
Effects of Forest Fragmentation on Connectivity and Genetic Diversity in an Endemic and an Invasive Rodent in Northwestern Madagascar

 , , , ,
, , , , 
Abstract
1. Introduction
2. Materials and Methods
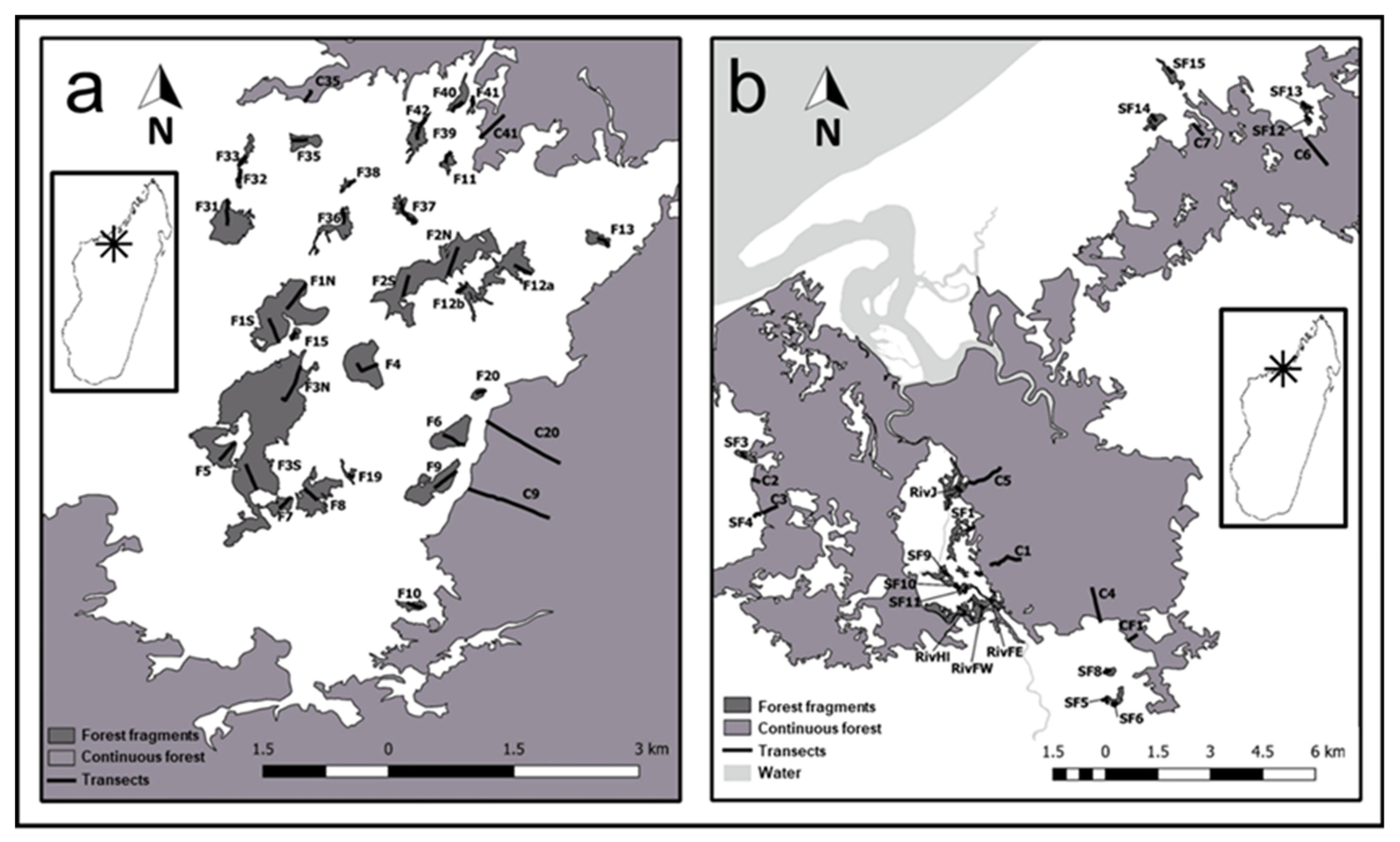
2.1. Study Site and Sampling
2.2. Laboratory Procedures and Data Preparation
2.3. Analyses of Local Movement and Dispersal
2.4. Analyses of Connectivity and Genetic Structure on the Landscape Level
2.5. Ecological Determinants of Genetic Diversity
3. Results
3.1. RADseq Dataset
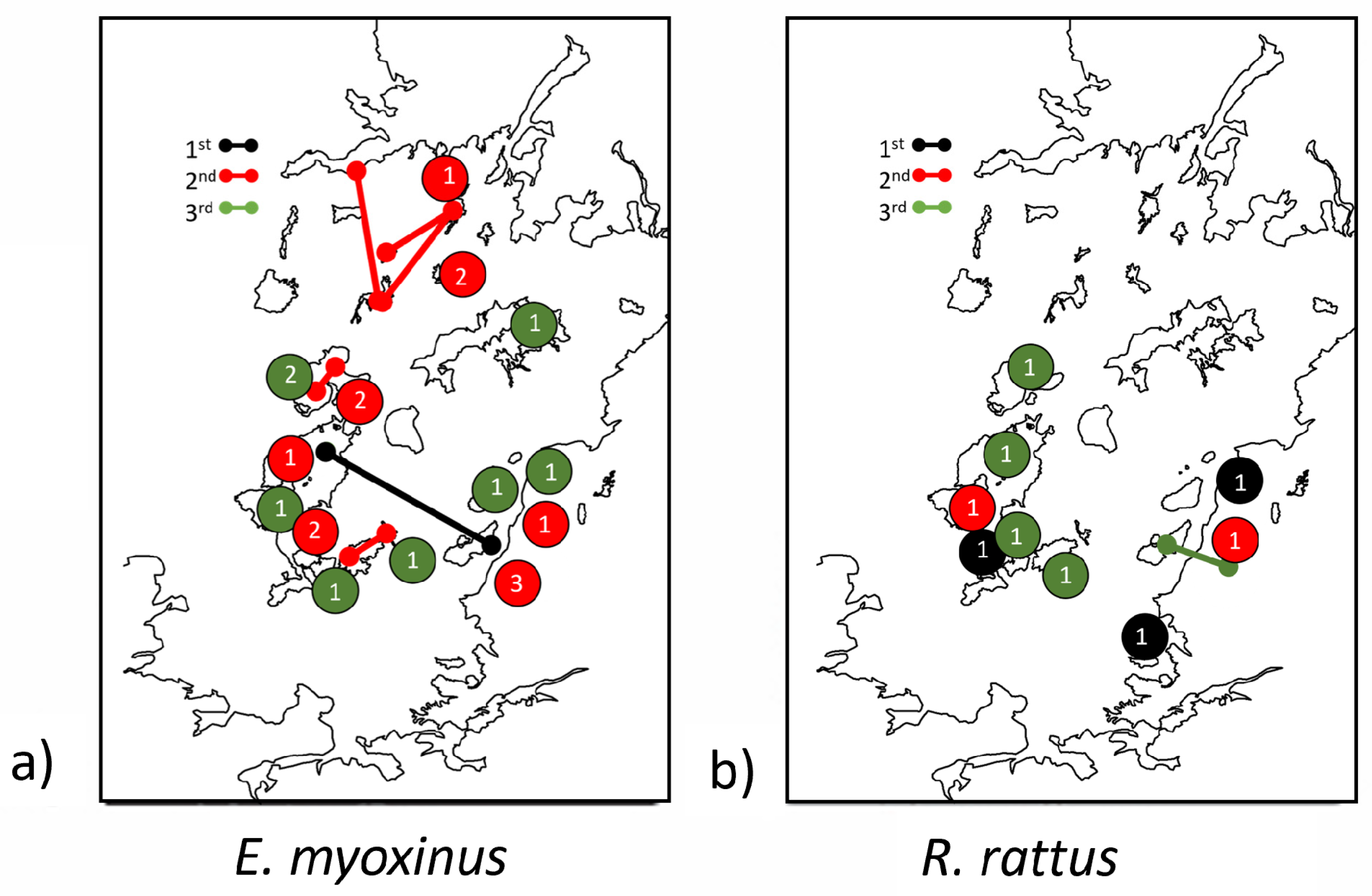
3.2. Patterns of Local Movement and Dispersal
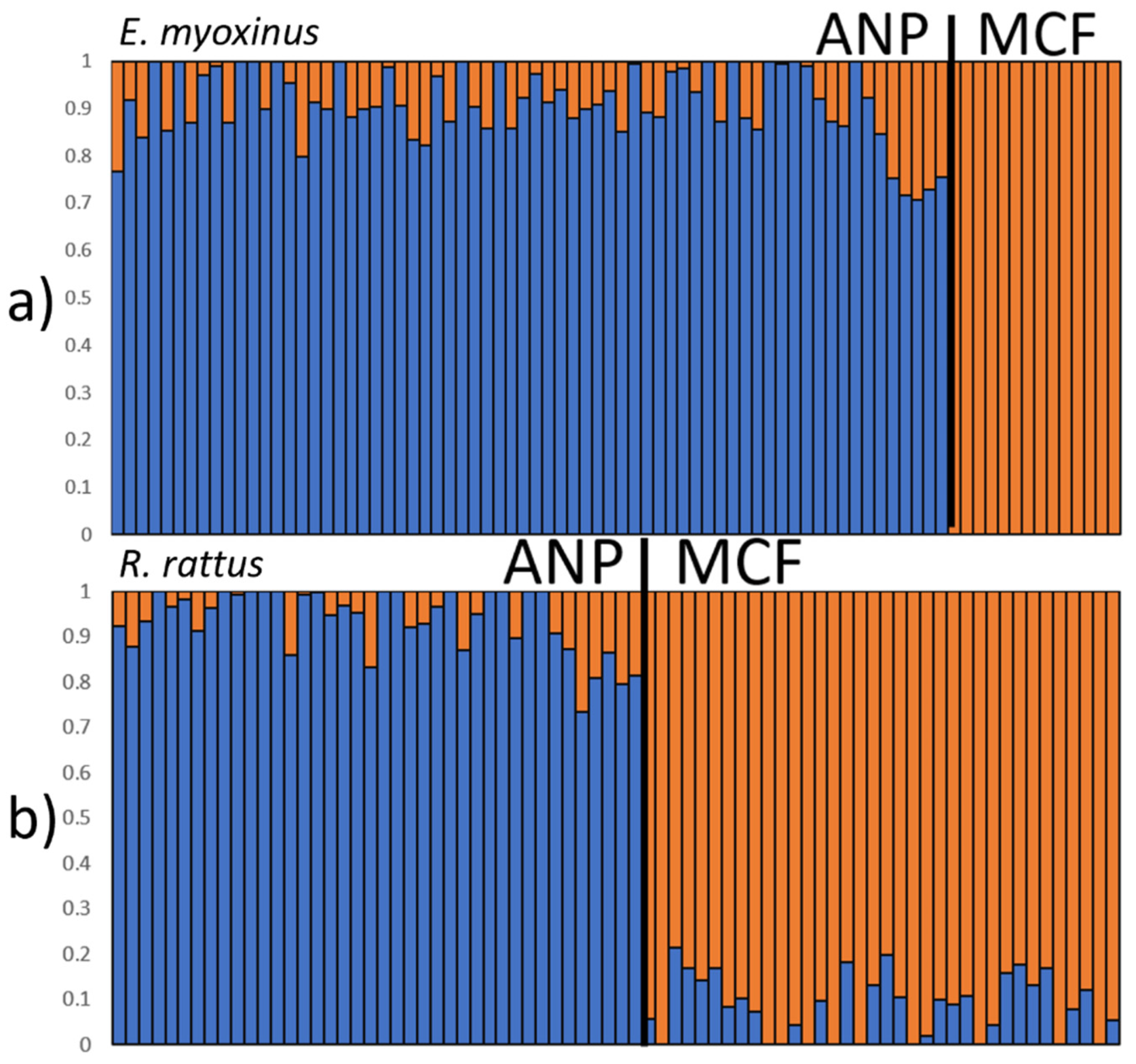
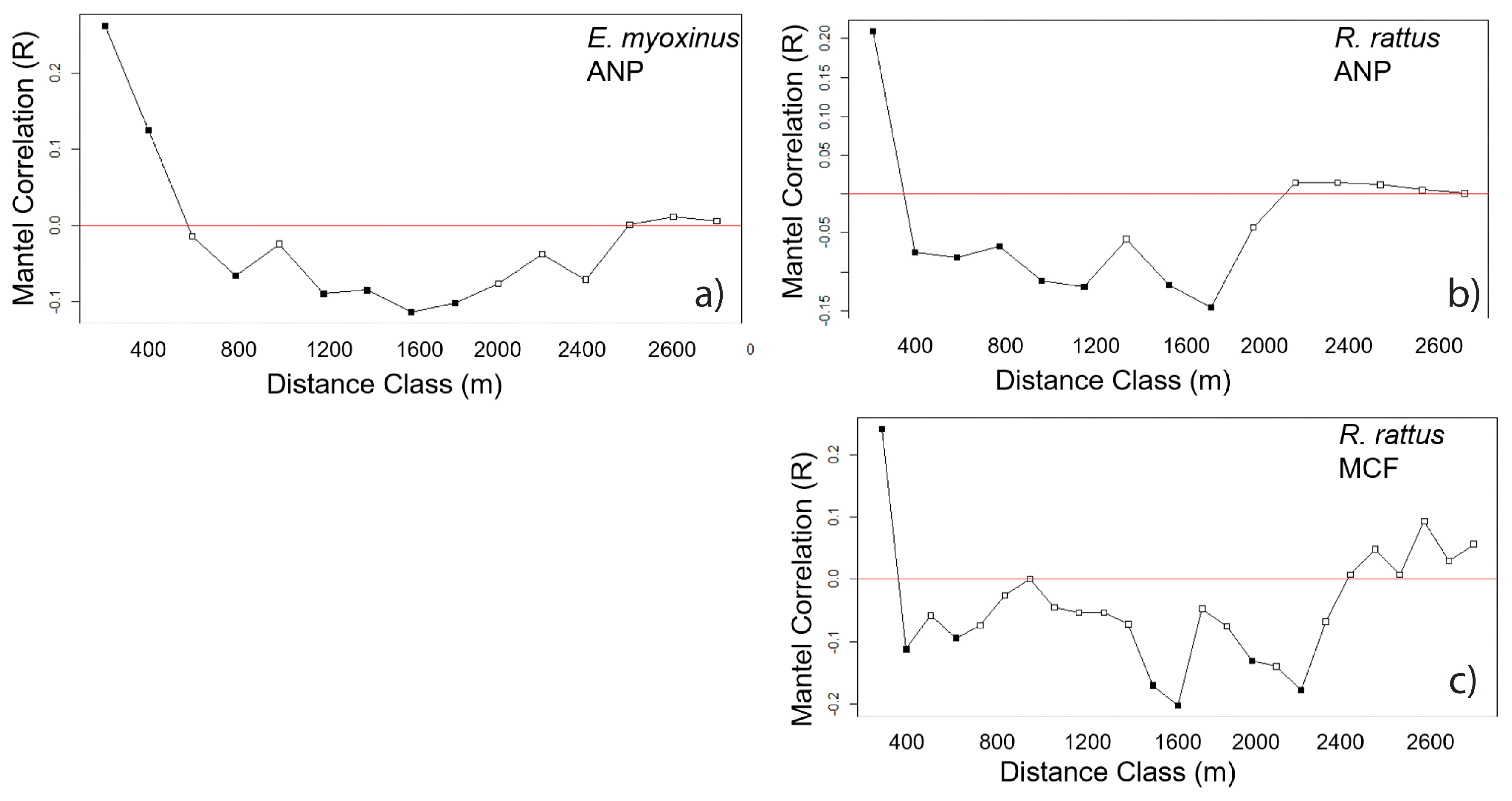
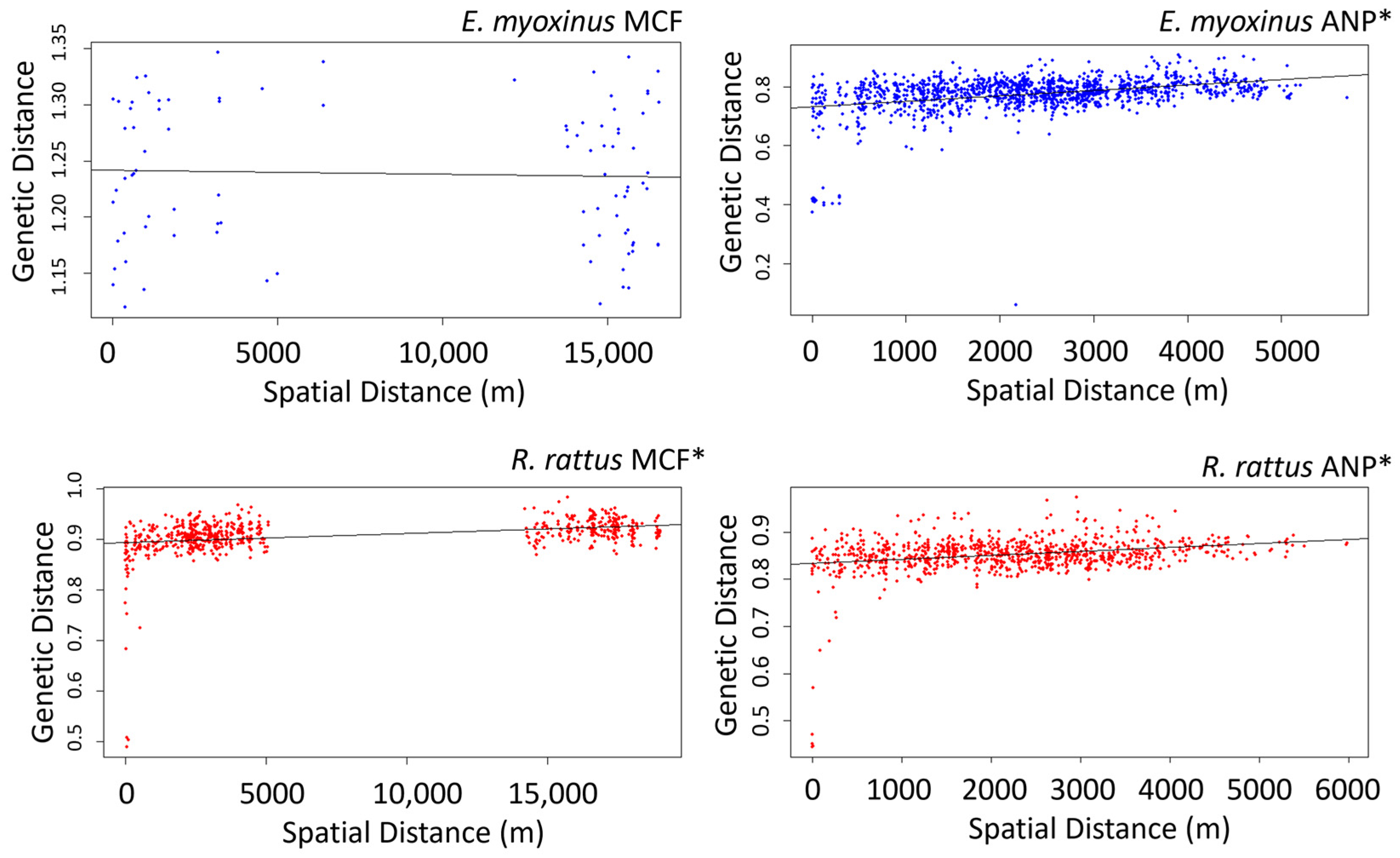
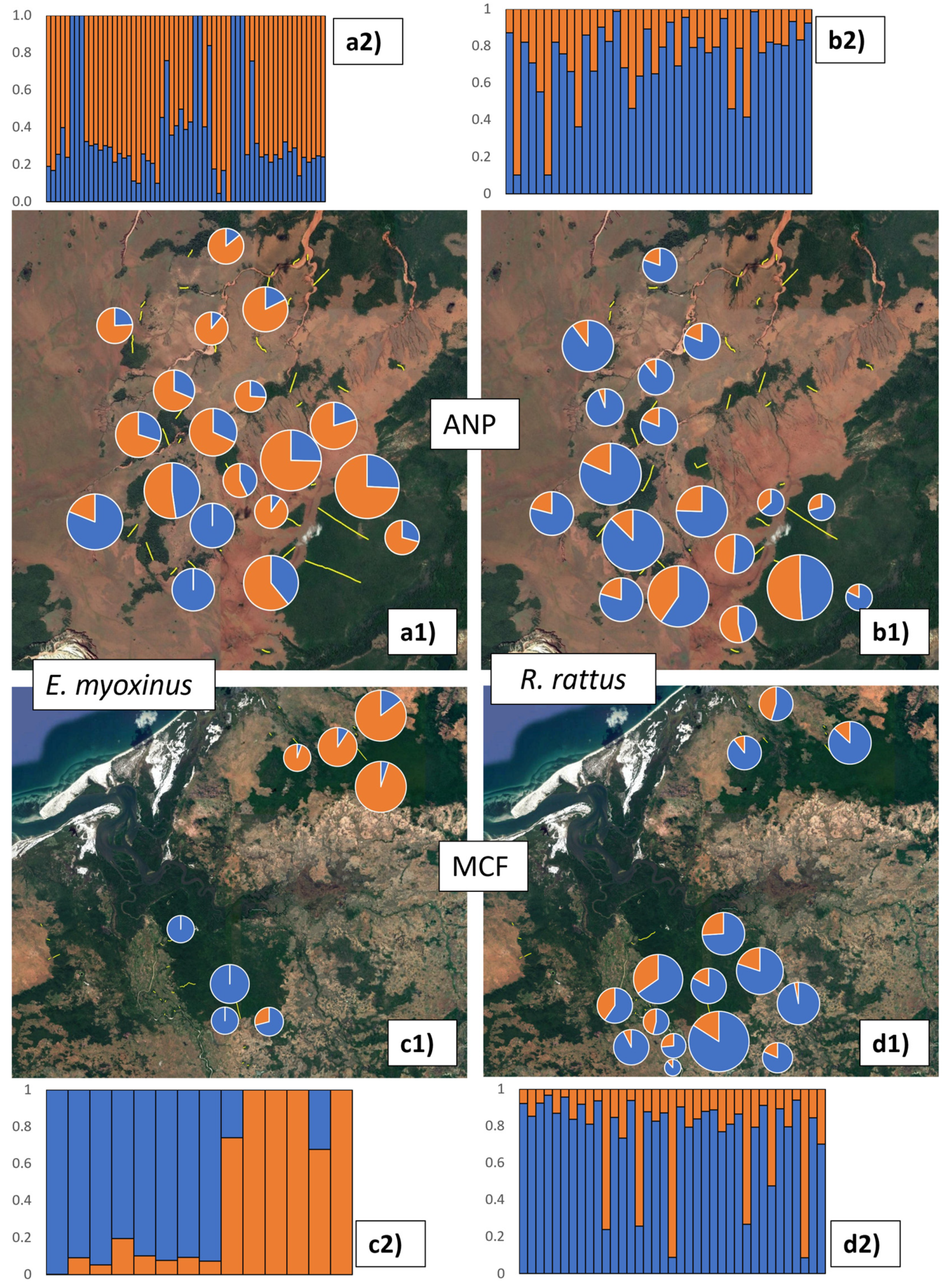
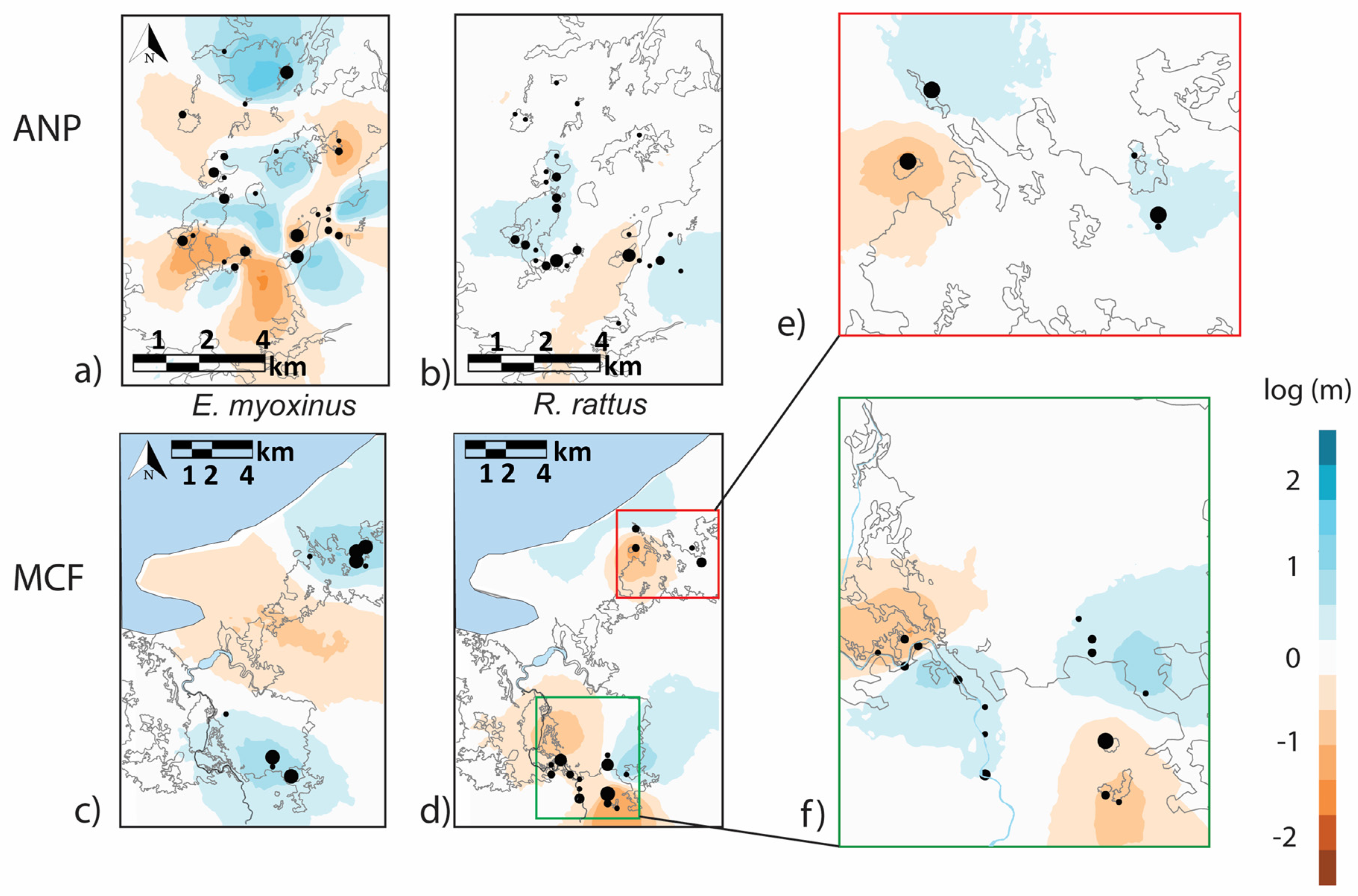
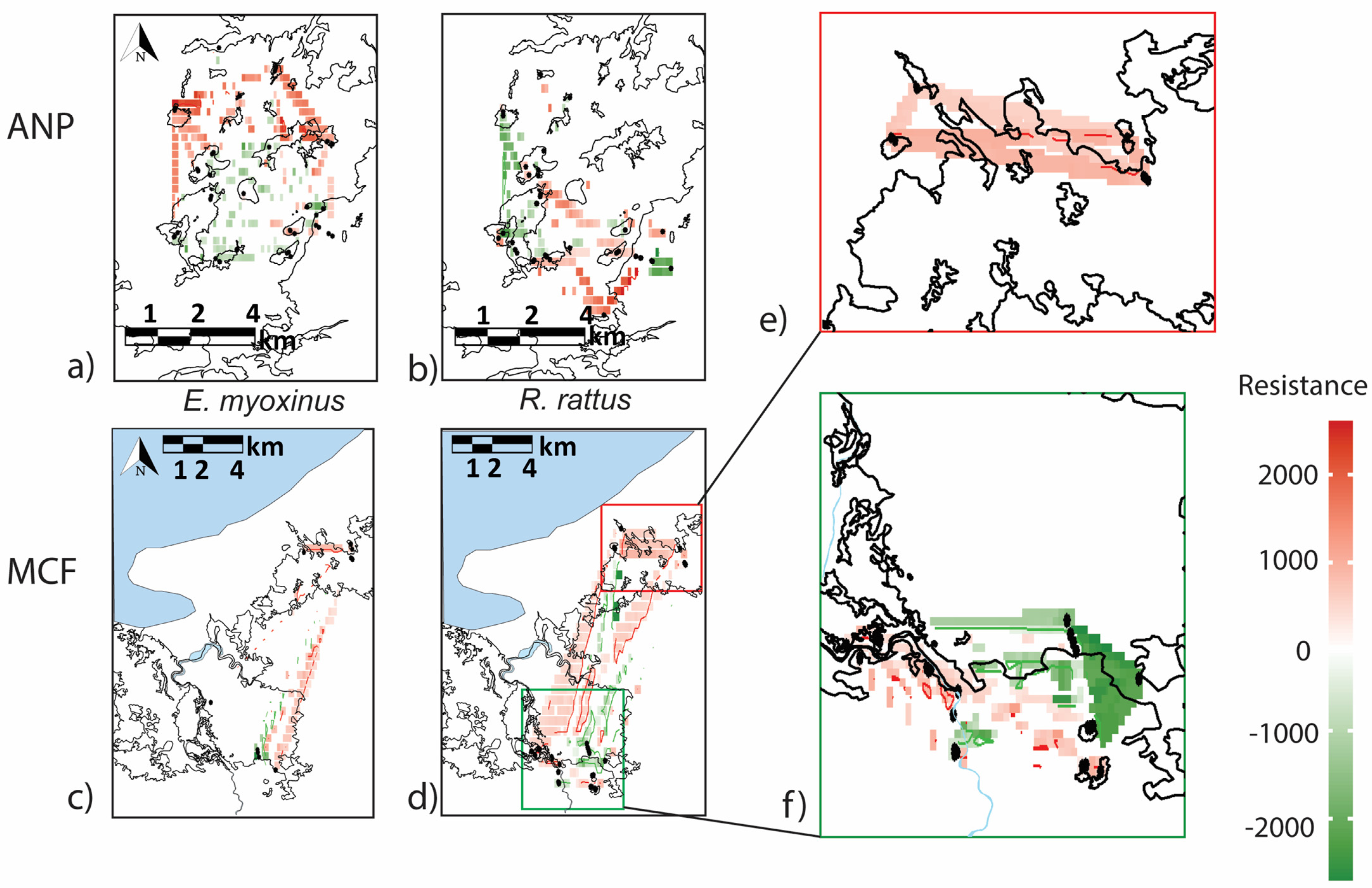
3.3. Connectivity and Genetic Structure on the Landscape Level
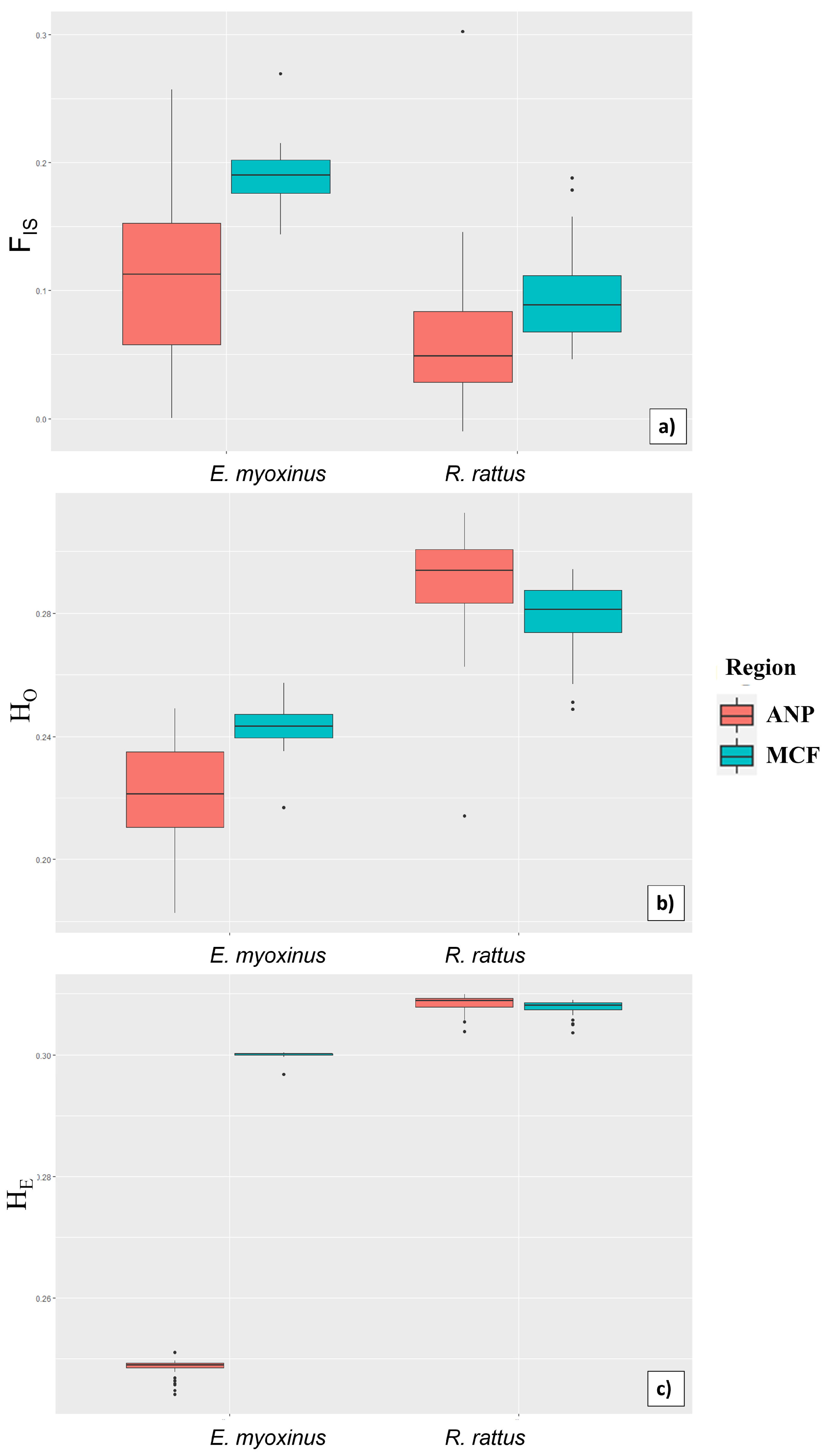
3.4. Ecological Determinants of Genetic Diversity
4. Discussion
4.1. Do Endemic and Invasive Rodents Use Non-Forest Matrices for Local Movement or Dispersal?
4.2. Are Connectivity and Genetic Structure Affected Primarily by Isolation by Distance or by Habitat?
4.3. What Ecological Factors Affect the Genetic Diversity of Rodent Sub-Populations?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilmé, L.; Goodman, S.; Ganzhorn, J. Biogeographic evolution of Madagascar’s microendemic biota. Science 2006, 312, 1063–1065. [Google Scholar] [CrossRef] [PubMed]
- Yoder, A.; Nowak, M. Has vicariance or dispersal been the predominant biogeographic force in Madagascar? Only time will tell. Ann. Rev. Ecol. Evol. Syst. 2006, 37, 405–431. [Google Scholar]
- Vences, M.; Wollenberg Valero, K.; Vieites, D.; Lees, D. Madagascar as a model region of species diversification. Trends Ecol. Evol. 2009, 24, 456–465. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Schwitzer, C.; Mittermeier, R.A.; Johnson, S.E.; Donati, G.; Irwin, M.; Peacock, H.; Ratsimbazafy, J.; Razafindramanana, J.; Louis Jr, E.E.; Chikhi, L.; et al. Averting lemur extinctions amid Madagascar’s political crisis. Science 2014, 343, 842–843. [Google Scholar] [CrossRef]
