
Genotypic and Haplotypic Association of Catechol-O-Methyltransferase rs4680 and rs4818 Gene Polymorphisms with Particular Clinical Symptoms in Schizophrenia

 , , , , , and
, , , , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Genotyping
2.3. Statistical Analyses
3. Results
Demographic Data
4. Discussion
Limitations and Strengths
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bradley, A.J.; Dinan, T.G. A systematic review of hypothalamic-pituitary-adrenal axis function in schizophrenia: Implications for mortality. J. Psychopharmacol. 2010, 24 (Suppl. S4), 91–118. [Google Scholar] [CrossRef]
- Mizuno, Y.; Hofer, A.; Suzuki, T.; Frajo-Apor, B.; Wartelsteiner, F.; Kemmler, G.; Saruta, J.; Tsukinoki, K.; Mimura, M.; Fleischhacker, W.W.; et al. Clinical and biological correlates of resilience in patients with schizophrenia and bipolar disorder: A cross-sectional study. Schizophr. Res. 2016, 175, 148–153. [Google Scholar] [CrossRef]
- Chuang, S.P.; Wu, J.Y.W.; Wang, C.S. Resilience and Quality of Life in People with Mental Illness: A Systematic Review and Meta-Analysis. Neuropsychiatr. Dis. Treat. 2023, 4, 507–514. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar]
- Alkan, E.; Davies, G.; Evans, S.L. Cognitive impairment in schizophrenia: Relationships with cortical thickness in fronto-temporal regions, and dissociability from symptom severity. NPJ Schizophr. 2021, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Harvey, P.D.; Bosia, M.; Cavallaro, R.; Howes, O.D.; Kahn, R.S.; Leucht, S.; Müller, D.R.; Penadés, R.; Vita, A. Cognitive dysfunction in schizophrenia: An expert group paper on the current state of the art. Schizophr. Res. Cogn. 2022, 29, 100249. [Google Scholar] [CrossRef] [PubMed]
- Rekhi, G.; Saw, Y.E.; Lim, K.; Keefe, R.S.E.; Lee, J. Impact of Cognitive Impairments on Health-Related Quality of Life in Schizophrenia. Brain Sci. 2023, 13, 215. [Google Scholar] [CrossRef]
- Etkin, A.; Gyurak, A.; O’Hara, R. A neurobiological approach to the cognitive deficits of psychiatric disorders. Dialogues Clin. Neurosci. 2013, 15, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Li, S.C. Neuromodulation and developmental contextual influences on neural and cognitive plasticity across the lifespan. Neurosci. Biobehav. Rev. 2013, 37 Pt B, 2201–2208. [Google Scholar] [CrossRef]
- Goldberg, T.E.; Weinberger, D.R. Genes and the parsing of cognitive processes. Trends Cogn. Sci. 2004, 8, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Nikolac Perkovic, M.; Nedic Erjavec, G.; Svob Strac, D.; Uzun, S.; Kozumplik, O.; Pivac, N. Theranostic biomarkers for schizophrenia. Int. J. Mol. Sci. 2017, 18, 733. [Google Scholar] [CrossRef]
- Brisch, R.; Saniotis, A.; Wolf, R.; Bielau, H.; Bernstein, H.G.; Steiner, J.; Bogerts, B.; Braun, K.; Jankowski, Z.; Kumaratilake, J.; et al. The role of dopamine in schizophrenia from a neurobiological and evolutionary perspective: Old fashioned, but still in vogue. Front. Psychiatry 2014, 19, 47. [Google Scholar]
- Westbrook, A.; Braver, T.S. Dopamine does double duty in motivating cognitive effort. Neuron 2016, 89, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Jatana, N.; Sharma, A.; Latha, N. Pharmacophore modeling and virtual screening studies to design potential COMT inhibitors as new leads. J. Mol. Graph. Model. 2013, 39, 145–164. [Google Scholar] [CrossRef] [PubMed]
- Zubieta, C.; Kota, P.; Ferrer, J.L.; Dixon, R.A.; Noel, J.P. Structural basis for the modulation of lignin monomer methylation by caffeic acid/5-hydroxyferulic acid 3/5-O-methyltransferase. Plant Cell 2002, 14, 1265–1277. [Google Scholar] [CrossRef]
- Craddock, N.; Owen, M.J.; O’Donovan, M.C. The catechol-O-methyl transferase (COMT) gene as a candidate for psychiatric phenotypes: Evidence and lessons. Mol. Psychiatry 2006, 11, 446–458. [Google Scholar] [CrossRef]
- Tunbridge, E.M.; Narajos, M.; Harrison, C.H.; Beresford, C.; Cipriani, A.; Harrison, P.J. Which dopamine polymorphisms are functional? Systematic review and meta-analysis of COMT, DAT, DBH, DDC, DRD1-5, MAOA, MAOB, TH, VMAT1, and VMAT2. Biol. Psychiatry 2019, 86, 608–620. [Google Scholar] [CrossRef]
- Lachman, H.M.; Papolos, D.F.; Saito, T.; Yu, Y.M.; Szumlanski, C.L.; Weinshilboum, R.M. Human catechol-O-methyltransferase pharmacogenetics: Description of a functional polymorphism and its potential application to neuropsychiatric disorders. Pharmacogenetics 1996, 6, 243–250. [Google Scholar] [CrossRef]
- Roussos, P.; Giakoumaki, S.G.; Pavlakis, S.; Bitsios, P. Planning, decision-making and the COMT rs4818 polymorphism in healthy males. Neuropsychologia 2008, 46, 757–763. [Google Scholar] [CrossRef]
