
Combine with RNA-seq Reveals the Effect of Melatonin in the Synthesis of Melanin in Primary Melanocytes of Silky Fowls Black-Bone Chicken
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Culture of Melanocytes
2.2. Indirect Immunofluorescence Assay
2.3. Treat the Cells with Melatonin and CCK8 Assay
2.4. Total RNA Extraction, cDNA Synthesis, and Real-Time Quantitative PCR (RT-qPCR)
2.5. Detection of Melanin Content
2.6. Determination of Tyrosinase Activity
2.7. RNA-seq
2.8. Statistical Analysis
3. Results
3.1. Isolation and Characterization of Silky Fowls Black-Bone Chicken Melanocytes
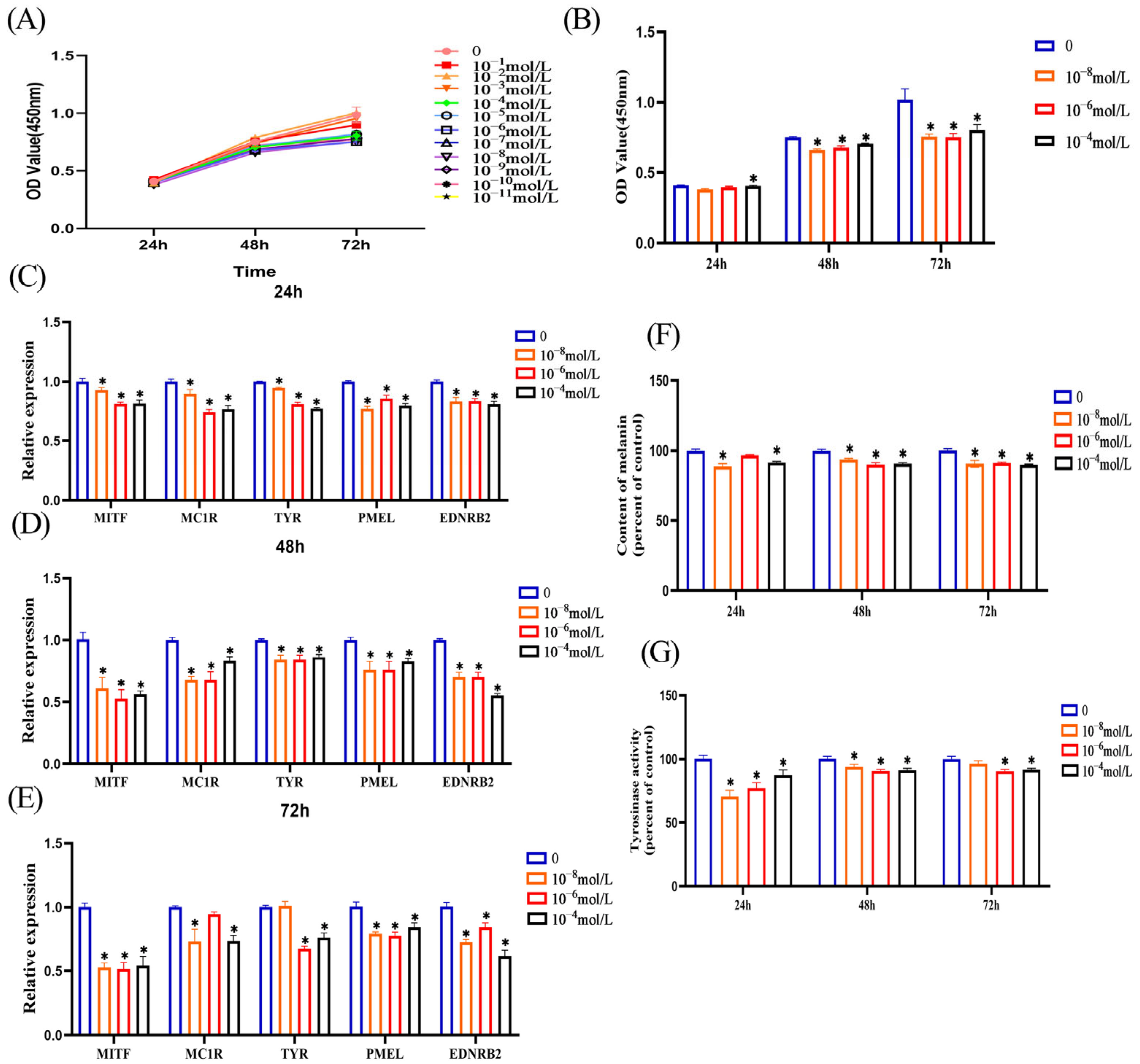
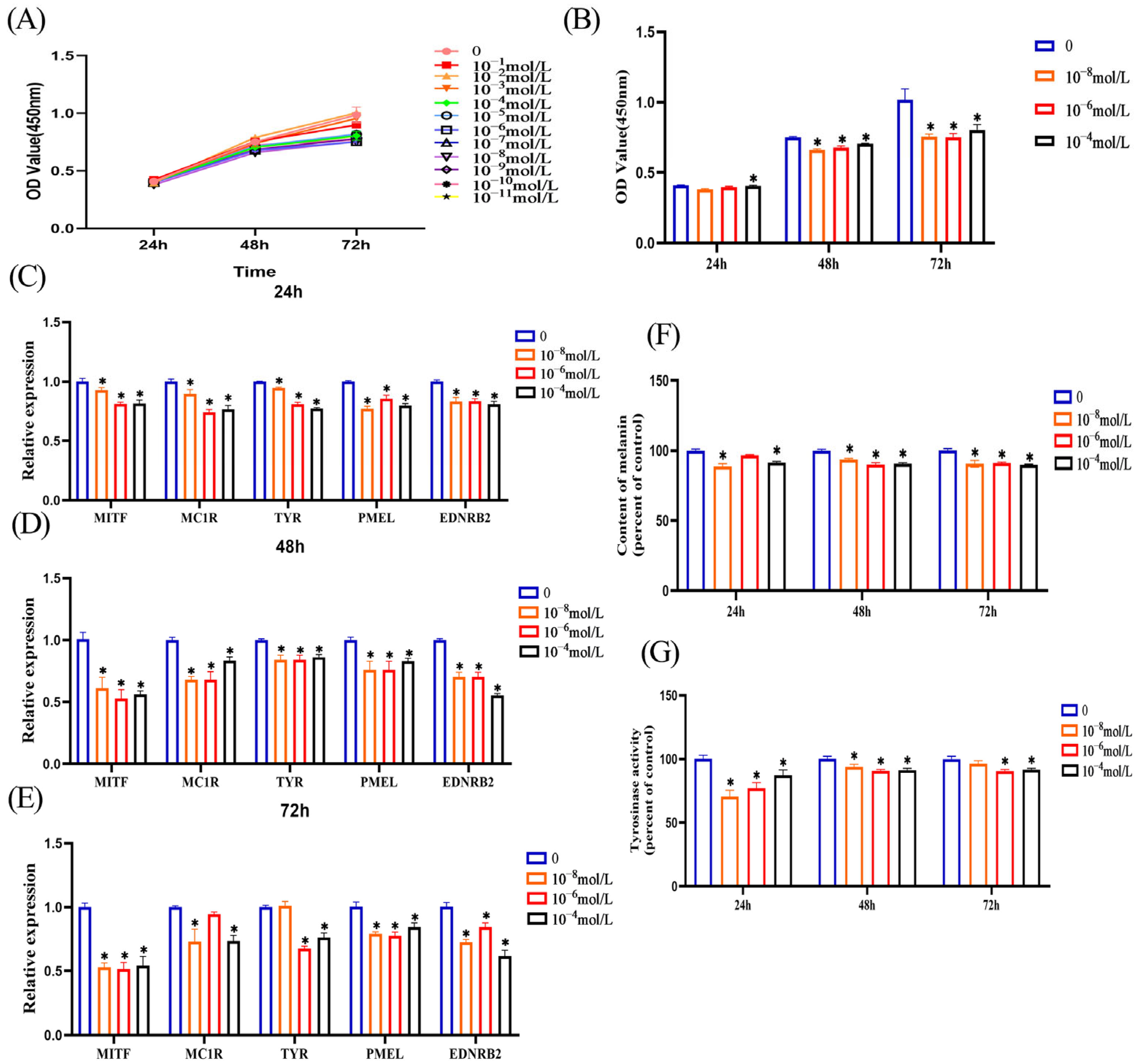
3.2. Melatonin Inhibits Melanin Synthesis in Melanocytes
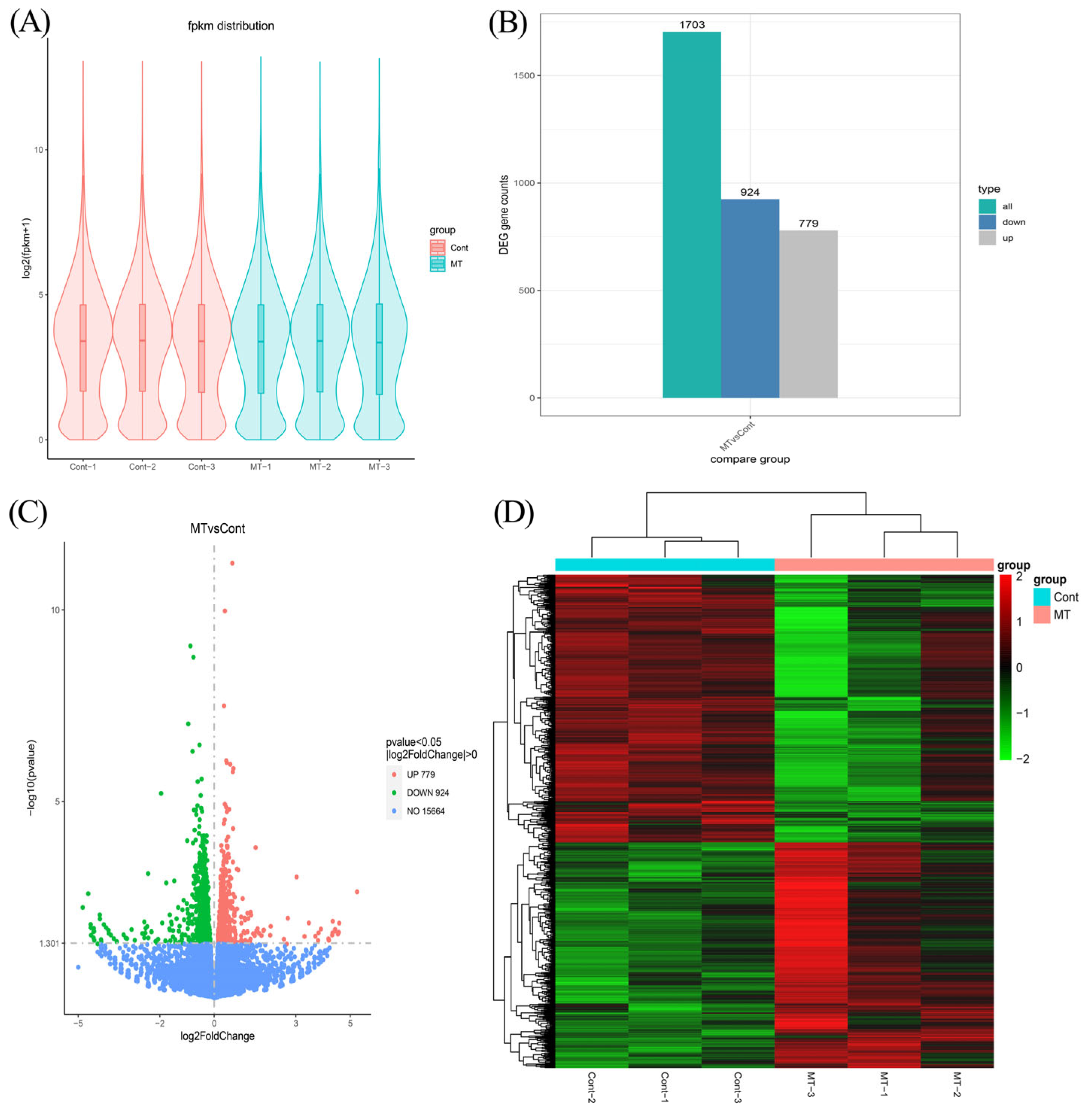
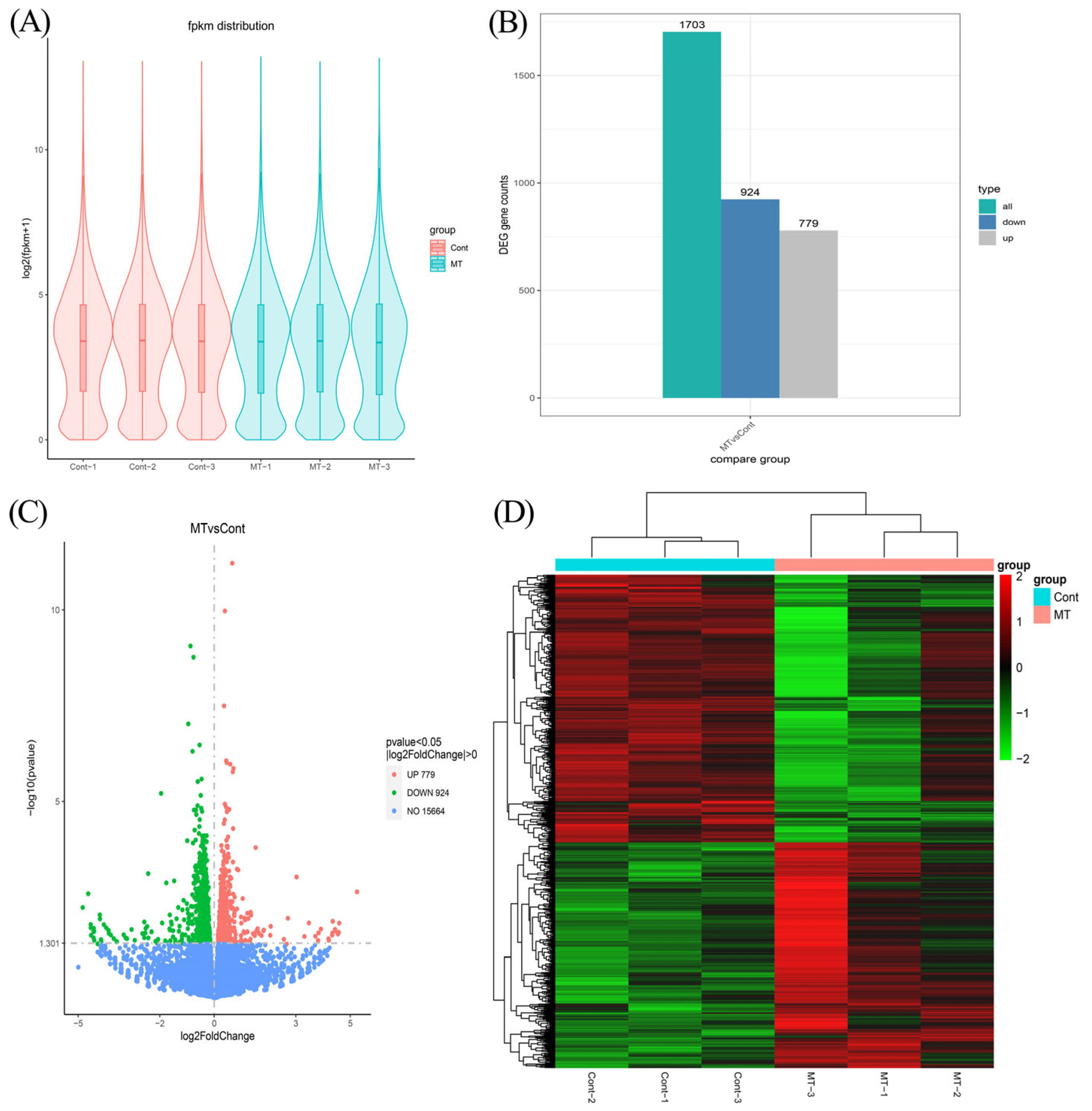
3.3. Overview of RNA Sequencing Data, Differential Expression Gene Screening
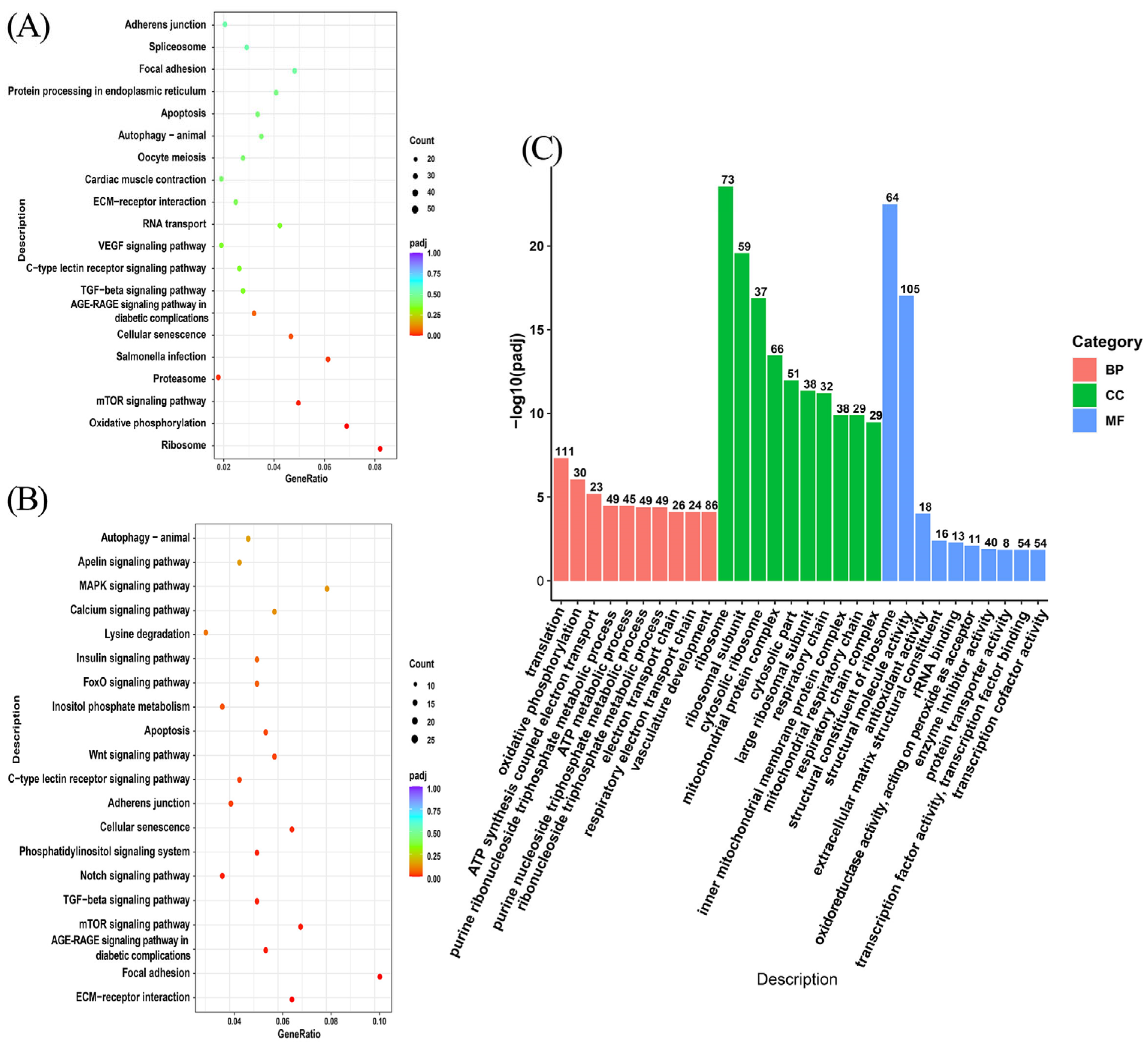
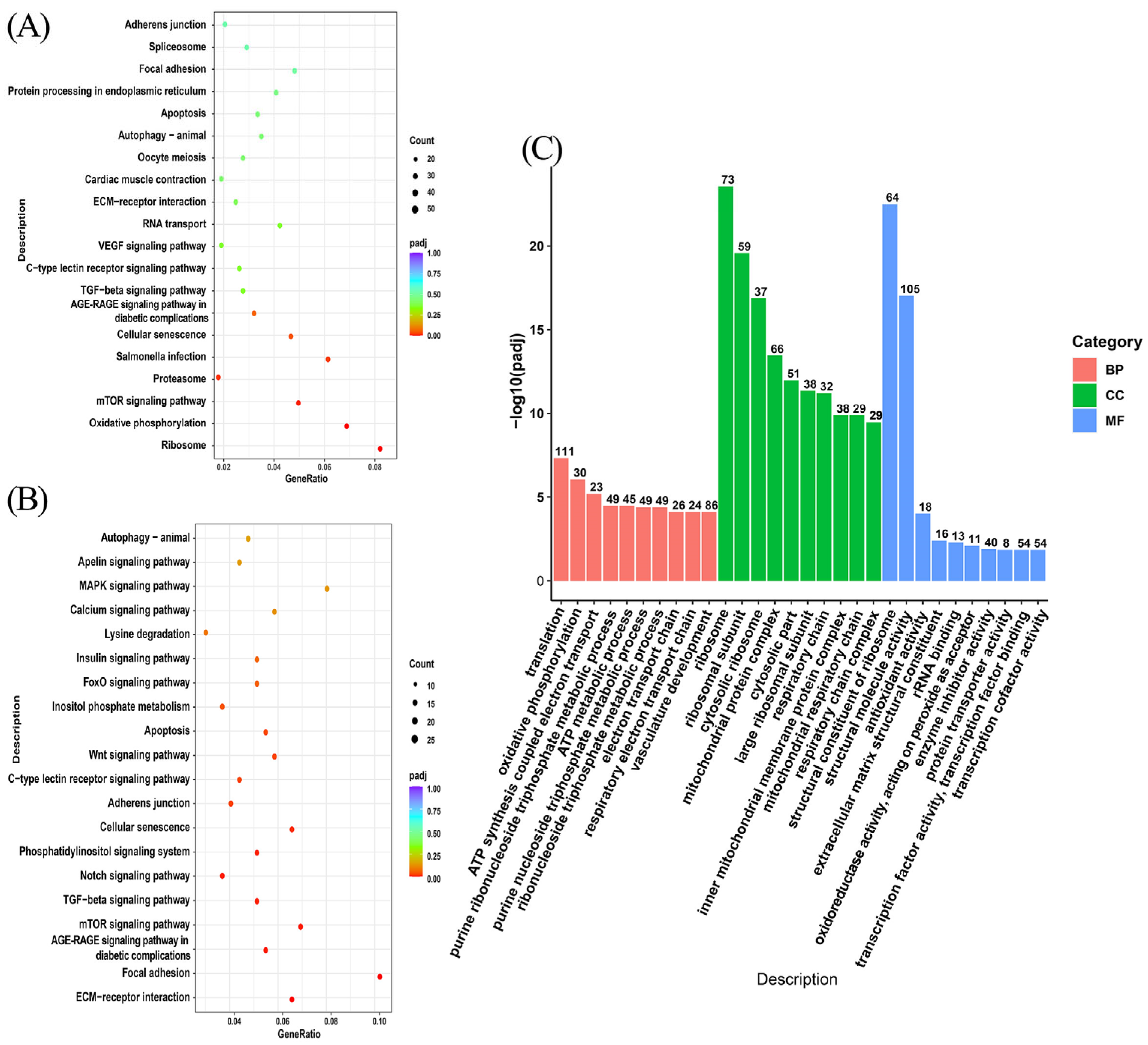
3.4. Enrichment Analysis of Differentially Expressed Genes
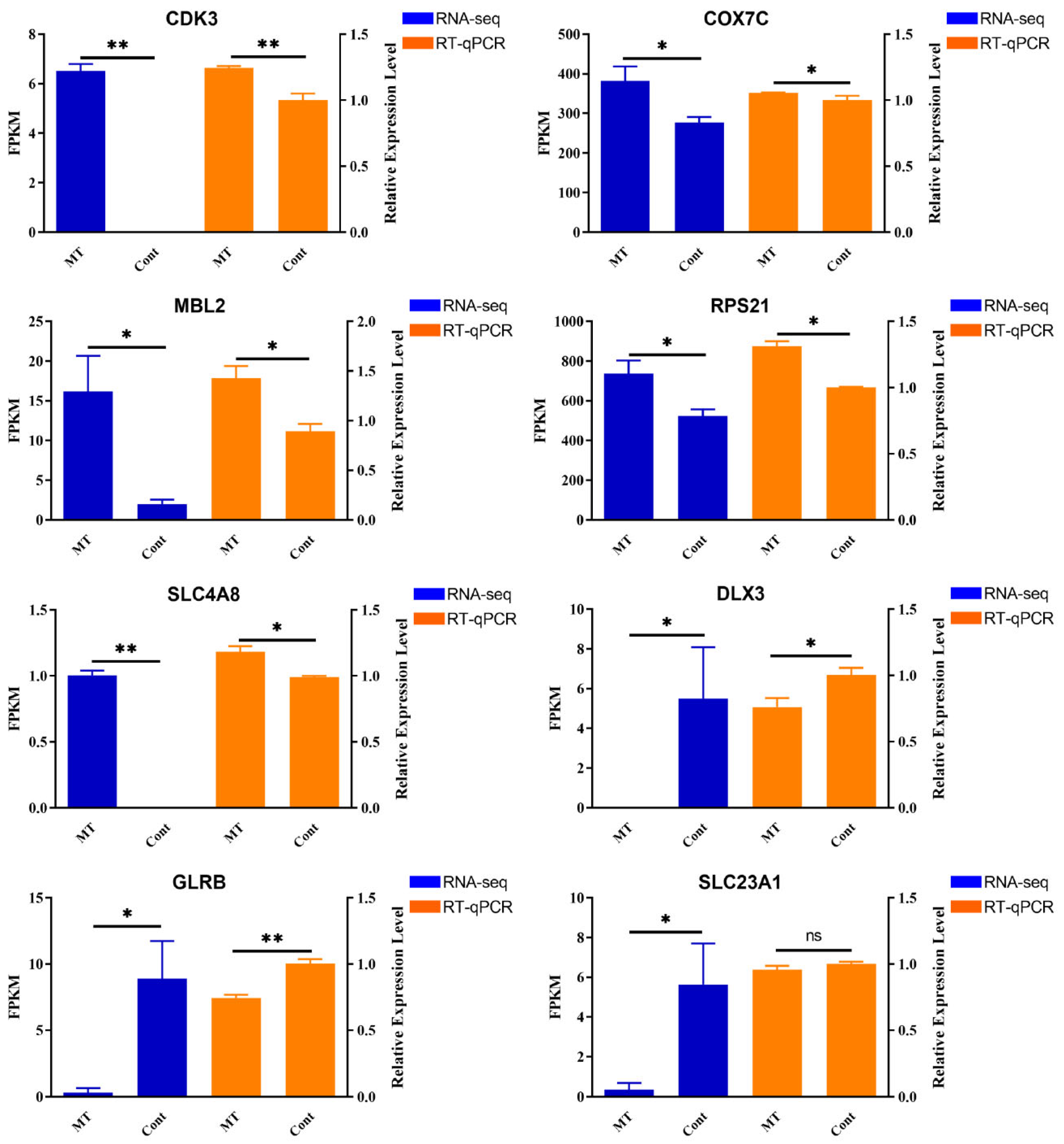
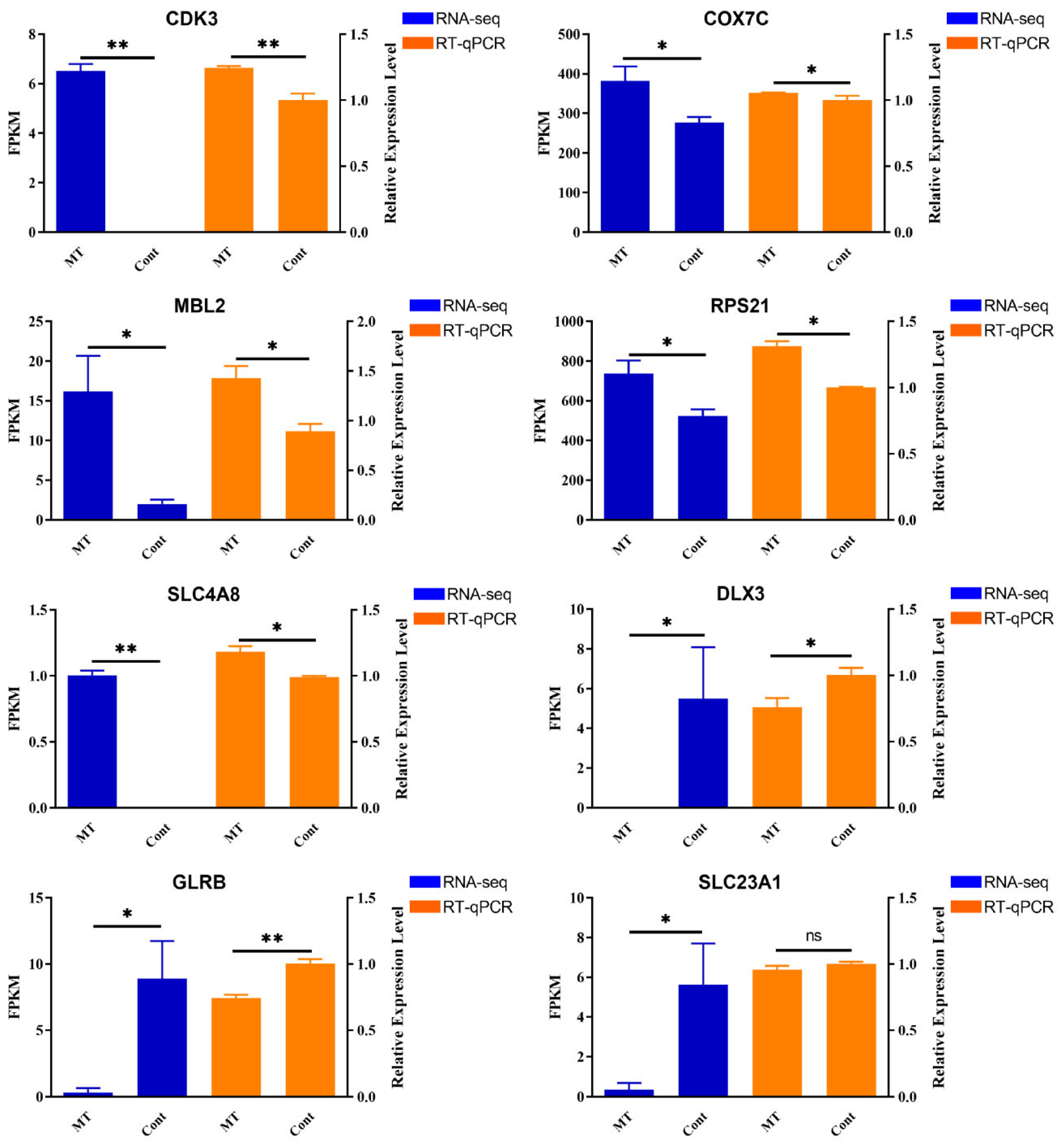
3.5. Validation of DEGs by RT-qPCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liang, L. Screening, Analysis, and Identification of Differentially Displayed (DD) Expression Sequence Tags (ESTs) in the Liver of Black Bone Chickens. Ph.D. Thesis, Sichuan Agricultural University, Sichuan, China, 2004. [Google Scholar]