- Vieilledent, G.; Grinand, C.; Rakotomalala, F.A.; Ranaivosoa, R.; Rakotoarijaona, J.R.; Allnutt, T.F.; Achard, F. Combining global tree cover loss data with historical national forest cover maps to look at six decades of deforestation and forest fragmentation in Madagascar. Biol. Conserv. 2018, 222, 189–197. [Google Scholar] [CrossRef]
- Harcourt, A.H.; Doherty, D.A. Species-area relationships of primates in tropical forest fragments: A global analysis. J. Appl. Ecol. 2005, 42, 630–637. [Google Scholar] [CrossRef]
- Villard, M.A.; Metzger, J.P. Review: Beyond the fragmentation debate: A conceptual model to predict when habitat configuration really matters. J. Appl. Ecol. 2014, 51, 309–318. [Google Scholar] [CrossRef]
- Watling, J.I.; Arroyo-Rodríguez, V.; Pfeifer, M.; Baeten, L.; Banks-Leite, C.; Cisneros, L.M.; Fang, R.; Hamel-Leigue, A.C.; Lachat, T.; Leal, I.R.; et al. Support for the habitat amount hypothesis from a global synthesis of species density studies. Ecol. Letters. 2020, 23, 674–681. [Google Scholar] [CrossRef]
- Fahrig, L. Ecological responses to habitat fragmentation per se. Ann. Rev. Ecol. Evol. Syst. 2017, 48, 1–23. [Google Scholar] [CrossRef]
- Ries, L.; Murphy, S.M.; Wimp, G.M.; Fletcher, R.J. Closing persistent gaps in knowledge about edge ecology. Curr. Landsc. Ecol. Rep. 2017, 2, 30–41. [Google Scholar] [CrossRef]
- Malcolm, J.R.; Valenta, K.; Lehman, S.M. Edge effects in tropical dry forests of Madagascar: Additivity or synergy? Landsc. Ecol. 2016, 32, 327–341. [Google Scholar] [CrossRef]
- Keinath, D.A.; Doak, D.F.; Hodges, K.E.; Prugh, L.R.; Fagan, W.; Sekercioglu, C.H.; Buchart, S.H.; Kauffman, M. A global analysis of traits predicting species sensitivity to habitat fragmentation. Global Ecol. Biogeogr. 2017, 26, 115–127. [Google Scholar] [CrossRef]
- Irwin, M.T.; Wright, P.C.; Birkinshaw, C.; Fisher, B.L.; Gardner, C.J.; Glos, J.; Goodman, S.M.; Loiselle, P.; Robeson, P.; Raharison, J.L.; et al. Patterns of species change in anthropogenically disturbed forests of Madagascar. Biol. Conserv. 2010, 143, 2351–2362. [Google Scholar]
- Eppley, T.M.; Santini, L.; Tinsman, J.C.; Donati, G. Do functional traits offset the effects of fragmentation? The case of large-bodied diurnal lemur species. Am. J. Primatol. 2020, 82, e23104. [Google Scholar] [CrossRef] [PubMed]