- Nackley, A.G.; Shabalina, S.A.; Tchivileva, I.E.; Satterfield, K.; Korchynskyi, O.; Makarov, S.S.; Maixner, W.; Diatchenko, L. Human catechol-O-methyltransferase haplotypes modulate protein expression by altering mRNA secondary structure. Science 2006, 314, 1930–1933. [Google Scholar] [CrossRef]
- González-Castro, T.B.; Hernández-Díaz, Y.; Juárez-Rojop, I.E.; López-Narváez, M.L.; Tovilla-Zárate, C.A.; Fresan, A. The role of a Catechol-O-Methyltransferase (COMT) Val158Met genetic polymorphism in schizophrenia: A Systematic review and updated meta-analysis on 32,816 subjects. Neuromolecular. Med. 2016, 18, 216–231. [Google Scholar] [CrossRef]
- Gong, J.; Zhang, T.; Zhou, L.; Mo, Y.; Yu, F.; Liu, M.; Yang, L.; Liu, J. Gender divergent effect of COMT gene rs4680 polymorphism on an association between executive dysfunction and psychotic-like experiences. Behav. Brain Res. 2023, 439, 114215. [Google Scholar] [CrossRef] [PubMed]
- Shifman, S.; Bronstein, M.; Sternfeld, M.; Pisanté-Shalom, A.; Lev-Lehman, E.; Weizman, A.; Reznik, I.; Spivak, B.; Grisaru, N.; Karp, L.; et al. A highly significant association between the COMT haplotype and schizophrenia. Am. J. Hum. Genet. 2002, 71, 1296–1302. [Google Scholar] [CrossRef] [PubMed]
- Dennison, C.A.; Legge, S.E.; Pardiñas, A.F.; Walters, J.T.R. Genome-wide association studies in schizophrenia: Recent advances, challenges and future perspective. Schizophr. Res. 2020, 217, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Nikolac, M.; Sagud, M.; Nedic, G.; Nenadic Sviglin, K.; Mihaljevic Peles, A.; Uzun, S.; Vuskan Cusa, B.; Kozumplik, O.; Zivkovic, M.; Mustapic, M.; et al. The lack of association between catechol-O-methyl-transferase Val108/158Met polymorphism and smoking in schizophrenia and alcohol dependence. Psychiatry Res. 2013, 205, 179–180. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Lu, R.B.; Yeh, Y.W.; Shih, M.C.; Huang, S.Y. Association study of catechol-O-methyltransferase gene polymorphisms with schizophrenia and psychopathological symptoms in Han Chinese. Genes Brain Behav. 2011, 10, 316–324. [Google Scholar] [CrossRef]
- Li, W.J.; Kou, C.G.; Yu, Y.; Sun, S.; Zhang, X.; Kosten, T.R.; Zhang, X.Y. Association of catechol-O-methyltransferase gene polymorphisms with schizophrenia and negative symptoms in a Chinese population. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2012, 159, 370–375. [Google Scholar] [CrossRef]
- Madzarac, Z.; Tudor, L.; Sagud, M.; Nedic Erjavec, G.; Mihaljevic Peles, A.; Pivac, N. The Associations between COMT and MAO-B genetic variants with negative symptoms in patients with schizophrenia. Curr. Issues Mol. Biol. 2021, 43, 618–636. [Google Scholar] [CrossRef]
- Sagud, M.; Tudor, L.; Uzun, S.; Nikolac Perkovic, M.; Zivkovic, M.; Konjevod, M.; Kozumplik, O.; Vuksan Cusa, B.; Svob Strac, D.; Rados, I.; et al. Haplotypic and genotypic association of Catechol-O-Methyltransferase rs4680 and rs4818 polymorphisms and treatment resistance in schizophrenia. Front. Pharmacol. 2018, 9, 705. [Google Scholar] [CrossRef]
- Nikolac Perkovic, M.; Sagud, M.; Zivkovic, M.; Uzun, S.; Nedic Erjavec, G.; Kozumplik, O.; Svob Strac, D.; Mimica, N.; Mihaljevic Peles, A.; Pivac, N. Catechol-O-methyltransferase rs4680 and rs4818 haplotype association with treatment response to olanzapine in patients with schizophrenia. Sci. Rep. 2020, 10, 10049. [Google Scholar] [CrossRef]
- Tunbridge, E.M.; Harrison, P.J. Importance of the COMT gene for sex differences in brain function and predisposition to psychiatric disorders. Curr. Top. Behav. Neurosci. 2011, 8, 119–140. [Google Scholar]
- Kay, S.R.; Fiszbein, A.; Opler, L.A. The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophr. Bull. 1987, 13, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Lindström, E.; Tuninger, E.; Levander, S. PECC-factor structure and findings in three longitudinal cohorts of patients with schizophrenia. Nord. J. Psychiatry 2012, 66, 33–39. [Google Scholar] [CrossRef]
- Bagney, A.; Dompablo, M.; Santabárbara, J.; Moreno-Ortega, M.; Lobo, A.; Jimenez-Arriero, M.A.; Palomo, T.; Rodriguez-Jimenez, R. Are negative symptoms really related to cognition in schizophrenia? Psychiatry Res. 2015, 230, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.; Peh, O.H.; Yang, Z.; Rekhi, G.; Rapisarda, A.; See, Y.M.; Rashid, N.A.A.; Ang, M.S.; Lee, S.A.; Sim, K.; et al. Large-scale evaluation of the Positive and Negative Syndrome Scale (PANSS) symptom architecture in schizophrenia. Asian J. Psychiatr. 2021, 62, 102732. [Google Scholar] [CrossRef]