- Mason, H.S. The Chemistry of Melanin. J. Biol. Chem. 1948, 172, 83–99. [Google Scholar] [CrossRef]
- Ito, S. IFPCS Presidential Lecture A Chemist’s View of Melanogenesis. Pigment Cell Res. 2003, 16, 230–236. [Google Scholar] [CrossRef]
- Wakamatsu, K.; Kavanagh, R.; Kadekaro, A.L.; Terzieva, S.; Sturm, R.A.; Leachman, S.; Abdel-Malek, Z.; Ito, S. Diversity of pigmentation in cultured human melanocytes is due to differences in the type as well as quantity of melanin. Pigment Cell Res. 2006, 19, 154–162. [Google Scholar] [CrossRef]
- Tsukamoto, V.J.H.A.K. Enzymatic control of pigmentation in mammals. FASEB J. 1991, 5, 2902–2909. [Google Scholar]
- Hanada, R.; Nakazato, M.; Matsukura, S.; Murakami, N.; Yoshimatsu, H.; Sakata, T. Differential regulation of melanin-concentrating hormone and orexin genes in the agouti-related protein/melanocortin-4 receptor system. Biochem. Biophys. Res. Commun. 2000, 268, 88–91. [Google Scholar] [CrossRef]
- Prota, G. The chemistry of melanins and melanogenesis. Fortschr. Chem. Org. Naturst 1995, 64, 93–148. [Google Scholar] [PubMed]
- Park, H.Y.; Kosmadaki, M.; Yaar, M.; Gilchrest, B.A. Cellular mechanisms regulating human melanogenesis. Cell Mol. Life Sci. 2009, 66, 1493–1506. [Google Scholar] [CrossRef]
- Thody, A.J. α-MSH and the regulation of melanocyte function. Ann. N. Y. Acad. Sci. 1999, 885, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Neurobiology, pathophysiology, and treatment of melatonin deficiency and dysfunction. Sci. World J. 2012, 2012, 640389. