
Genetic Landscape of Masticatory Muscle Tendon–Aponeurosis Hyperplasia

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
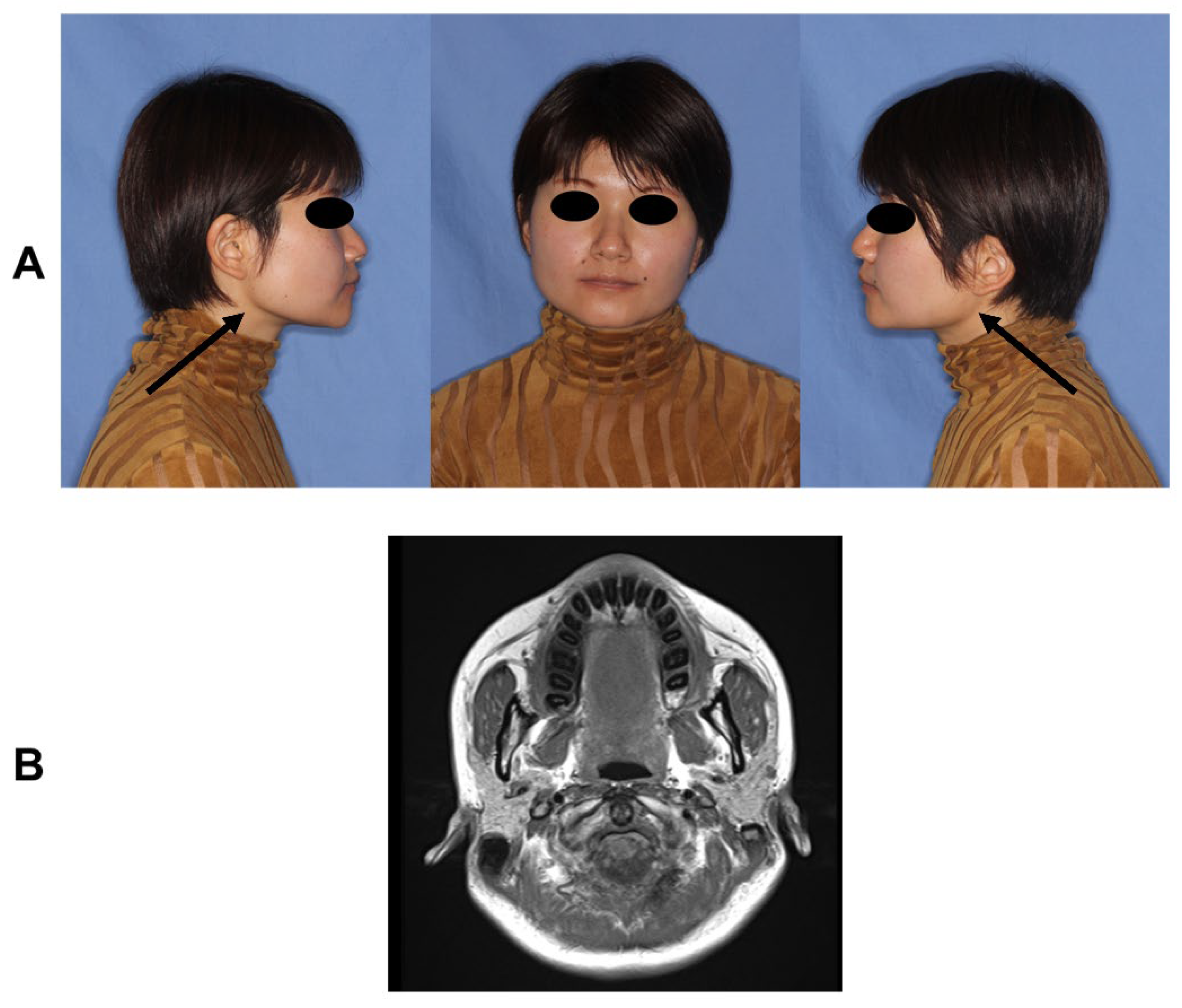
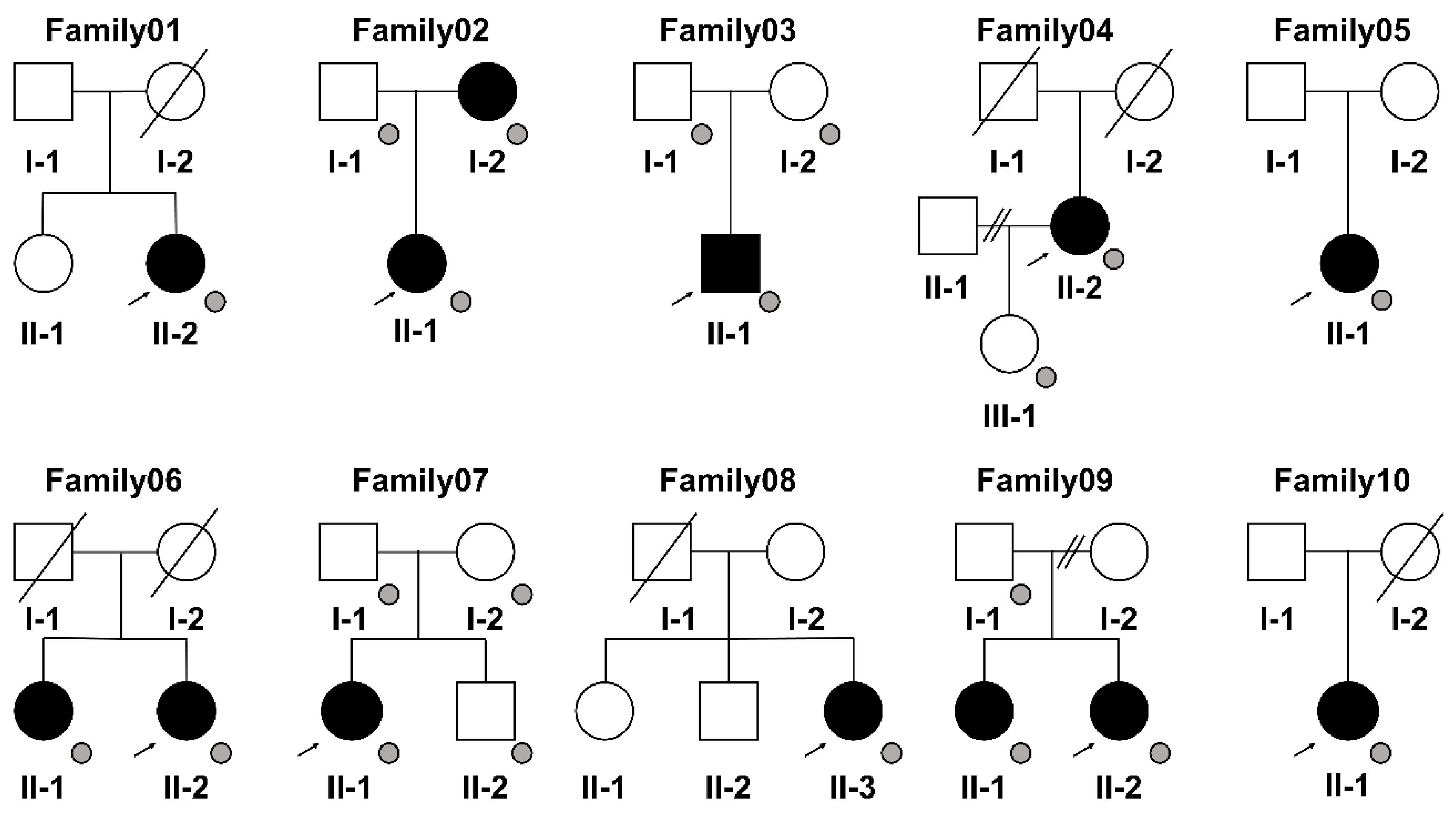
2.1. Patients
2.2. Pig Samples
2.3. Whole Genome Sequencing (WGS) and Variant Calling Pipelines
2.4. Variant Prioritization Pipeline
2.5. Transcriptome Sequencing (RNA-seq) of Pig Tissue Samples
3. Results
3.1. Variant Detection by WGS Analysis
3.2. Integration of the WGS Analysis of Patients with the RNA-seq Analysis of Pig Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schiffman, E.; Ohrbach, R.; Truelove, E.; Look, J.; Anderson, G.; Goulet, J.-P.; List, T.; Svensson, P.; Gonzalez, Y.; Lobbezoo, F.; et al. Diagnostic Criteria for Temporomandibular Disorders (DC/TMD) for Clinical and Research Applications: Recommendations of the International RDC/TMD Consortium Network and Orofacial Pain Special Interest Group. J. Oral Facial Pain Headache 2014, 28, 6–27. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.-H.; Lv, K.; Yang, R.-T.; Li, Z.; Yang, X.-W.; Li, Z.-B. Clinical, retrospective case-control study on the mechanics of obstacle in mouth opening and malocclusion in patients with maxillofacial fractures. Sci. Rep. 2018, 8, 7724. [Google Scholar] [CrossRef]
- Smith, M.J.; Myall, R.W. Tetanus: Review of the literature and report of a case. Oral Surg. Oral Med. Oral Pathol. 1976, 41, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Aliko, A.; Ciancaglini, R.; Alushi, A.; Tafaj, A.; Ruci, D. Temporomandibular joint involvement in rheumatoid arthritis, systemic lupus erythematosus and systemic sclerosis. Int. J. Oral Maxillofac. Surg. 2011, 40, 704–709. [Google Scholar] [CrossRef]
- Jager-Wittenaar, H.; Dijkstra, P.U.; Vissink, A.; van Oort, R.P.; Roodenburg, J.L. Variation in repeated mouth-opening measurements in head and neck cancer patients with and without trismus. Int. J. Oral Maxillofac. Surg. 2009, 38, 26–30. [Google Scholar] [CrossRef]