- First, M.B.; Spitzer, R.L.; Williams, J.B.W.; Gibbons, M. Structured Clinical Interview for DSM-IV Patient Edition (SCID-P); American Psychiatric Press: Washington, DC, USA, 1995. [Google Scholar]
- Montoya, A.; Valladares, A.; Lizán, L.; San, L.; Escobar, R.; Paz, S. Validation of the Excited Component of the Positive and Negative Syndrome Scale (PANSS-EC) in a naturalistic sample of 278 patients with acute psychosis and agitation in a psychiatric emergency room. Health Qual. Life Outcomes 2011, 9, 18. [Google Scholar] [CrossRef]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A Simple Salting out Procedure for Extracting DNA from Human Nucleated Cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef]
- Unwin, A. Discovering Statistics Using R by Andy Field, Jeremy Miles, Zoë Field. Int. Stat. Rev. 2013, 81, 169–170. [Google Scholar] [CrossRef]
- Rodriguez, S.; Gaunt, T.R.; Day, I.N. Hardy-Weinberg equilibrium testing of biological ascertainment for Mendelian randomization studies. Am. J. Epidemiol. 2009, 169, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and Visualization of LD and Haplotype Maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Shafer, A.; Dazzi, F. Meta-analysis of the positive and Negative Syndrome Scale (PANSS) factor structure. J. Psychiatr. Res. 2019, 115, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Mihaljević-Peleš, A.; Bajs Janović, M.; Šagud, M.; Živković, M.; Janović, Š.; Jevtović, S. Cognitive deficit in schizophrenia: An overview. Psychiatr. Danub. 2019, 31 (Suppl. S2), 139–142. [Google Scholar]
- Rosen, C.; Harrow, M.; Tong, L.; Jobe, T.H.; Harrow, H. A word is worth a thousand pictures: A 20-year comparative analysis of aberrant abstraction in schizophrenia, affective psychosis, and non-psychotic depression. Schizophr. Res. 2021, 238, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Suetani, S.; Baker, A.; Garner, K.; Cosgrove, P.; Mackay-Sim, M.; Siskind, D.; Murray, G.K.; Scott, J.G.; Kesby, J.P. Impairments in goal-directed action and reversal learning in a proportion of individuals with psychosis. Cogn. Affect. Behav. Neurosci. 2022, 22, 1390–1403. [Google Scholar] [CrossRef]
- Ricarte, J.J.; Del Rey, F.; Ros, L.; Latorre, J.M.; Berna, F. Abstract and experiential thinking differentially account for anomalous perception of reality in people with or without schizophrenia. Schizophr. Res. 2018, 193, 43–50. [Google Scholar] [CrossRef]
- Vignapiano, A.; Koenig, T.; Mucci, A.; Giordano, G.M.; Amodio, A.; Altamura, M.; Bellomo, A.; Brugnoli, R.; Corrivetti, G.; Di Lorenzo, G.; et al. Italian Network for Research on Psychoses. Disorganization and cognitive impairment in schizophrenia: New insights from electrophysiological findings. Int. J. Psychophysiol. 2019, 145, 99–108. [Google Scholar] [CrossRef]
- Ortiz, B.B.; Higuchi, C.H.; Noto, C.; Joyce, D.W.; Correll, C.U.; Bressan, R.A.; Gadelha, A. A symptom combination predicting treatment-resistant schizophrenia—A strategy for real-world clinical practice. Schizophr. Res. 2020, 218, 195–200. [Google Scholar] [CrossRef]
- Farreny, A.; Usall, J.; Cuevas-Esteban, J.; Ochoa, S.; Brébion, G. Amendment of traditional assessment measures for the negative symptoms of schizophrenia. Eur. Psychiatry 2018, 49, 50–55. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, Y.; Lu, Z.; Yan, H.; Guo, L.; Liao, Y.; Lu, T.; Wang, L.; Li, J.; Li, W.; et al. Longitudinal network analysis reveals interactive change of schizophrenia symptoms during acute antipsychotic treatment. Schizophr. Bull. 2023, 49, 208–217. [Google Scholar] [CrossRef]
- Cheng, Z.; Yuan, Y.; Han, X.; Yang, L.; Zeng, X.; Yang, F.; Lu, Z.; Wang, C.; Deng, H.; Zhao, J.; et al. Which subgroup of first-episode schizophrenia patients can remit during the first year of antipsychotic treatment? Front. Psychiatry 2020, 11, 566. [Google Scholar] [CrossRef] [PubMed]
- Hoprekstad, G.E.; Kjelby, E.; Gjestad, R.; Fathian, F.; Larsen, T.K.; Reitan, S.K.; Rettenbacher, M.; Torsvik, A.; Skrede, S.; Johnsen, E.; et al. Depression trajectories and cytokines in schizophrenia spectrum disorders—A longitudinal observational study. Schizophr. Res. 2023, 252, 77–87. [Google Scholar] [CrossRef]
- Morozova, A.; Zorkina, Y.; Pavlov, K.; Pavlova, O.; Storozheva, Z.; Zubkov, E.; Zakharova, N.; Karpenko, O.; Reznik, A.; Chekhonin, V.; et al. Association of rs4680 COMT, rs6280 DRD3, and rs7322347 5HT2A With Clinical Features of Youth-Onset Schizophrenia. Front. Psychiatry 2019, 10, 830. [Google Scholar] [CrossRef] [PubMed]
- Rajji, T.K.; Ismail, Z.; Mulsant, B.H. Age at onset and cognition in schizophrenia: Meta-analysis. Br. J. Psychiatry 2009, 195, 286–293. [Google Scholar] [CrossRef]