- Lehtinen, R.M.; Ramanamanjato, J.B.; Raveloarison, J.G. Edge effects and extinction proneness in a herpetofauna from Madagascar. Biodiv. Conserv. 2003, 12, 1357–1370. [Google Scholar] [CrossRef]
- Lehman, S.M.; Rajaonson, A.; Day, S. Edge effects on the density of Cheirogaleus major. Int. J. Prim. 2006, 27, 1569–1588. [Google Scholar] [CrossRef]
- Andriamandimbiarisoa, L.; Blanthorn, T.S.; Ernest, R.; Ramanamanjato, J.B.; Randriatafika, F.; Ganzhorn, J.U.; Donati, G. Habitat corridor utilization by the gray mouse lemur, Microcebus murinus, in the littoral forest fragments of southeastern Madagascar. Mad. Conserv. Develop. 2015, 10, 144–150. [Google Scholar] [CrossRef]
- Steffens, T.S.; Lehman, S.M. Lemur species-specific metapopulation responses to habitat loss and fragmentation. PLoS ONE 2018, 13, e0195791. [Google Scholar] [CrossRef] [PubMed]
- Ganzhorn, J.U. Effects of introduced Rattus rattus on endemic small mammals in dry deciduous forest fragments of western Madagascar. Anim. Conserv. 2003, 6, 147–157. [Google Scholar] [CrossRef]
- Andriatsitohaina, B.; Ramsay, M.S.; Kiene, F.; Lehman, S.M.; Rasoloharijaona, S.; Rakotondravony, R.; Radespiel, U. Ecological fragmentation effects in mouse lemurs and small mammals in northwestern Madagascar. Am. J. Primatol. 2020, 82, e23059. [Google Scholar] [CrossRef]
- Radespiel, U.; Bruford, M.W. Fragmentation genetics of rainforest animals: Insights from recent studies. Conserv. Genet. 2014, 15, 245–260. [Google Scholar] [CrossRef]
- Larsen, P.; Matocq, M. Emerging genomic applications in mammalian ecology, evolution, and conservation. J. Mammal. 2019, 3, 786–801. [Google Scholar] [CrossRef]
- Keyghobadi, N. The genetic implications of habitat fragmentation for animals. Can. J. Zool. 2007, 85, 1049–1064. [Google Scholar] [CrossRef]
- Keller, L.F.; Waller, D.M. Inbreeding effects in wild populations. Trends Ecol. Evol. 2002, 17, 230–241. [Google Scholar] [CrossRef]
- Banks, S.C.; Cary, G.J.; Smith, A.L.; Davies, I.D.; Driscoll, D.A.; Gill, A.M.; Lindenmayer, D.B.; Peakall, R. How does ecological disturbance influence genetic diversity? Trends Ecol. Evol. 2013, 28, 670–679. [Google Scholar] [CrossRef]