- Çorumlu, U.; Kopuz, C.; Demir, M.T.; Pirzirenli, M.E. Bilateral elongated mandibular coronoid process in an Anatolian skull. Anat. Cell Biol. 2016, 49, 217–220. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Khandelwal, P.; Dhupar, V.; Akkara, F.; Louis, A. Zygomatico-coronoid ankylosis as sequel of inadequate treatment. Ann. Maxillofac. Surg. 2018, 8, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Arika, T.; Kakudo, K. Hyperplasia of tendon and aponeurosis of masticatory muscles (HyTAM)—Clinical appearance. J. Jpn. Soc. Temporomandibular Jt. 2009, 21, 31–34. [Google Scholar]
- Kakudo, K.; Yoda, T. New concept of limited mouth opening associated with square mandible: Diagnosis and treatment for masticatory muscle tendon-aponeurosis hyperplasia. J. Jpn. Soc. Temporomandibular Jt. 2009, 21, 28–30. [Google Scholar]
- Hayashi, N.; Yoda, T.; Tomoda, T.; Yumoto, M.; Kitamura, T.; Okubo, M.; Iwasaki, Y.; Enoki, Y.; Sato, T. Longitudinal epidemiological survey on suspected masticatory muscle tendon-aponeurosis hyperplasia in adolescents. J. Jpn. Soc. Temporomandibular Jt. 2018, 30, 187–194. [Google Scholar] [CrossRef]
- Worni, A.; Mericske-Stern, R.; Iizuka, T.; Büttner, M. Limited mouth opening—What now? Swiss Dent. J. 2014, 124, 935–944. [Google Scholar] [PubMed]
- Yoda, T.; Sato, T.; Abe, T.; Sakamoto, I.; Tomaru, Y.; Omura, K.; Hatano, N.; Takato, T.; Ishii, Y. Long-term results of surgical therapy for masticatory muscle tendon-aponeurosis hyperplasia accompanied by limited mouth opening. Int. J. Oral Maxillofac. Surg. 2009, 38, 1143–1147. [Google Scholar] [CrossRef] [PubMed]
- Ajila, V.; Hegde, S.; Gopakumar, R.; Babu, G.S. Imaging and Histopathological Features of Jacob’s Disease: A Case Study. Head Neck Pathol. 2012, 6, 51–53. [Google Scholar] [CrossRef][Green Version]
- Kobayashi, K.; Shimoda, S.; Yoda, T.; Kakudo, K. Current status of MR imaging for the masticatory muscle tendon-aponeurosis hyperplasia. J. Jpn. Soc. Temporomandibular Jt. 2009, 21, 35–39. [Google Scholar]
- Hayashi, N.; Sato, T.; Fukushima, Y.; Takano, A.; Sakamoto, I.; Yoda, T. A two-year follow-up of surgical and non-surgical treatments in patients with masticatory muscle tendon-aponeurosis hyperplasia. Int. J. Oral Maxillofac. Surg. 2018, 47, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Hori, N.; Nakamoto, N.; Akita, M.; Yoda, T. Masticatory muscle tendon-aponeurosis hyperplasia exhibits heterotopic calcification in tendons. Oral Dis. 2014, 20, 404–408. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, A.; Sato, T.; Hirosawa, N.; Nakamoto, N.; Enoki, Y.; Chida, D.; Usui, M.; Takeda, S.; Nagai, T.; Sasaki, A.; et al. Proteomics-based identification of novel proteins in temporal tendons of patients with masticatory muscle tendon–aponeurosis hyperplasia. Int. J. Oral Maxillofac. Surg. 2014, 43, 113–119. [Google Scholar] [CrossRef]