- Twamley, E.W.; Hua, J.P.; Burton, C.Z.; Vella, L.; Chinh, K.; Bilder, R.M.; Kelsoe, J.R. Effects of COMT genotype on cognitive ability and functional capacity in individuals with schizophrenia. Schizophr. Res. 2014, 159, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T.; Lu, W.; Wan, L.; Yan, H.; Wang, C.; Yang, F.; Tan, Y.; Li, L.; Chinese Schizophrenia Collaboration Group; Yu, H.; et al. Individual differences in schizophrenia. BJPsych Open 2017, 3, 265–273. [Google Scholar] [CrossRef]
- Parkin, G.M.; Udawela, M.; Gibbons, A.; Scarr, E.; Dean, B. Catechol-O-methyltransferase (COMT) genotypes are associated with varying soluble, but not membrane-bound COMT protein in the human prefrontal cortex. J. Hum. Genet. 2018, 63, 1251–1258. [Google Scholar] [CrossRef]
- Oh, J.; Chun, J.W.; Joon, J.H.; Kim, E.; Park, H.J.; Lee, B.; Kim, J.J. The neural basis of a deficit in abstract thinking in patients with schizophrenia. Psychiatry Res. 2015, 234, 66–73. [Google Scholar] [CrossRef]
- Martens, M.; McConnell, F.K.; Filippini, N.; Mackay, C.E.; Harrison, P.J.; Tunbridge, E.M. Dopaminergic modulation of regional cerebral blood flow: An arterial spin labelling study of genetic and pharmacological manipulation of COMT activity. Neuroimage 2021, 234, 117999. [Google Scholar] [CrossRef]
- Marder, S.R.; Davis, J.M.; Chouinard, G. The effects of risperidone on the five dimensions of schizophrenia derived by factor analysis: Combined results of the North American trials. J. Clin. Psychiatry 1997, 58, 538–546. [Google Scholar] [CrossRef]
- Peitl, V.; Štefanović, M.; Karlović, D. Depressive symptoms in schizophrenia and dopamine and serotonin gene polymorphisms. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 77, 209–215. [Google Scholar] [CrossRef]
- Tosato, S.; Bonetto, C.; de Santi, K.; Lasalvia, A.; Gennarelli, M.; Cristofalo, D.; Bertani, M.; Ruggeri, M. on behalf of the Picos-Veneto group. COMT but Not 5HTTLPR gene is associated with depression in first-episode psychosis: The role of stressful life events. Genes 2023, 14, 350. [Google Scholar] [CrossRef] [PubMed]
- Nour El Huda, A.R.; Norsidah, K.Z.; Nabil Fikri, M.R.; Hanisah, M.N.; Kartini, A.; Norlelawati, A.T. DNA methylation of membrane-bound catechol-O-methyltransferase in Malaysian schizophrenia patients. Psychiatry Clin. Neurosci. 2018, 72, 266–279. [Google Scholar] [CrossRef]
- Pirio Richardson, S.; Triggiani, A.I.; Matsuhashi, M.; Voon, V.; Peckham, E.; Nahab, F.; Mari, Z.; Hallett, M. Timing of the sense of volition in patients with schizophrenia. Front. Neurosci. 2020, 14, 574472. [Google Scholar] [CrossRef]
- Cesková, E.; Prikryl, R.; Kaspárek, T.; Ondrusová, M. Psychopathology and treatment responsiveness of patients with first-episode schizophrenia. Neuropsychiatr. Dis. Treat. 2005, 1, 179–185. [Google Scholar] [CrossRef]
- Mellem, M.S.; Kollada, M.; Tiller, J.; Lauritzen, T. Explainable AI enables clinical trial patient selection to retrospectively improve treatment effects in schizophrenia. BMC Med. Inform. Decis. Mak. 2021, 21, 162. [Google Scholar] [CrossRef]
- Ogyu, K.; Noda, Y.; Yoshida, K.; Kurose, S.; Masuda, F.; Mimura, Y.; Nishida, H.; Plitman, E.; Tarumi, R.; Tsugawa, S.; et al. Early improvements of individual symptoms as a predictor of treatment response to asenapine in patients with schizophrenia. Neuropsychopharmacol. Rep. 2020, 40, 138–149. [Google Scholar] [CrossRef]
- Laatikainen, L.M.; Sharp, T.; Harrison, P.J.; Tunbridge, E.M. Sexually dimorphic effects of catechol-O-methyltransferase (COMT) inhibition on dopamine metabolism in multiple brain regions. PLoS ONE 2013, 8, e61839. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Bai, W.; Wang, W.; Jiang, H.; Jin, B.; Liu, Y.; Liu, S.; Wang, K.; Jia, J.; Qin, L. Mechanisms underlying alterations in norepinephrine levels in the locus coeruleus of ovariectomized rats: Modulation by estradiol valerate and black cohosh. Neuroscience 2017, 354, 110–121. [Google Scholar] [CrossRef]
- Louis, C.C.; Jacobs, E.; D’Esposito, M.; Moser, J. Estradiol and the Catechol-O-methyltransferase Gene Interact to Predict Working Memory Performance: A Replication and Extension. J. Cogn. Neurosci. 2023, 35 (Suppl. S7), 1144–1153. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.K.; Yang, M.C.; Chen, C.S.; Wang, I.C.; Kao, W.T.; Shi, M.D. A gender-specific COMT haplotype contributes to risk modulation rather than disease severity of major depressive disorder in a Chinese population. J. Affect. Disord. 2019, 246, 376–386. [Google Scholar] [CrossRef]
- Sagud, M.; Mück-Seler, D.; Mihaljević-Peles, A.; Vuksan-Cusa, B.; Zivković, M.; Jakovljević, M.; Pivac, N. Catechol-O-methyl transferase and schizophrenia. Psychiat. Danub. 2010, 22, 270–274. [Google Scholar]