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Liu, D.; Di, S.; Zhang, Z.; Li, W.; Zhang, J.; Xu, L.; Guo, K.; Zhu, Y.; Li, X.; et al. Histone deacetylase 9 downregulation decreases tumor growth and promotes apoptosis in non-small cell lung cancer after melatonin treatment. J. Pineal. Res. 2019, 67, e12587. [Google Scholar] [CrossRef]
- Cipolla-Neto, J.; Amaral, F.G.D. Melatonin as a Hormone: New Physiological and Clinical Insights. Endocr. Rev. 2018, 39, 990–1028. [Google Scholar] [CrossRef] [PubMed]
- Perdomo, J.; Quintana, C.; Gonzalez, I.; Hernandez, I.; Rubio, S.; Loro, J.F.; Reiter, R.J.; Estevez, F.; Quintana, J. Melatonin Induces Melanogenesis in Human SK-MEL-1 Melanoma Cells Involving Glycogen Synthase Kinase-3 and Reactive Oxygen Species. Int. J. Mol. Sci. 2020, 21, 4970. [Google Scholar] [CrossRef]
- Kleszczynski, K.; Kim, T.K.; Bilska, B.; Sarna, M.; Mokrzynski, K.; Stegemann, A.; Pyza, E.; Reiter, R.J.; Steinbrink, K.; Bohm, M.; et al. Melatonin exerts oncostatic capacity and decreases melanogenesis in human MNT-1 melanoma cells. J. Pineal. Res. 2019, 67, e12610. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Shuaishuai, H. Study on the Mechanism of Deubiquitination Enzyme USP13 Regulating Melanin Production in Rex Rabbits. Ph.D. Thesis, Yangzhou University, Yangzhou, China, 2022. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Eisinger, M.; Marko, O. Selective proliferation of normal human melanocytes in vitro in the presence of phorbol ester and cholera toxin. Proc. Natl. Acad. Sci. USA 1982, 79, 2018–2022. [Google Scholar] [CrossRef] [PubMed]