- Radespiel, U.; Schulte, J.; Burke, R.J.; Lehman, S.M. Molecular edge effects in the Endangered golden-brown mouse lemur Microcebus ravelobensis. Oryx 2019, 53, 716–726. [Google Scholar] [CrossRef]
- Nathan, R.; Getz, W.M.; Revilla, E.; Holyoak, M.; Kadmon, R.; Saltz, D.; Smouse, P.E. A movement ecology paradigm for unifying organismal movement research. Proc. Natl. Acad. Sci. USA 2008, 105, 19052–19059. [Google Scholar] [CrossRef]
- Galán-Acedo, C.; Arroyo-Rodríguez, V.; Andresen, E.; Verde Arregoitia, L.; Vega, E.; Peres, C.A.; Ewers, R.M. The conservation value of human-modified landscapes for the world’s primates. Nat. Commun. 2019, 10, 152. [Google Scholar] [CrossRef]
- Richardson, J.L.; Michaelides, S.; Combs, M.; Djan, M.; Bisch, L.; Barrett, K.; Silveira, G.; Butler, J.; Aye, T.T.; Munshi-South, J.; et al. Dispersal ability predicts spatial genetic structure in native mammals persisting across an urbanization gradient. Evol. Appl. 2021, 14, 163–177. [Google Scholar] [CrossRef]
- Carleton, M.D.; Goodman, S.M. A new species of the Eliurus majori complex (Rodentia: Muroidea: Nesomyidae) from south-central Madagascar, with remarks on emergent species groupings in the genus Eliurus. Am. Mus. Novitates. 2007, 3547, 1–21. [Google Scholar] [CrossRef]
- Shi, J.; Chan, L.; Rakotomalala, Z.; Heilman, A.; Goodman, S.; Yoder, A. Latitude drives diversification in Madagascar’s endemic dry forest rodent Eliurus myoxinus (subfamily Nesomyinae). Biol. J. Linn. Soc. 2013, 110, 500–517. [Google Scholar] [CrossRef]
- Ramanamanjato, J.B.; Ganzhorn, J.U. Effects of forest fragmentation, introduced Rattus rattus and the role of exotic tree plantations and secondary vegetation for the conservation of an endemic rodent and a small lemur in littoral forests of southeastern Madagascar. Anim. Conserv. 2001, 4, 175–183. [Google Scholar] [CrossRef]
- Sommer, S.; Volahy, A.T.; Seal, U.S. A population and habitat viability assessment for the highly endangered giant jumping rat (Hypogeomys antimena), the largest extant endemic rodent of Madagascar. Anim. Conserv. 2002, 5, 263–273. [Google Scholar] [CrossRef]