- Correll, C.U.; Schooler, N.R. Negative symptoms in schizophrenia: A review and clinical guide for recognition, assessment, and treatment. Neuropsychiatr. Dis. Treat. 2020, 16, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Vidal, P.M.; Pacheco, R. The cross-talk between the dopaminergic and the immune system involved in schizophrenia. Front. Pharmacol. 2020, 11, 394. [Google Scholar] [CrossRef] [PubMed]
- Cumming, P.; Abi-Dargham, A.; Gründer, G. Molecular imaging of schizophrenia: Neurochemical findings in a heterogeneous and evolving disorder. Behav. Brain Res. 2021, 398, 113004. [Google Scholar] [CrossRef] [PubMed]
- Werner, M.C.F.; Wirgenes, K.V.; Haram, M.; Bettella, F.; Lunding, S.H.; Rødevand, L.; Hjell, G.; Agartz, I.; Djurovic, S.; Melle, I.; et al. Indicated association between polygenic risk score and treatment-resistance in a naturalistic sample of patients with schizophrenia spectrum disorders. Schizophr. Res. 2020, 218, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Wimberley, T.; Gasse, C.; Meier, S.M.; Agerbo, E.; MacCabe, J.H.; Horsdal, H.T. Polygenic Risk Score for Schizophrenia and Treatment-Resistant Schizophrenia. Schizophr. Bull. 2017, 43, 1064–1069. [Google Scholar] [CrossRef]
- Zhang, J.P.; Robinson, D.; Yu, J.; Gallego, J.; Fleischhacker, W.W.; Kahn, R.S.; Crespo-Facorro, B.; Vazquez-Bourgon, J.; Kane, J.M.; Malhotra, A.K.; et al. Schizophrenia Polygenic Risk Score as a Predictor of Antipsychotic Efficacy in First-Episode Psychosis. Am. J. Psychiatry 2019, 176, 21–28. [Google Scholar] [CrossRef]
- Richards, A.L.; Pardinas, A.F.; Frizzati, A.; Tansey, K.E.; Lynham, A.J.; Holmans, P.; Legge, S.E.; Savage, J.E.; Agartz, I.; Andreassen, O.A.; et al. The Relationship Between Polygenic Risk Scores and Cognition in Schizophrenia. Schizophr. Bull. 2020, 46, 336–344. [Google Scholar] [CrossRef]
- Habtewold, T.D.; Liemburg, E.J.; Islam, M.A.; de Zwarte, S.M.C.; Boezen, H.M.; GROUP Investigators; Bruggeman, R.; Alizadeh, B.Z. Association of schizophrenia polygenic risk score with data-driven cognitive subtypes: A six-year longitudinal study in patients, siblings and controls. Schizophr. Res. 2020, 223, 135–147. [Google Scholar] [CrossRef]
- Rantala, M.J.; Luoto, S.; Borráz-León, J.I.; Krams, I. Schizophrenia: The new etiological synthesis. Neurosci. Biobehav. Rev. 2022, 142, 104894. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.J.; Tunbridge, E.M. Catechol-O-methyltransferase (COMT): A gene contributing to sex differences in brain function, and to sexual dimorphism in the predisposition to psychiatric disorders. Neuropsychopharmacology 2008, 33, 3037–3045. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Shang, S.; Su, Y. Genetic influences on insight problem solving: The role of catechol-O-methyltransferase (COMT) gene polymorphisms. Front. Psychol. 2015, 6, 1569. [Google Scholar] [CrossRef] [PubMed]
- Piffer, D. Correlation of the COMT Val158Met polymorphism with latitude and hunter-gather lifestyle suggests culture–gene coevolution and selective pressure on cognition genes due to climate. Anthropol. Sci. 2013, 121, 161–171. [Google Scholar] [CrossRef]
- Käenmäki, M.; Tammimäki, A.; Myöhänen, T.; Pakarinen, K.; Amberg, C.; Karayiorgou, M.; Gogos, J.A.; Männistö, P.T. Quantitative role of COMT in dopamine clearance in the prefrontal cortex of freely moving mice. J. Neurochem. 2010, 114, 1745–1755. [Google Scholar] [CrossRef]
- Tunbridge, E.; Burnet, P.W.; Sodhi, M.S.; Harrison, P.J. Catechol-O-methyltransferase (COMT) and proline dehydrogenase (PRODH) mRNAs in the dorsolateral prefrontal cortex in schizophrenia, bipolar disorder, and major depression. Synapse 2004, 51, 112–118. [Google Scholar] [CrossRef]
- Aggarwal, N.K.; Tao, H.; Xu, K.; Stefanovics, E.; Liu, Z.; Rosenheck, R.A. Comparing the PANSS in Chinese and American inpatients: Cross-cultural psychiatric analyses of instrument translation and implementation. Schizophr. Res. 2011, 132, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Bray, N.J.; Buckland, P.R.; Williams, N.M.; Williams, H.J.; Norton, N.; Owen, M.J.; O’Donovan, M.C. A haplotype implicated in schizophrenia susceptibility is associated with reduced COMT expression in human brain. Am. J. Hum. Genet. 2003, 73, 152–161. [Google Scholar] [CrossRef]




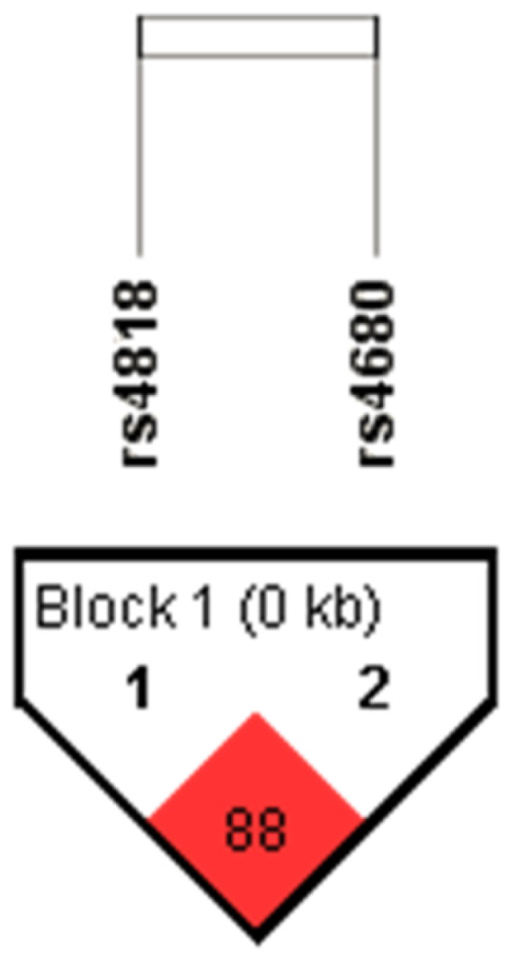{kind=link}
{kind=link}
{kind=link}
| Male Subjects | Female Subjects | Statistics | |
|---|---|---|---|
| Age | 38 (30; 47) | 48 (38; 56) | U = 28,290.0; p < 0.001 |