- Hayashi, N.; Sato, T.; Kokabu, S.; Usui, M.; Yumoto, M.; Ikami, E.; Sakamoto, Y.; Nifuji, A.; Hayata, T.; Noda, M.; et al. Possible association of oestrogen and Cryba4 with masticatory muscle tendon-aponeurosis hyperplasia. Oral Dis. 2019, 25, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, N.; Sato, T.; Yumoto, M.; Kokabu, S.; Fukushima, Y.; Kawata, Y.; Kajihara, T.; Mizuno, Y.; Mizuno, Y.; Kawakami, T.; et al. Cyclic stretch induces decorin expression via yes-associated protein in tenocytes: A possible mechanism for hyperplasia in masticatory muscle tendon-aponeurosis hyperplasia. J. Oral Maxillofac. Surg. Med. Pathol. 2019, 31, 175–179. [Google Scholar] [CrossRef]
- Ito, K.; Go, Y.; Tatsumoto, S.; Usui, C.; Mizuno, Y.; Ikami, E.; Isozaki, Y.; Usui, M.; Kajihara, T.; Yoda, T.; et al. Gene expression profiling of the masticatory muscle tendons and Achilles tendons under tensile strain in the Japanese macaque Macaca fuscata. PLoS ONE 2023, 18, e0280649. [Google Scholar] [CrossRef] [PubMed]
- Yumoto, M.; Mizuno, Y.; Isozaki, Y.; Ito, K.O.; Yoda, T.; Sato, T. Analysis of Masticatory Muscle Tendon-aponeurosis Hyperplasia by Using Next-generation Sequencing. In Vivo 2022, 36, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Austin-Tse, C.A.; Jobanputra, V.; Perry, D.L.; Bick, D.; Taft, R.J.; Venner, E.; Gibbs, R.A.; Young, T.; Barnett, S.; Belmont, J.W.; et al. Best practices for the interpretation and reporting of clinical whole genome sequencing. NPJ Genom. Med. 2022, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Marshall, C.R.; Chowdhury, S.; Taft, R.J.; Lebo, M.S.; Buchan, J.G.; Harrison, S.M.; Rowsey, R.; Klee, E.W.; Liu, P.; Worthey, E.A.; et al. Best practices for the analytical validation of clinical whole-genome sequencing intended for the diagnosis of germline disease. NPJ Genom. Med. 2020, 5, 47. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows—Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. GigaScience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Van der Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From FastQ Data to high-confidence variant calls: The Genome Analysis Toolkit best practices pipeline. Curr. Protoc. Bioinform. 2013, 43, 11.10.11–11.10.33. [Google Scholar] [CrossRef]