- Bennett, D.C.; Cooper, P.J.; Dexter, T.J.; Devlin, L.M.; Heasman, J.; Nester, B. Cloned mouse melanocyte lines carrying the germline mutations albino and brown complementation in culture. Development 1989, 105, 379–385. [Google Scholar] [CrossRef]
- Xiong, M.; Xu, L.; Qu, M.; Li, G. Adenosine cyclic phosphate promotes melanin synthesis in skin melanocytes of Taihe black bone chickens. Chin. J. Cell Biol. 2018, 40, 1363–1368. [Google Scholar] [CrossRef]
- Brzezinska, K.A.; Bhardwaj, S.; Teng, M.S.; Si, Q.; Sun, J.; Westra, W.H.; Zakowski, M.F.; Szporn, A.H. Melanotic medullary thyroid carcinoma: A case report with review of the literature. Diagn Cytopathol. 2023, 51, E14–E20. [Google Scholar] [CrossRef]
- Ikeda, Y.; Wada, A.; Hasegawa, T.; Yokota, M.; Koike, M.; Ikeda, S. Melanocyte progenitor cells reside in human subcutaneous adipose tissue. PLoS ONE 2021, 16, e0256622. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.Y.; Ngo, H.H.; Choi, W.J.; Keum, Y.S. Dimethyl Itaconate Inhibits Melanogenesis in B16F10 Cells. Antioxidants 2023, 12, 692. [Google Scholar] [CrossRef] [PubMed]
- Opdecamp, K.; Nakayama, A.; Nguyen, M.T.T.; Hodgkinson, C.A.; Pavan, W.J.; Arnheiter, H. Melanocyte development in vivo and in neural crest cell cultures crucial dependence on the Mitf basic-helix-loop-helix-zipper transcription factor. Development 1997, 124, 2377–2386. [Google Scholar] [CrossRef]
