloumi, A.; Trigo-Gonzalez, G.; Bashashati, A.; Xu, J.; et al. CDK12 regulates alternative last exon mRNA splicing and promotes breast cancer cell invasion. Nucleic Acids Res. 2017, 45, 6698–6716. [Google Scholar] [CrossRef] [PubMed]
- Nakka, K.; Ghigna, C.; Gabellini, D.; Dilworth, F.J. Diversification of the muscle proteome through alternative splicing. Skelet. Muscle 2018, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Cho, N.; Kim, K.K. The implications of alternative pre-mRNA splicing in cell signal transduction. Exp. Mol. Med. 2023, 55, 755–766. [Google Scholar] [CrossRef]
- Irimia, M.; Weatheritt, R.J.; Ellis, J.D.; Parikshak, N.N.; Gonatopoulos-Pournatzis, T.; Babor, M.; Quesnel-Vallières, M.; Tapial, J.; Raj, B.; O’Hanlon, D.; et al. A Highly Conserved Program of Neuronal Microexons Is Misregulated in Autistic Brains. Cell 2014, 159, 1511–1523. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Manley, J.L. Alternative polyadenylation of mRNA precursors. Nat. Rev. Mol. Cell Biol. 2016, 18, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, A.; Au, K.F. PacBio Sequencing and Its Applications. Genom. Proteom. Bioinform. 2015, 13, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-Y.; Deng, F.; Jia, X.; Li, C.; Lai, S.-J. A transcriptome atlas of rabbit revealed by PacBio single-molecule long-read sequencing. Sci. Rep. 2017, 7, 7648. [Google Scholar] [CrossRef] [PubMed]
- Mou, C.-Y.; Li, Q.; Huang, Z.-P.; Ke, H.-Y.; Zhao, H.; Zhao, Z.-M.; Duan, Y.-L.; Li, H.-D.; Xiao, Y.; Qian, Z.-M.; et al. PacBio single-molecule long-read sequencing provides new insights into the complexity of full-length transcripts in oriental river prawn, macrobrachium nipponense. BMC Genom. 2023, 24, 340. [Google Scholar] [CrossRef]
- Beiki, H.; Liu, H.; Huang, J.; Manchanda, N.; Nonneman, D.; Smith, T.P.L.; Reecy, J.M.; Tuggle, C.K. Improved annotation of the domestic pig genome through integration of Iso-Seq and RNA-seq data. BMC Genom. 2019, 20, 344. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Li, W.; Liu, W.; Wang, Z.; Xie, B.; Yang, X.; Tang, Z. Exploring Multi-Tissue Alternative Splicing and Skeletal Muscle Metabolism Regulation in Obese- and Lean-Type Pigs. Genes 2024, 15, 196. https://doi.org/10.3390/genes15020196
Wang W, Li W, Liu W, Wang Z, Xie B, Yang X, Tang Z. Exploring Multi-Tissue Alternative Splicing and Skeletal Muscle Metabolism Regulation in Obese- and Lean-Type Pigs. Genes. 2024; 15(2):196. https://doi.org/10.3390/genes15020196
Chicago/Turabian StyleWang, Wei, Wangchang Li, Weiwei Liu, Zishuai Wang, Bingkun Xie, Xiaogan Yang, and Zhonglin Tang. 2024. "Exploring Multi-Tissue Alternative Splicing and Skeletal Muscle Metabolism Regulation in Obese- and Lean-Type Pigs" Genes 15, no. 2: 196. https://doi.org/10.3390/genes15020196
APA StyleWang, W., Li, W., Liu, W., Wang, Z., Xie, B., Yang, X., & Tang, Z. (2024). Exploring Multi-Tissue Alternative Splicing and Skeletal Muscle Metabolism Regulation in Obese- and Lean-Type Pigs. Genes, 15(2), 196. https://doi.org/10.3390/genes15020196