- Wang, K.; Li, M.; Hakonarson, H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar] [CrossRef]
- Kishita, Y.; Shimura, M.; Kohda, M.; Akita, M.; Imai-Okazaki, A.; Yatsuka, Y.; Nakajima, Y.; Ito, T.; Ohtake, A.; Murayama, K.; et al. A novel homozygous variant in MICOS13/QIL1 causes hepato-encephalopathy with mitochondrial DNA depletion syndrome. Mol. Genet. Genom. Med. 2020, 8, e1427. [Google Scholar] [CrossRef] [PubMed]
- Sherry, S.T.; Ward, M.-H.; Kholodov, M.; Baker, J.; Phan, L.; Smigielski, E.M.; Sirotkin, K. dbSNP: The NCBI database of genetic variation. Nucleic Acids Res. 2001, 29, 308–311. [Google Scholar] [CrossRef]
- Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; Abecasis, G.R.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef]
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alfoldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef]
- Tadaka, S.; Hishinuma, E.; Komaki, S.; Motoike, I.N.; Kawashima, J.; Saigusa, D.; Inoue, J.; Takayama, J.; Okamura, Y.; Aoki, Y.; et al. jMorp updates in 2020: Large enhancement of multi-omics data resources on the general Japanese population. Nucleic Acids Res. 2021, 49, D536–D544. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. feature Counts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Caruso, N.; Herberth, B.; Bartoli, M.; Puppo, F.; Dumonceaux, J.; Zimmermann, A.; Denadai, S.; Lebossé, M.; Roche, S.; Geng, L.; et al. Deregulation of the protocadherin gene FAT1 alters muscle shapes: Implications for the pathogenesis of facioscapulohumeral dystrophy. PLoS Genet. 2013, 9, e1003550. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.G.; Van Hateren, N.; Tickle, C.; Wilson, S.A. The expression of Fat-1 cadherin during chick limb development. Int. J. Dev. Biol. 2007, 51, 173–176. [Google Scholar] [CrossRef]