- Zhu, H.; Zhao, Z.J.; Liu, H.Y.; Cai, J.; Lu, Q.K.; Ji, L.D.; Xu, J. The melatonin receptor 1B gene links circadian rhythms and type 2 diabetes mellitus: An evolutionary story. Ann. Med. 2023, 55, 1262–1286. [Google Scholar] [CrossRef] [PubMed]
- Dubocovich, M.L. Melatonin receptors: Role on sleep and circadian rhythm regulation. Sleep Med. 2007, 8, 34–42. [Google Scholar] [CrossRef]
- Kleszczynski, K.; Bilska, B.; Stegemann, A.; Flis, D.J.; Ziolkowski, W.; Pyza, E.; Luger, T.A.; Reiter, R.J.; Bohm, M.; Slominski, A.T. Melatonin and Its Metabolites Ameliorate UVR-Induced Mitochondrial Oxidative Stress in Human MNT-1 Melanoma Cells. Int. J. Mol. Sci. 2018, 19, 3786. [Google Scholar] [CrossRef]
- Zhao, C.N.; Wang, P.; Mao, Y.M.; Dan, Y.L.; Wu, Q.; Li, X.M.; Wang, D.G.; Davis, C.; Hu, W.; Pan, H.F. Potential role of melatonin in autoimmune diseases. Cytokine Growth Factor Rev. 2019, 48, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Pruski, D. Melatonin inhibits proliferation and melanogenesis in rodent melanoma cells. Exp. Cell Res. 1993, 206, 189–194. [Google Scholar] [CrossRef]
- Slominski, R.M.; Reiter, R.J.; Schlabritz-Loutsevitch, N.; Ostrom, R.S.; Slominski, A.T. Melatonin membrane receptors in peripheral tissues: Distribution and functions. Mol. Cell Endocrinol. 2012, 351, 152–166. [Google Scholar] [CrossRef]