- Randrianjafy, V.; Ramilijaona, O.; Rakotondravony, D. Growth of the tuft-tailed rat. Integr. Zool. 2007, 2, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Rakotoarisoa, J.E.; Bailey, C.A.; Hinger, P.H.; Brenneman, R.A.; Louis, E.E. Isolation and characterization of nine microsatellite loci in a Malagasy endemic rodent, Eliurus carletoni (Rodentia: Nesomyinae). Cons. Gen. Resources. 2013, 5, 203–205. [Google Scholar] [CrossRef]
- Rakotoarisoa, J.E.; Raheriarisena, M.; Goodman, S.M. A phylogeographic study of the endemic rodent Eliurus carletoni (Rodentia: Nesomyinae) in an ecological transition zone of northern Madagascar. J. Hered. 2013, 104, 23–35. [Google Scholar] [CrossRef]
- Sgarlata, G.M.; Salmona, J.; Aleixo-Pais, I.; Rakotonanahary, A.; Sousa, A.P.; Kun-Rodrigues, C.; Ralantoharijaona, T.; Jan, F.; Zaranaina, R.; Rasolondraibe, E.; et al. Genetic differentiation and demographic history of the northern rufous mouse lemur (Microcebus tavaratra) across a fragmented landscape in northern Madagascar. Int. J. Prim. 2018, 39, 65–89. [Google Scholar] [CrossRef]
- Shiels, A.B.; Pitt, W.C.; Sugihara, R.T.; Witmer, G.W. Biology and Impacts of Pacific Island Invasive Species. 11. Rattus rattus, the Black Rat (Rodentia: Muridae). Pac. Sci. 2014, 68, 145–184. [Google Scholar] [CrossRef]
- Rahelinirina, S.; Duplantier, J.M.; Ratovonjato, J.; Ramilijaona, O.; Ratsimba, M.; Rahalison, L. Study on the movement of Rattus rattus and evaluation of the plague dispersion in Madagascar. Vector Borne Zoon Dis. 2010, 10, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Goodman, S.M.; Patterson, B.D. Natural Change and Human Impact in Madagascar; Smithsonian Institution Press: Washington, DC, USA, 1997. [Google Scholar]
- Brouat, C.; Tollenaere, C.; Estoup, A.; Loiseau, A.; Sommer, S.; Soanandrasana, R.; Rahalison, L.; Rajerison, M.; Piry, S.; Goodman, S.M.; et al. Invasion genetics of a human commensal rodent: The black rat Rattus rattus in Madagascar. Mol. Ecol. 2014, 23, 4153–4167. [Google Scholar] [CrossRef] [PubMed]
- Gilabert, A.; Loiseau, A.; Duplantier, J.M.; Rahelinirina, S.; Rahalison, L.; Chanteau, S.; Brouat, C. Genetic structure of black rat populations in a rural plague focus in Madagascar. Can. J. Zool. 2007, 85, 965–972. [Google Scholar] [CrossRef]