- Tissir, F.; Qu, Y.; Montcouquiol, M.; Zhou, L.; Komatsu, K.; Shi, D.; Fujimori, T.; Labeau, J.; Tyteca, D.; Courtoy, P.; et al. Lack of cadherins Celsr2 and Celsr3 impairs ependymal ciliogenesis, leading to fatal hydrocephalus. Nat. Neurosci. 2010, 13, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Seeger-Nukpezah, T.; Golemis, E.A. The extracellular matrix and ciliary signaling. Curr. Opin. Cell Biol. 2012, 24, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Sbardella, D.; Tundo, G.R.; Fasciglione, G.F.; Gioia, M.; Bisicchia, S.; Gasbarra, E.; Ippolito, E.; Tarantino, U.; Coletta, M.; Marini, S. Role of metalloproteinases in tendon pathophysiology. Mini-Rev. Med. Chem. 2014, 14, 978–987. [Google Scholar] [CrossRef]
- Shi, Y.; Hu, G.; Su, J.; Li, W.; Chen, Q.; Shou, P.; Xu, C.; Chen, X.; Huang, Y.; Zhu, Z.; et al. Mesenchymal stem cells: A new strategy for immunosuppression and tissue repair. Cell Res. 2010, 20, 510–518. [Google Scholar] [CrossRef]
- Sadeghi Shaker, M.; Rokni, M.; Mahmoudi, M.; Farhadi, E. Ras family signaling pathway in immunopathogenesis of inflammatory rheumatic diseases. Front. Immunol. 2023, 14, 1151246. [Google Scholar] [CrossRef]
- Ayres, J.A.; Shum, L.; Akarsu, A.; Dashner, R.; Takahashi, K.; Ikura, T.; Slavkin, H.C.; Nuckolls, G.H. DACH: Genomic Characterization, evaluation as a candidate for postaxial polydactyly type A2, and developmental expression pattern of the mouse homologue. Genomics 2001, 77, 18–26. [Google Scholar] [CrossRef]
- Li, X.; Ohgi, K.A.; Rosenfeld, M.G.; Zhang, J.; Krones, A.; Bush, K.T.; Glass, C.K.; Nigam, S.K.; Aggarwal, A.K.; Maas, R.L.; et al. Eya protein phosphatase activity regulates Six1–Dach–Eya transcriptional effects in mammalian organogenesis. Nature 2003, 426, 247–254. [Google Scholar] [CrossRef]
- Maire, P.; Dos Santos, M.; Madani, R.; Sakakibara, I.; Viaut, C.; Wurmser, M. Myogenesis control by SIX transcriptional complexes. Semin. Cell Dev. Biol. 2020, 104, 51–64. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Patient (Family) | Sex | Age at the Time of Sample Collection (Years) | Maximum Mouth Opening (mm) |
|---|---|---|---|
| Patient01 (II-2 of Family01) | Female | 48 | 28 |
| Patient02 (II-1 of Family02) | Female | 37 | 25 |
| Patient03 (II-1 of Family03) | Male | 43 | 22 |
| Patient04 (II-2 of Family04) | Female | 79 | 22 |
| Patient05 (II-1 of Family05) | Female | 39 | 22 |
| Patient06 (II-2 of Family06) | Female | 60 | 14 |
| Patient07 (II-1 of Family07) | Female | 44 | 23 |