| Smoking (Yes/No) | 376 (69.1%)/168 (30.9%) | 199 (51.7%)/186 (48.3%) | χ2 = 29.038; p < 0.001 |
| PANSS total | 116 (98; 133) | 93 (78; 104) | U = 50,937.5; p < 0.001 |
| PANSS positive | 30 (24; 36) | 23 (18; 27) | U = 52,917.0; p < 0.001 |
| PANSS negative | 29 (24; 35) | 23 (20; 26) | U = 55,396.5; p < 0.001 |
| PANSS general | 56 (49; 63) | 46 (39; 52) | U = 57,075.0; p < 0.001 |
| PANSS cognitive | 15 (12; 18) | 12 (10; 13) | U = 55,604.0; p < 0.001 |
| SNP | Male Subjects | Female Subjects | Statistics | Smokers | Non-Smokers | Statistics | |
|---|---|---|---|---|---|---|---|
| COMT rs4818 | CC | 204 (37.5%) | 148 (38.4%) | χ2 = 0.560; df = 2; p = 0.756 | 228 (39.7%) | 124 (35.0%) | χ2 = 2.183; df = 2; p = 0.336 |
| CG | 261 (48.0%) | 176 (45.7%) | 265 (46.1%) | 172 (48.6%) | |||
| GG | 79 (14.5%) | 61 (15.8%) | 82 (14.3%) | 58 (16.4%) | |||
| C | 465 (85.5%) | 324 (84.2%) | χ2 = 0.308; df = 1; p = 0.579 | 493 (85.7%) | 296 (83.6%) | χ2 = 0.772; df = 1; p = 0.380 | |
| GG | 79 (14.5%) | 61 (15.8%) | 82 (14.3%) | 58 (16.4%) | |||
| G | 340 (62.5%) | 237 (61.6%) | χ2 = 0.085; df = 1; p = 0.771 | 347 (60.3%) | 230 (65.0%) | χ2 = 1.991; df = 1; p = 0.158 | |
| CC | 204 (37.5%) | 148 (38.4%) | 228 (39.7%) | 124 (35.0%) | |||
| C | 669 (61.5%) | 472 (61.3%) | χ2 = 0.007; df = 1; p = 0.934 | 721 (62.7%) | 420 (59.3%) | χ2 = 2.105; df = 1; p = 0.147 | |
| G | 419 (38.5%) | 298 (38.7%) | 429 (37.3%) | 288 (40.7%) | |||
| COMT rs4680 | AA | 143 (26.3%) | 95 (24.7%) | χ2 = 0.919; df = 2; p = 0.632 | 154 (26.8%) | 84 (23.7%) | χ2 = 1.140; df = 2; p = 0.566 |
| GA | 270 (49.6%) | 187 (48.6%) | 280 (48.7%) | 177 (50.0%) | |||
| GG | 131 (24.1%) | 103 (26.8%) | 141 (24.5%) | 93 (26.3%) | |||
| A | 413 (75.9%) | 282 (73.2%) | χ2 = 0.854; df = 1; p = 0.355 | 434 (75.5%) | 261 (73.7%) | χ2 = 0.356; df = 1; p = 0.551 | |
| GG | 131 (24.1%) | 103 (26.8%) | 141 (24.5%) | 93 (26.3%) | |||
| G | 417 (76.7%) | 298 (77.4%) | χ2 = 0.071; df = 1; p = 0.790 | 438 (76.2%) | 277 (78.2%) | χ2 = 0.532; df = 1; p = 0.466 | |
| AA | 127 (23.3%) | 87 (22.6%) | 137 (23.8%) | 77 (21.8%) | |||
| A | 556 (51.1%) | 377 (49.0%) | χ2 = 0.827; df = 1; p = 0.363 | 588 (51.1%) | 345 (48.7%) | χ2 = 1.011; df = 1; p = 0.315 | |
| G | 532 (48.9%) | 393 (51.0%) | 562 (48.9%) | 363 (51.3%) | |||
| COMT rs4818–rs4680 haplotype | CA | 534 (49.1%) | 363 (47.1%) | χ2 = 1.515; df = 3; p = 0.679 | 564 (49.0%) | 333 (47.0%) | χ2 = 3.030; df = 3; p = 0.387 |
| GG | 397 (36.5%) | 284 (36.9%) | 405 (35.2%) | 276 (39.0%) | |||
| CG | 135 (12.4%) | 109 (14.2%) | 157 (13.7%) | 87 (12.3%) | |||
| GA | 22 (2.0%) | 14 (1.8%) | 24 (2.1%) | 12 (1.7%) |
| PANSS Scores | COMT rs4818 | COMT rs4680 | rs4818–rs4680 Haplotype | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CC vs. GC vs. GG | C vs. GG | G vs. CC | C vs. G | AA vs. GA vs. GG | A vs. GG | G vs. AA | A vs. G | ||
| PANSS total | 0.850 | 0.757 | 0.724 | 0.928 | 0.711 | 0.565 | 0.640 | 0.902 | 0.794 |
| PANSS positive | 0.929 | 0.722 | 0.982 | 0.868 | 0.581 | 0.993 | 0.371 | 0.536 | 0.904 |
| P1 | 0.803 | 0.983 | 0.526 | 0.648 | 0.833 | 0.889 | 0.785 | 0.647 | 0.928 |
| P2 | 0.770 | 0.642 | 0.707 | 0.979 | 0.247 | 0.505 | 0.260 | 0.722 | 0.967 |
| P3 | 0.675 | 0.404 | 0.582 | 0.415 | 0.652 | 0.611 | 0.691 | 0.969 | 0.674 |
| P4 | 0.618 | 0.331 | 0.658 | 0.419 | 0.223 | 0.957 | 0.088 | 0.289 | 0.769 |
| P5 | 0.687 | 0.799 | 0.387 | 0.460 | 0.525 | 0.904 | 0.317 | 0.447 | 0.854 |
| P6 | 0.667 | 0.435 | 0.861 | 0.782 | 0.958 | 0.965 | 0.625 | 0.893 | 0.740 |
| P7 | 0.507 | 0.244 | 0.724 | 0.398 | 0.260 | 0.407 | 0.165 | 0.132 | 0.400 |
| PANSS negative | 0.688 | 0.840 | 0.389 | 0.478 | 0.391 | 0.190 | 0.881 | 0.405 | 0.869 |
| N1 | 0.602 | 0.914 | 0.325 | 0.455 | 0.227 | 0.100 | 0.910 | 0.297 | 0.740 |
| N2 | 0.642 | 0.565 | 0.602 | 0.943 | 0.745 | 0.478 | 0.720 | 0.454 | 0.692 |
| N3 | 0.404 | 0.553 | 0.339 | 0.712 | 0.934 | 0.762 | 0.651 | 0.711 | 0.973 |
| N4 | 0.616 | 0.503 | 0.370 | 0.330 | 0.324 | 0.134 | 0.849 | 0.204 | 0.328 |
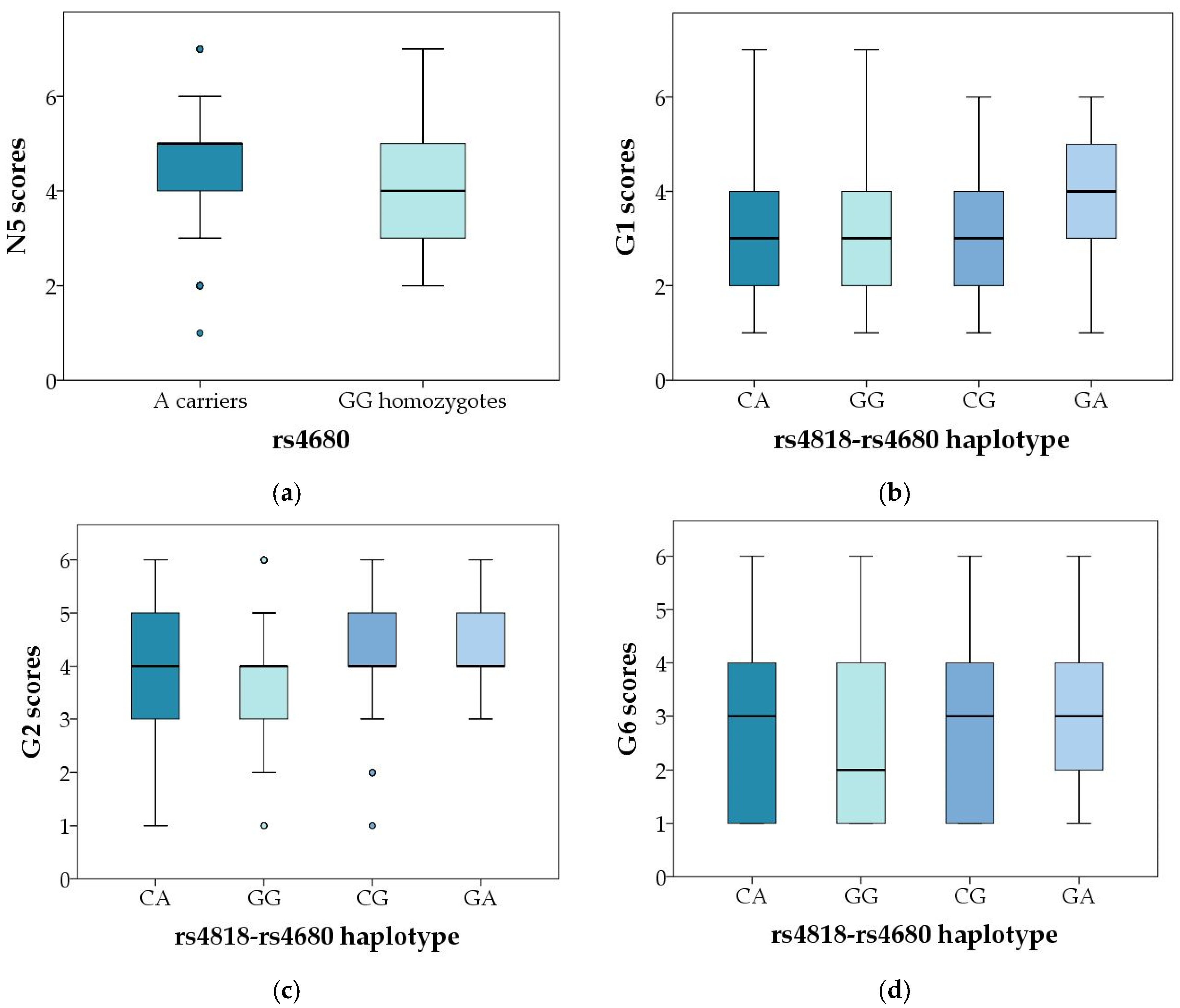
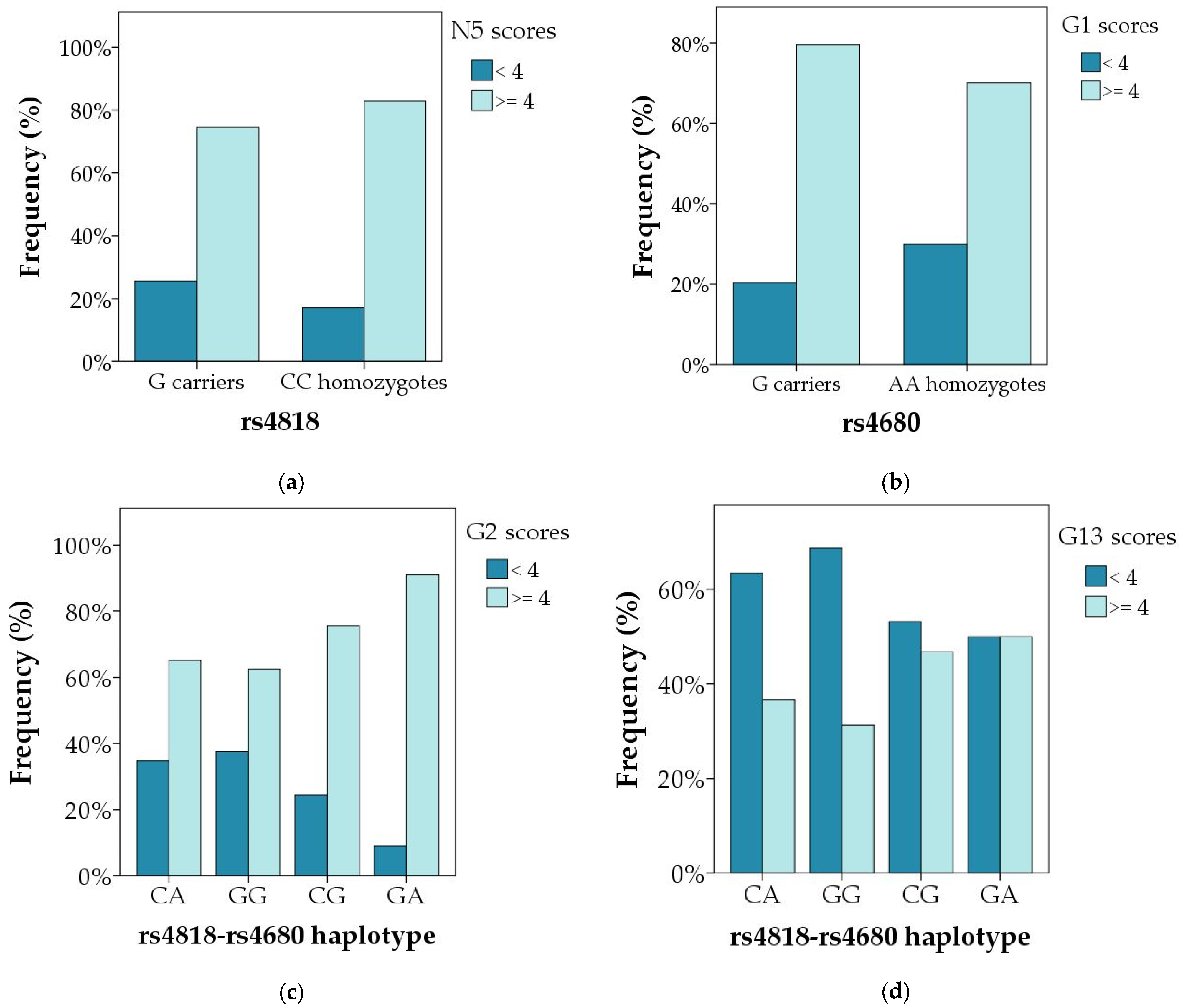
| N5 | 0.272 | 0.210 | 0.173 | 0.109 | 0.060 | 0.023 | 0.695 | 0.141 | 0.401 |
| N6 | 0.871 | 0.755 | 0.618 | 0.609 | 0.581 | 0.416 | 0.951 | 0.780 | 0.719 |
| N7 | 0.802 | 0.554 | 0.923 | 0.814 | 0.223 | 0.191 | 0.532 | 0.690 | 0.967 |
| PANSS general | 0.753 | 0.543 | 0.821 | 0.879 | 0.941 | 0.784 | 0.594 | 0.924 | 0.164 |
| G1 | 0.577 | 0.397 | 0.752 | 0.833 | 0.468 | 0.690 | 0.809 | 0.313 | 0.011 |
| G2 | 0.254 | 0.939 | 0.111 | 0.246 | 0.569 | 0.887 | 0.781 | 0.616 | 0.003 |
| G3 | 0.839 | 0.693 | 0.771 | 0.998 | 0.952 | 0.999 | 0.587 | 0.853 | 0.038 |