- Gautier, L.; Tahinarivony, A.J.; Ranirison, P.; Wohlhauser, S. Vegetation. In The Terrestrial Protected Areas of Madagascar: Their History, Description, and Biota; Goodman, S.M., Raherilalao, M.J., Wohlhauser, S., Eds.; Association Vahatra: Antananarivo, Madagascar, 2018; pp. 207–242. [Google Scholar]
- Ramilison, M.L.; Andriatsitohaina, B.; Chell, C.; Rakotondravony, R.; Radespiel, U.; Ramsay, M.S. Distribution of the critically endangered Coquerel’s sifaka (Propithecus coquereli) across a fragmented landscape in NW Madagascar. Afr. J. Ecol. 2021, 59, 350–358. [Google Scholar] [CrossRef]
- Kiene, F.; Andriatsitohaina, B.; Ramsay, M.S.; Rakotondravony, R.; Strube Radespiel, U. Habitat fragmentation and vegetation structure impact gastrointestinal parasites of small mammalian hosts in Madagascar. Ecol. Evol. 2021, 11, 6766–6788. [Google Scholar] [CrossRef]
- Kiene, F.; Andriatsitohaina, B.; Ramsay, M.S.; Rakotondramanana, H.; Rakotondravony, R.; Radespiel, U.; Strube, C. Forest edges affect ectoparasite infestation patterns of small mammalian hosts in fragmented forests in Madagascar. Int. J. Parasitol. 2020, 50, 299–313. [Google Scholar] [CrossRef]
- Radespiel, U.; Rakotondravony, R.; Chikhi, L. Natural and anthropogenic determinants of genetic structure in the largest remaining population of the endangered golden-brown mouse lemur, Microcebus ravelobensis. Am. J. Primatol. 2008, 70, 860–870. [Google Scholar] [CrossRef]
- Davey, J.W.; Blaxter, M.L. RADSeq: Next-generation population genetics. Brief. Funct. Genom. 2010, 9, 416–423. [Google Scholar] [CrossRef]
- Andrews, K.R.; Good, J.M.; Miller, M.R.; Luikart, G.; Hohenlohe, P.A. Harnessing the power of RADseq for ecological and evolutionary genomics. Nature Rev. Gen. 2016, 17, 81–92. [Google Scholar] [CrossRef]
- Catchen, J.; Hohenlohe, P.A.; Bassham, S.; Amores, A.; Cresko, W.A. Stacks: An analysis tool set for population genomics. Mol. Ecol. 2013, 22, 3124–3140. [Google Scholar] [CrossRef]
- Rochette, N.C.; Catchen, J.M. Deriving genotypes from RAD-seq short-read data using Stacks. Nat. Protoc. 2017, 12, 2640–2659. [Google Scholar] [CrossRef]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Alexander, D.H.; Lange, K. Enhancements to the ADMIXTURE algorithm for individual ancestry estimation. BMC Bioinform. 2011, 12, 246. [Google Scholar] [CrossRef] [PubMed]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. Clumpak: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Res. 2015, 15, 1179–1191. [Google Scholar] [CrossRef]