- Ma, L.P.; Liu, M.M.; Liu, F.; Sun, B.; Wang, S.N.; Chen, J.; Yu, H.J.; Yan, J.; Tian, M.; Gao, L.; et al. Melatonin inhibits senescence-associated melanin pigmentation through the p53-TYR pathway in human primary melanocytes and the skin of C57BL/6 J mice after UVB irradiation. J. Mol. Med. 2023, 101, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Lin, Z.; Tidwell, W.J.; Li, W.; Slominski, A.T. Melatonin and its metabolites accumulate in the human epidermis in vivo and inhibit proliferation and tyrosinase activity in epidermal melanocytes in vitro. Mol. Cell Endocrinol. 2015, 404, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zeng, H.; Jiang, L.; Fu, C.; Gao, L.; Zhang, L.; Zhang, Y.; Zhang, X.; Zhu, L.; Zhang, F.; et al. Melatonin reduces melanogenesis by inhibiting the paracrine effects of keratinocytes. Exp. Dermatol. 2023, 32, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Sevilla, A.; Cheret, J.; Lee, W.; Paus, R. Concentration-dependent stimulation of melanin production as well as melanocyte and keratinocyte proliferation by melatonin in human eyelid epidermis. Exp. Dermatol. 2023, 32, 684–693. [Google Scholar] [CrossRef]
- Sevilla, A.; Cheret, J.; Slominski, R.M.; Slominski, A.T.; Paus, R. Revisiting the role of melatonin in human melanocyte physiology: A skin context perspective. J. Pineal Res. 2022, 72, e12790. [Google Scholar] [CrossRef]
- Shen, Q.; Jiang, Y.; Jia, X.; Zhou, X.; Zhou, Q.H. Amelioratory Effect of Melatonin on Cognition Dysfunction Induced by Sevoflurane Anesthesia in Aged Mice. Iran. J. Pharm. Res. 2022, 21, e133971. [Google Scholar] [CrossRef]