| G4 | 0.580 | 0.380 | 0.415 | 0.306 | 0.784 | 0.608 | 0.236 | 0.487 | 0.126 |
| G5 | 0.770 | 0.488 | 0.976 | 0.706 | 0.554 | 0.277 | 0.643 | 0.373 | 0.814 |
| G6 | 0.214 | 0.536 | 0.079 | 0.121 | 0.341 | 0.870 | 0.369 | 0.323 | 0.020 |
| G7 | 0.336 | 0.760 | 0.203 | 0.460 | 0.188 | 0.476 | 0.339 | 0.683 | 0.297 |
| G8 | 0.161 | 0.089 | 0.172 | 0.067 | 0.262 | 0.485 | 0.104 | 0.151 | 0.199 |
| G9 | 0.789 | 0.570 | 0.583 | 0.498 | 0.663 | 0.436 | 0.856 | 0.719 | 0.900 |
| G10 | 0.197 | 0.336 | 0.080 | 0.085 | 0.222 | 0.844 | 0.047 | 0.241 | 0.239 |
| G11 | 0.541 | 0.361 | 0.760 | 0.800 | 0.042 | 0.084 | 0.293 | 0.735 | 0.990 |
| G12 | 0.888 | 0.660 | 0.735 | 0.643 | 0.090 | 0.282 | 0.156 | 0.807 | 0.798 |
| G13 | 0.969 | 0.832 | 0.848 | 0.808 | 0.977 | 0.858 | 0.767 | 0.827 | 0.935 |
| G14 | 0.272 | 0.114 | 0.414 | 0.166 | 0.240 | 0.386 | 0.137 | 0.119 | 0.450 |
| G15 | 0.584 | 0.314 | 0.931 | 0.564 | 0.747 | 0.476 | 0.966 | 0.673 | 0.607 |
| G16 | 0.296 | 0.721 | 0.120 | 0.201 | 0.253 | 0.455 | 0.125 | 0.140 | 0.309 |
| PANSS cognitive | 0.916 | 0.782 | 0.835 | 0.996 | 0.105 | 0.252 | 0.217 | 0.908 | 0.993 |
| PANSS Scores | COMT rs4818 | COMT rs4680 | rs4818–rs4680 Haplotype | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CC vs. GC vs. GG | C vs. GG | G vs. CC | C vs. G | AA vs. GA vs. GG | A vs. GG | G vs. AA | A vs. G | ||
| PANSS total | 0.817 | 0.988 | 0.546 | 0.664 | 0.455 | 0.386 | 0.955 | 0.836 | 0.199 |
| PANSS positive | 0.926 | 0.744 | 0.925 | 0.915 | 0.478 | 0.421 | 0.835 | 0.878 | 0.249 |
| P1 | 0.915 | 0.839 | 0.781 | 0.929 | 0.948 | 0.749 | 0.551 | 0.820 | 0.542 |
| P2 | 0.603 | 0.907 | 0.369 | 0.567 | 0.882 | 0.824 | 0.897 | 0.657 | 0.411 |
| P3 | 0.499 | 0.247 | 0.852 | 0.456 | 0.150 | 0.101 | 0.800 | 0.439 | 0.638 |
| P4 | 0.735 | 0.437 | 0.864 | 0.593 | 0.909 | 0.744 | 0.456 | 0.914 | 0.294 |
| P5 | 0.928 | 0.741 | 0.764 | 0.698 | 0.387 | 0.970 | 0.648 | 0.449 | 0.163 |
| P6 | 0.774 | 0.777 | 0.475 | 0.513 | 0.109 | 0.253 | 0.381 | 0.956 | 0.399 |
| P7 | 0.754 | 0.453 | 0.797 | 0.562 | 0.576 | 0.297 | 0.286 | 0.420 | 0.061 |
| PANSS negative | 0.815 | 0.772 | 0.526 | 0.547 | 0.680 | 0.554 | 0.881 | 0.902 | 0.154 |
| N1 | 0.744 | 0.555 | 0.506 | 0.434 | 0.478 | 0.327 | 0.562 | 0.218 | 0.614 |
| N2 | 0.880 | 0.765 | 0.779 | 0.968 | 0.421 | 0.242 | 0.922 | 0.524 | 0.498 |
| N3 | 0.876 | 0.733 | 0.632 | 0.603 | 0.740 | 0.854 | 0.804 | 0.558 | 0.294 |
| N4 | 0.310 | 0.808 | 0.133 | 0.234 | 0.527 | 0.464 | 0.839 | 0.905 | 0.095 |
| N5 | 0.719 | 0.452 | 0.974 | 0.707 | 0.302 | 0.353 | 0.810 | 0.944 | 0.604 |
| N6 | 0.680 | 0.421 | 0.545 | 0.393 | 0.378 | 0.259 | 0.975 | 0.635 | 0.445 |
| N7 | 0.445 | 0.296 | 0.299 | 0.198 | 0.780 | 0.608 | 0.899 | 0.880 | 0.103 |
| PANSS general | 0.708 | 0.953 | 0.424 | 0.551 | 0.564 | 0.435 | 0.948 | 0.814 | 0.271 |
| G1 | 0.866 | 0.881 | 0.593 | 0.647 | 0.135 | 0.338 | 0.160 | 0.070 | 0.185 |
| G2 | 0.765 | 0.558 | 0.540 | 0.457 | 0.200 | 0.367 | 0.492 | 0.895 | 0.572 |
| G3 | 0.495 | 0.734 | 0.236 | 0.309 | 0.889 | 0.627 | 0.921 | 0.678 | 0.715 |
| G4 | 0.483 | 0.570 | 0.421 | 0.790 | 0.624 | 0.332 | 0.415 | 0.410 | 0.648 |
| G5 | 0.453 | 0.214 | 0.545 | 0.277 | 0.892 | 0.770 | 0.835 | 0.644 | 0.146 |
| G6 | 0.365 | 0.158 | 0.533 | 0.234 | 0.524 | 0.262 | 0.723 | 0.298 | 0.596 |
| G7 | 0.490 | 0.678 | 0.363 | 0.672 | 0.522 | 0.706 | 0.367 | 0.764 | 0.545 |
| G8 | 0.998 | 0.979 | 0.960 | 0.983 | 0.433 | 0.796 | 0.412 | 0.610 | 0.806 |
| G9 | 0.835 | 0.631 | 0.614 | 0.541 | 0.654 | 0.718 | 0.787 | 0.855 | 0.547 |
| G10 | 0.192 | 0.084 | 0.263 | 0.088 | 0.644 | 0.353 | 0.601 | 0.394 | 0.321 |
| G11 | 0.321 | 0.300 | 0.502 | 0.939 | 0.593 | 0.326 | 0.422 | 0.326 | 0.244 |
| G12 | 0.899 | 0.853 | 0.739 | 0.891 | 0.995 | 0.959 | 0.699 | 0.995 | 0.555 |
| G13 | 0.138 | 0.535 | 0.047 | 0.083 | 0.938 | 0.994 | 0.740 | 0.842 | 0.062 |
| G14 | 0.978 | 0.861 | 0.867 | 0.832 | 0.973 | 0.827 | 0.375 | 0.815 | 0.585 |
| G15 | 0.071 | 0.206 | 0.170 | 0.764 | 0.092 | 0.204 | 0.337 | 0.963 | 0.888 |
| G16 | 0.608 | 0.993 | 0.347 | 0.504 | 0.275 | 0.959 | 0.434 | 0.381 | 0.221 |
| PANSS cognitive | 0.753 | 0.841 | 0.453 | 0.524 | 0.856 | 0.578 | 0.495 | 0.619 | 0.221 |
| PANSS Scores | COMT rs4818 | COMT rs4680 | rs4818–rs4680 Haplotype | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CC vs. GC vs. GG | C vs. GG | G vs. CC | C vs. G | AA vs. GA vs. GG | A vs. GG | G vs. AA | A vs. G | ||
| PANSS total | 0.685 | 0.423 | 0.563 | 0.414 | 0.675 | 0.803 | 0.485 | 0.767 | 0.831 |
| PANSS positive | 0.873 | 0.822 | 0.604 | 0.631 | 0.741 | 0.803 | 0.619 | 0.527 | 0.504 |
| P1 | 0.741 | 0.911 | 0.490 | 0.669 | 0.852 | 0.571 | 0.668 | 0.644 | 0.579 |