- Greenwood, P.J. Mating systems, philopatry and dispersal in birds and mammals. Anim. Behav. 1980, 28, 1140–1162. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Combs, M.; Puckett, E.E.; Richardson, J.; Mims, D.; Munshi-South, J. Spatial population genomics of the brown rat (Rattus norvegicus) in New York City. Mol. Ecol. 2018, 27, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Petkova, D.; Novembre, J.; Stephens, M. Visualizing spatial population structure with estimated effective migration surfaces. Nat. Genet. 2016, 48, 94–100. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E. Vegan: Community Ecology Package. R Package Version. 2.6-4. CRAN. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 10 January 2022).
- Lack, J.B.; Hamilton, M.J.; Braun, J.K.; Mares, M.A.; Bussche, R.A.V.D. Comparative phylogeography of invasive Rattus rattus and Rattus norvegicus in the U.S. reveals distinct colonization histories and dispersal. Biol. Invasions 2013, 15, 1067–1087. [Google Scholar] [CrossRef]
- Wagner, H.H.; Holderegger, R.; Werth, S.; Gugerli, F.; Hoebee, S.E.; Scheidegger, C. Variogram analysis of the spatial genetic structure of continuous populations using multilocus microsatellite data. Genetics 2005, 169, 1739–1752. [Google Scholar] [CrossRef]
- Barratt, C.D.; Bwong, B.A.; Jehle, R.; Liedtke, H.C.; Nagel, P.; Onstein, R.E.; Portik, D.M.; Streicher, J.W.; Loader, S.P. Vanishing refuge? Testing the forest refuge hypothesis in coastal East Africa using genome-wide sequence data for seven amphibians. Mol. Ecol. 2018, 27, 4289–4308. [Google Scholar] [CrossRef]
- Tang, Q.; Fung, T.; Rheindt, F.E. ResDisMapper: An R package for fine-scale mapping of resistance to dispersal. Mol. Ecol. Res. 2020, 20, 819–831. [Google Scholar] [CrossRef]
- Wood, S.N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J. R. Stat. Soc. Series B 2011, 73, 3–36. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Bartoń, K. MuMIn: Multi-Model Inference. 2018. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 10 January 2021).