- Yang, Z.; Zeng, B.; Pan, Y.; Huang, P.; Wang, C. Autophagy participates in isoliquiritigenin-induced melanin degradation in human epidermal keratinocytes through PI3K/AKT/mTOR signaling. Biomed. Pharmacother. 2018, 97, 248–254. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) | Product Size (bp) | Accession Number |
|---|---|---|---|
| MITF | F: TGTGACTGAACCAACTGGCACTTAC | 157 | ENSGALT00010053658.1 |
| R: GCTCCGCCTGCTACTCGTT | 191 | ENSGALT00000077972.3 | |
| PMEL | F: TTGTCTACGTGTGGTGGAC | ||
| R: CTGGTCGGTGATGCTGAACT | |||
| EDNRB2 | F: GAGGAAGTTTAATTCACTAGGAACC | 104 | ENSGALT00010029474.1 |
| R: TTGCTTGGGTCTTGGTCTGAT | |||
| TYR | F: CACTCTTAGGTGGCTCCAATGTG | 154 | ENSGALT00010018719.1 |
| R: CAGTCCCAGTAGGGGATGGTGAA | |||
| MC1R | F: GCCCTTCTTCTTCCACCTCAT | 83 | ENSGALT00010023383.1 |
| R: AGAGGTTGAAATAGCTGAAGAAGCA | |||
| CDK3 | F: ACTTGAAGCCACAGAACTTGC | 251 | ENSGALT00010071972.1 |
| R: ATCGATCTCAGAGTCCCCTT | |||
| SLC4A8 | F: TTGCCTACAAAGCCAAGGACCG | 259 | ENSGALT00010062208.1 |
| R: CGCTTCACATCCAGGATCAAACC | |||
| MBL2 | F: TCCTGCAGTCAATGGATTACCAG | 127 | ENSGALT00000143518.1 |
| R: TTTTAATCCTTGGGGTCCTG | |||
| RPS21 | F: GATCTCTACGTGCCTCGGAA | 162 | ENSGALT00010051720.1 |
| R: CCCCATCCTACGAATTGCT | |||
| COX7C | F: TTCACTACCTCCGCCCTTCGT | 109 | ENSGALT00010014887.1 |
| R: AGAATGCACACATCATTGCCAGT | |||
| GLRB | F: GACTCAAACTGCCCAACGA | 219 | ENSGALT00010019616.1 |
| R: TCAAGTCCAAAGGGCACGA | |||