| Patient08 (II-3 of Family08) | Female | 46 | 20 |
| Patient09 (II-2 of Family09) | Female | 47 | 25 |
| Patient10 (II-1 of Family10) | Female | 44 | 23 |
| ID/Gene | Family01 | Family02 | Family03 | Family04 | Family05 | Family06 | Family07 | Family08 | Family09 | Family10 |
|---|---|---|---|---|---|---|---|---|---|---|
| ALPL | DYSF | BCO2 | ARHGAP27 | CAT | DHTKD1 | FUT1 | SOX18 | BAIAP3 | ||
| DACH1 | ESAM | CELSR2 | CD300LG | CCDC68 | EDNRB | HIP1R | SURF2 | CASKIN1 | ||
| EFCAB5 | NCR1 | ENPEP | CTNNBIP1 | GIPR | NEURL1B | MCAT | CYP27A1 | |||
| MMP15 | RASGRP3 | EPN3 | DHTKD1 | NBEA | STOML3 | TDRD10 | LLGL2 | |||
| NECTIN2 | FHIP1B | DYSF | ADAP2 | SULT2B1 | ||||||
| NT5DC3 | ETFB | |||||||||
| PCDH1 | FBLIM1 | |||||||||
| RASIP1 | GPR4 | |||||||||
| TBC1D30 | HPDL | |||||||||
| TMEM140 | PCDH1 | |||||||||
| TMEM266 | SCARB1 | |||||||||
| TRAF3IP3 | ST6GALNAC2 | |||||||||
| BAIAP3 | WIPF3 |
| Gene Symbols | Descriptions | Family_1 | Family_2 |
|---|---|---|---|
| DACH1 | DACHSHUND FAMILY TRANSCRIPTION FACTOR 1; DACH1 | Family01 | |
| EFCAB5 | DIHYDROPYRIMIDINASE-LIKE 2; DPYSL2 | Family01 | |
| MMP15 | MATRIX METALLOPROTEINASE 15; MMP15 | Family01 | |
| NECTIN2 | NECTIN CELL ADHESION MOLECULE 2; NECTIN2 | Family01 | |
| NCR1 | NATURAL CYTOTOXICITY TRIGGERING RECEPTOR 1; NCR1 | Family02 * | |
| RASGRP3 | RAS GUANYL NUCLEOTIDE-RELEASING PROTEIN 3; RASGRP3 | Family02 * | |
| BCO2 | BETA-CAROTENE OXYGENASE 2; BCO2 | Family04 * | |
| CELSR2 | CADHERIN EGF LAG SEVEN-PASS G-TYPE RECEPTOR 2; CELSR2 | Family04 * | |
| ENPEP | GLUTAMYL AMINOPEPTIDASE; ENPEP | Family04 * | |
| EPN3 | EPSIN 3; EPN3 | Family04 * | |
| FHIP1B | FHF COMPLEX SUBUNIT HOOK-INTERACTING PROTEIN 1B; FHIP1B | Family04 * | |
| NT5DC3 | 5-PRIME-NUCLEOTIDASE DOMAIN-CONTAINING PROTEIN 3; NT5DC3 | Family04 * | |
| PCDH1 | PROTOCADHERIN 1; PCDH1 | Family04 * | Family05 |
| RASIP1 | RAS-INTERACTING PROTEIN 1; RASIP1 | Family04 * | |
| TBC1D30 | TBC1 DOMAIN FAMILY, MEMBER 30; TBC1D30 | Family04 * | |
| TMEM140 | TRANSMEMBRANE PROTEIN 140; TMEM140 | Family04 * | |
| TMEM266 | TRANSMEMBRANE PROTEIN 266; TMEM266 | Family04 * | |
| TRAF3IP3 | TRAF3-INTERACTING PROTEIN 3; TRAF3IP3 | Family04 * | |
| ARHGAP27 | RHO GTPase-ACTIVATING PROTEIN 27; ARHGAP27 | Family05 | |