- Burnham, K.; Anderson, D. Model selection and multimodel inference. In A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Jacoby, W.G. Loess: A nonparametric, graphical tool for depicting relationships between variables. Elect. Stud. 2000, 19, 577–613. [Google Scholar] [CrossRef]
- Richardson, J.L.; Burak, M.K.; Hernandez, C.; Shirvell, J.M.; Mariani, C.; Carvalho-Pereira, T.S.A.; Pertile, A.C.; Panti-May, J.A.; Pedra, G.G.; Serrano, S.; et al. Using fine-scale spatial genetics of Norway rats to improve control efforts and reduce leptospirosis risk in urban slum environments. Evol. Appl. 2017, 10, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.R.; MacDonald, R.L.; Chen, H.Y.H. Disturbance increases negative spatial autocorrelation in species diversity. Landscape Ecol. 2017, 32, 823–834. [Google Scholar] [CrossRef]
- McLoughlin, P.D.; Morris, D.W.; Fortin, D.; Vander Wal, E.; Contasti, A.L. Considering ecological dynamics in resource selection functions. J. Anim. Ecol. 2010, 79, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.J.; Goodman, S.M.; Farris, Z.J.; Karpanty, S.M.; Andrianjakarivelo, V.; Kelly, M.J. Landscape trends in small mammal occupancy in the Makira–Masoala protected areas, northeastern Madagascar. J. Mammal. 2017, 98, 272–282. [Google Scholar] [CrossRef]
- Narum, S.R.; Buerkle, C.A.; Davey, J.W.; Miller, M.R.; Hohenlohe, P.A. Genotyping-by-sequencing in ecological and conservation genomics. Mol. Ecol. 2013, 22, 2841–2847. [Google Scholar] [CrossRef]
- Hingston, M.; Goodman, S.M.; Ganzhorn, J.U.; Sommer, S. Reconstruction of the colonization of southern Madagascar by introduced Rattus rattus. J. Biogeogr. 2005, 32, 1549–1559. [Google Scholar] [CrossRef]
- Allendorf, F.W.; Lundquist, L.L. Introduction: Population Biology, Evolution, and Control of Invasive Species. Cons Biol. 2003, 17, 24–30. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Data Set | Categorical Variables | Continuous Variables |
|---|---|---|---|
| A | All individuals | Species, Region, Sex | Body mass, Forest Area |
| B | All individuals | Species, Region, Sex, Forest Type | Body mass, Distance to Edge |
| C | Only individuals from fragments | Species, Region, Sex | Body mass, Forest Area |
| D | Only individuals from fragments | Species, Region, Sex | Body mass, Distance to Edge |
| E | Only individuals from fragments | Species, Region, Sex | Body mass, Distance to Continuous Forest, Percentage of Edge |
| Species | Region | N FOR | Distance (m) | Range (m) | N SOR | Distance (m) | Range (m) | N DR | Distance (m) | Range (m) |
|---|---|---|---|---|---|---|---|---|---|---|
| E. myoxinus | ANP | 1 | 2174 | 2174 | 17 | 35 | 0–294 | 8 | 770 | 494–2523 |
| MCF | 0 | NA | NA | 0 | NA | NA | 0 | NA | NA | |
| R. rattus | ANP | 3 | 6 | 0–10 | 2 | 5 | 0–11 | 5 | 263 | 93–758 |
| MCF | 0 | NA | NA | 3 | 62 | 49–110 | 3 | 75 | 40–520 |
| Data Set | Model | Species | Region | Sex | Forest Type | Body Mass | Forest Area | Distance to Edge | Distanceto CF | Edge Percentage | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| FIS | All | A | Em > Rr | MCF > ANP | |||||||
| All | B | Em > Rr | MCF > ANP | + | |||||||
| Fragments | C | Em > Rr | MCF > ANP | ||||||||
| Fragments | D | Em > Rr | MCF > ANP | ns | |||||||
| Fragments | E | Em > Rr | MCF > ANP | ns | |||||||
| HO | All | A | Rr > Em | ||||||||
| All | B | Rr > Em | |||||||||
| Fragments | C | Rr > Em | ns | ||||||||
| Fragments | D | Rr > Em | |||||||||
| Fragments | E | Rr > Em | ns | ||||||||
| HE | All | A | Rr > Em | MCF > ANP | ns | ||||||
| All | B | Rr > Em | MCF > ANP | ns | ns | ||||||
| Fragments | C | Rr > Em | MCF > ANP | ||||||||
| Fragments | D | Rr > Em | MCF > ANP | ||||||||
| Fragments | E | Rr > Em | MCF > ANP |
| Species | Data Set | Model | Region | Sex | Forest Type | Body Mass | Forest Area | Distance to Edge | Distance to CF | Edge Percentage | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| FIS | Em | All | A | MCF > ANP | |||||||
| All | B | MCF > ANP | ns | ||||||||
| Fragments | C | MCF > ANP | ns | ||||||||
| Fragments | D | ns | |||||||||
| Fragments | E | ns | ns | ||||||||
| HO | Em | All | A | MCF > ANP | |||||||
| All | B | MCF > ANP | - | ||||||||
| Fragments | C | MCF > ANP | ns | ||||||||
| Fragments | D | MCF > ANP | |||||||||
| Fragments | E | MCF > ANP | ns | ||||||||
| HO | Em | All | A | MCF > ANP | |||||||
| All | B | MCF > ANP | ns | ||||||||
| Fragments | C | MCF > ANP | ns | ||||||||
| Fragments | D | MCF > ANP | |||||||||
| Fragments | E | ns | |||||||||
| FIS | Rr | All | A | MCF > ANP | |||||||
| All | B | MCF > ANP | |||||||||
| Fragments | C | ns | |||||||||
| Fragments | D | ns | |||||||||
| Fragments | E | ns | |||||||||
| HO | Rr | All | A | ANP > MCF | |||||||
| All | B | ANP > MCF | |||||||||
| Fragments | C | ||||||||||
| Fragments | D | ||||||||||
| Fragments | E | ||||||||||
| HE | Rr | All | A | ANP > MCF | |||||||
| All | B | ANP > MCF | |||||||||
| Fragments | C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramsay, M.S.; Sgarlata, G.M.; Barratt, C.D.; Salmona, J.; Andriatsitohaina, B.; Kiene, F.; Manzi, S.; Ramilison, M.L.; Rakotondravony, R.; Chikhi, L.; et al. Effects of Forest Fragmentation on Connectivity and Genetic Diversity in an Endemic and an Invasive Rodent in Northwestern Madagascar. Genes 2023, 14, 1451. https://doi.org/10.3390/genes14071451
Ramsay MS, Sgarlata GM, Barratt CD, Salmona J, Andriatsitohaina B, Kiene F, Manzi S, Ramilison ML, Rakotondravony R, Chikhi L, et al. Effects of Forest Fragmentation on Connectivity and Genetic Diversity in an Endemic and an Invasive Rodent in Northwestern Madagascar. Genes. 2023; 14(7):1451. https://doi.org/10.3390/genes14071451
Chicago/Turabian StyleRamsay, Malcolm S., Gabriele M. Sgarlata, Christopher D. Barratt, Jordi Salmona, Bertrand Andriatsitohaina, Frederik Kiene, Sophie Manzi, Miarisoa L. Ramilison, Romule Rakotondravony, Lounès Chikhi, and et al. 2023. "Effects of Forest Fragmentation on Connectivity and Genetic Diversity in an Endemic and an Invasive Rodent in Northwestern Madagascar" Genes 14, no. 7: 1451. https://doi.org/10.3390/genes14071451
APA StyleRamsay, M. S., Sgarlata, G. M., Barratt, C. D., Salmona, J., Andriatsitohaina, B., Kiene, F., Manzi, S., Ramilison, M. L., Rakotondravony, R., Chikhi, L., Lehman, S. M., & Radespiel, U. (2023). Effects of Forest Fragmentation on Connectivity and Genetic Diversity in an Endemic and an Invasive Rodent in Northwestern Madagascar. Genes, 14(7), 1451. https://doi.org/10.3390/genes14071451