| DLX3 | F: ATTACAGCGGCCAGCACGACT | 243 | ENSGALT00010063622.1 |
| R: TCCCGTTGACCATCCGCACCT | |||
| SLC23A1 | F: CACCCTCATCCAGACCACCGT R: ACCCAGTTGCCATAGATCTGCT | 141 | ENSGALT00010019860.1 |
| GAPDH | F: CGATCTGAACTACATGGTTTAC | 153 | ENSGALT00000114062.1 |
| R: TCTGCCCATTTGATGTTGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, T.; Qiu, L.; Chen, S.; Wang, Z.; Jiang, Y.; Bai, H.; Bi, Y.; Chang, G. Combine with RNA-seq Reveals the Effect of Melatonin in the Synthesis of Melanin in Primary Melanocytes of Silky Fowls Black-Bone Chicken. Genes 2023, 14, 1648. https://doi.org/10.3390/genes14081648
Yang T, Qiu L, Chen S, Wang Z, Jiang Y, Bai H, Bi Y, Chang G. Combine with RNA-seq Reveals the Effect of Melatonin in the Synthesis of Melanin in Primary Melanocytes of Silky Fowls Black-Bone Chicken. Genes. 2023; 14(8):1648. https://doi.org/10.3390/genes14081648
Chicago/Turabian StyleYang, Ting, Lingling Qiu, Shihao Chen, Zhixiu Wang, Yong Jiang, Hao Bai, Yulin Bi, and Guobin Chang. 2023. "Combine with RNA-seq Reveals the Effect of Melatonin in the Synthesis of Melanin in Primary Melanocytes of Silky Fowls Black-Bone Chicken" Genes 14, no. 8: 1648. https://doi.org/10.3390/genes14081648
APA StyleYang, T., Qiu, L., Chen, S., Wang, Z., Jiang, Y., Bai, H., Bi, Y., & Chang, G. (2023). Combine with RNA-seq Reveals the Effect of Melatonin in the Synthesis of Melanin in Primary Melanocytes of Silky Fowls Black-Bone Chicken. Genes, 14(8), 1648. https://doi.org/10.3390/genes14081648