| CD300LG | CD300 ANTIGEN-LIKE FAMILY, MEMBER G; CD300LG | Family05 | |
| CTNNBIP1 | CATENIN, BETA-INTERACTING PROTEIN 1; CTNNBIP1 | Family05 | |
| FBLIM1 | FILAMIN-BINDING LIM PROTEIN 1; FBLIM1 | Family05 | |
| GPR4 | G PROTEIN-COUPLED RECEPTOR 4; GPR4 | Family05 | |
| ST6GALNAC2 | ST6 ALPHA-N-ACETYL-NEURAMINYL-2,3-BETA-GALACTOSYL-1,3-N-ACETYLGALACTOSAMINIDE ALPHA-2,6-SIALYLTRANSFERASE 2; ST6GALNAC2 | Family05 | |
| WIPF3 | WAS/WASL-INTERACTING PROTEIN FAMILY, MEMBER 3; WIPF3 | Family05 | |
| CCDC68 | COILED-COIL DOMAIN-CONTAINING PROTEIN 68; CCDC68 | Family06 * | |
| GIPR | GASTRIC INHIBITORY POLYPEPTIDE RECEPTOR; GIPR | Family06 * | |
| NEURL1B | NEURALIZED E3 UBIQUITIN PROTEIN LIGASE 1B; NEURL1B | Family07 * | |
| STOML3 | STOMATIN-LIKE PROTEIN 3; STOML3 | Family07 * | |
| HIP1R | HUNTINGTIN-INTERACTING PROTEIN 1-RELATED PROTEIN; HIP1R | Family08 | |
| MCAT | MALONYL CoA:ACP ACYLTRANSFERASE, MITOCHONDRIAL; MCAT | Family08 | |
| TDRD10 | TUDOR DOMAIN CONTAINING 10 | Family08 | |
| SURF2 | SURFEIT 2; SURF2 | Family09 * | |
| BAIAP3 | BAI1-ASSOCIATED PROTEIN 3; BAIAP3 | Family04 * | Family10 |
| CASKIN1 | CASK-INTERACTING PROTEIN 1; CASKIN1 | Family10 | |
| LLGL2 | LLGL SCRIBBLE CELL POLARITY COMPLEX COMPONENT 2; LLGL2 | Family10 | |
| ADAP2 | ARFGAP WITH DUAL PLECKSTRIN HOMOLOGY DOMAINS 2; ADAP2 | Family8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tajima, R.; Okazaki, A.; Sato, T.; Ozaki, K.; Motooka, D.; Okazaki, Y.; Yoda, T. Genetic Landscape of Masticatory Muscle Tendon–Aponeurosis Hyperplasia. Genes 2023, 14, 1718. https://doi.org/10.3390/genes14091718
Tajima R, Okazaki A, Sato T, Ozaki K, Motooka D, Okazaki Y, Yoda T. Genetic Landscape of Masticatory Muscle Tendon–Aponeurosis Hyperplasia. Genes. 2023; 14(9):1718. https://doi.org/10.3390/genes14091718
Chicago/Turabian StyleTajima, Rina, Atsuko Okazaki, Tsuyoshi Sato, Kokoro Ozaki, Daisuke Motooka, Yasushi Okazaki, and Tetsuya Yoda. 2023. "Genetic Landscape of Masticatory Muscle Tendon–Aponeurosis Hyperplasia" Genes 14, no. 9: 1718. https://doi.org/10.3390/genes14091718
APA StyleTajima, R., Okazaki, A., Sato, T., Ozaki, K., Motooka, D., Okazaki, Y., & Yoda, T. (2023). Genetic Landscape of Masticatory Muscle Tendon–Aponeurosis Hyperplasia. Genes, 14(9), 1718. https://doi.org/10.3390/genes14091718