| P2 | 0.747 | 0.788 | 0.445 | 0.500 | 0.328 | 0.523 | 0.378 | 0.785 | 0.607 |
| P3 | 0.873 | 0.611 | 0.787 | 0.652 | 0.362 | 0.686 | 0.338 | 0.258 | 0.177 |
| P4 | 0.667 | 0.368 | 0.757 | 0.497 | 0.690 | 0.514 | 0.323 | 0.389 | 0.842 |
| P5 | 0.952 | 0.814 | 0.787 | 0.756 | 0.704 | 0.686 | 0.452 | 0.447 | 0.861 |
| P6 | 0.763 | 0.470 | 0.723 | 0.535 | 0.853 | 0.765 | 0.366 | 0.600 | 0.773 |
| P7 | 0.483 | 0.261 | 0.439 | 0.263 | 0.201 | 0.176 | 0.147 | 0.073 | 0.298 |
| PANSS negative | 0.539 | 0.893 | 0.277 | 0.404 | 0.237 | 0.128 | 0.935 | 0.426 | 0.853 |
| N1 | 0.479 | 0.884 | 0.235 | 0.363 | 0.611 | 0.486 | 0.985 | 0.878 | 0.587 |
| N2 | 0.530 | 0.504 | 0.518 | 0.911 | 0.842 | 0.618 | 0.905 | 0.821 | 0.980 |
| N3 | 0.317 | 0.509 | 0.279 | 0.672 | 0.970 | 0.927 | 0.998 | 0.835 | 0.825 |
| N4 | 0.913 | 0.747 | 0.714 | 0.672 | 0.883 | 0.627 | 0.938 | 0.643 | 0.630 |
| N5 | 0.064 | 0.211 | 0.022 | 0.025 | 0.117 | 0.038 | 0.183 | 0.089 | 0.076 |
| N6 | 0.972 | 0.863 | 0.835 | 0.814 | 0.591 | 0.993 | 0.601 | 0.552 | 0.669 |
| N7 | 0.719 | 0.476 | 0.887 | 0.791 | 0.295 | 0.273 | 0.463 | 0.811 | 0.961 |
| PANSS general | 0.777 | 0.906 | 0.531 | 0.704 | 0.410 | 0.366 | 0.325 | 0.190 | 0.181 |
| G1 | 0.094 | 0.046 | 0.146 | 0.041 | 0.280 | 0.386 | 0.024 | 0.136 | 0.054 |
| G2 | 0.382 | 0.771 | 0.168 | 0.263 | 0.532 | 0.589 | 0.968 | 0.890 | 0.003 |
| G3 | 0.974 | 0.873 | 0.837 | 0.821 | 0.798 | 0.916 | 0.938 | 0.635 | 0.027 |
| G4 | 0.677 | 0.832 | 0.379 | 0.467 | 0.253 | 0.744 | 0.027 | 0.223 | 0.187 |
| G5 | 0.191 | 0.136 | 0.606 | 0.690 | 0.332 | 0.198 | 0.999 | 0.538 | 0.882 |
| G6 | 0.398 | 0.802 | 0.236 | 0.481 | 0.878 | 0.929 | 0.907 | 0.824 | 0.096 |
| G7 | 0.541 | 0.289 | 0.978 | 0.574 | 0.418 | 0.415 | 0.795 | 0.956 | 0.534 |
| G8 | 0.284 | 0.245 | 0.163 | 0.115 | 0.310 | 0.509 | 0.176 | 0.178 | 0.388 |
| G9 | 0.901 | 0.672 | 0.980 | 0.842 | 0.505 | 0.992 | 0.345 | 0.499 | 0.585 |
| G10 | 0.475 | 0.346 | 0.665 | 0.859 | 0.832 | 0.548 | 0.477 | 0.660 | 0.044 |
| G11 | 0.281 | 0.281 | 0.442 | 0.991 | 0.067 | 0.125 | 0.291 | 0.823 | 0.985 |
| G12 | 0.774 | 0.517 | 0.621 | 0.497 | 0.108 | 0.102 | 0.492 | 0.583 | 0.780 |
| G13 | 0.915 | 0.765 | 0.707 | 0.676 | 0.892 | 0.953 | 0.636 | 0.819 | 0.930 |
| G14 | 0.362 | 0.156 | 0.565 | 0.258 | 0.431 | 0.473 | 0.139 | 0.223 | 0.453 |
| G15 | 0.259 | 0.128 | 0.281 | 0.124 | 0.498 | 0.923 | 0.089 | 0.541 | 0.132 |
| G16 | 0.203 | 0.546 | 0.162 | 0.499 | 0.639 | 0.834 | 0.558 | 0.711 | 0.555 |
| PANSS cognitive | 0.882 | 0.679 | 0.688 | 0.531 | 0.670 | 0.471 | 0.789 | 0.449 | 0.807 |
| PANSS Scores | COMT rs4818 | COMT rs4680 | rs4818–rs4680 Haplotype | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CC vs. GC vs. GG | C vs. GG | G vs. CC | C vs. G | AA vs. GA vs. GG | A vs. GG | G vs. AA | A vs. G | ||
| PANSS total | 0.276 | 0.344 | 0.123 | 0.112 | 0.032 | 0.403 | 0.071 | 0.034 | 0.098 |
| PANSS positive | 0.309 | 0.189 | 0.770 | 0.625 | 0.709 | 0.451 | 0.928 | 0.407 | 0.791 |
| P1 | 0.526 | 0.543 | 0.491 | 0.870 | 0.765 | 0.702 | 0.989 | 0.971 | 0.334 |
| P2 | 0.403 | 0.879 | 0.190 | 0.314 | 0.855 | 0.590 | 0.756 | 0.713 | 0.517 |
| P3 | 0.807 | 0.566 | 0.921 | 0.815 | 0.865 | 0.601 | 0.700 | 0.717 | 0.986 |
| P4 | 0.843 | 0.802 | 0.681 | 0.875 | 0.860 | 0.969 | 0.322 | 0.779 | 0.354 |
| P5 | 0.825 | 0.546 | 0.940 | 0.709 | 0.403 | 0.640 | 0.706 | 0.740 | 0.136 |
| P6 | 0.904 | 0.660 | 0.815 | 0.690 | 0.218 | 0.303 | 0.512 | 0.956 | 0.285 |
| P7 | 0.932 | 0.722 | 0.814 | 0.723 | 0.393 | 0.172 | 0.398 | 0.254 | 0.319 |
| PANSS negative | 0.947 | 0.776 | 0.797 | 0.740 | 0.264 | 0.925 | 0.371 | 0.393 | 0.758 |
| N1 | 0.768 | 0.575 | 0.531 | 0.460 | 0.468 | 0.256 | 0.633 | 0.220 | 0.645 |
| N2 | 0.731 | 0.453 | 0.980 | 0.678 | 0.322 | 0.134 | 0.660 | 0.239 | 0.378 |
| N3 | 0.998 | 0.994 | 0.955 | 0.972 | 0.877 | 0.774 | 0.847 | 0.998 | 0.156 |
| N4 | 0.787 | 0.932 | 0.500 | 0.602 | 0.929 | 0.712 | 0.537 | 0.710 | 0.243 |
| N5 | 0.603 | 0.802 | 0.407 | 0.650 | 0.075 | 0.071 | 0.987 | 0.476 | 0.551 |
| N6 | 0.668 | 0.400 | 0.560 | 0.391 | 0.434 | 0.268 | 0.862 | 0.586 | 0.281 |
| N7 | 0.604 | 0.352 | 0.500 | 0.332 | 0.671 | 0.404 | 0.710 | 0.605 | 0.049 |
| PANSS general | 0.028 | 0.083 | 0.012 | 0.007 | 0.005 | 0.006 | 0.045 | 0.001 | 0.007 |
| G1 | 0.645 | 0.740 | 0.479 | 0.746 | 0.381 | 0.543 | 0.128 | 0.221 | 0.517 |
| G2 | 0.864 | 0.597 | 0.943 | 0.741 | 0.411 | 0.501 | 0.586 | 0.924 | 0.942 |
| G3 | 0.813 | 0.909 | 0.578 | 0.740 | 0.669 | 0.925 | 0.619 | 0.556 | 0.424 |
| G4 | 0.605 | 0.414 | 0.790 | 0.807 | 0.878 | 0.685 | 0.563 | 0.606 | 0.727 |
| G5 | 0.570 | 0.291 | 0.660 | 0.384 | 0.766 | 0.678 | 0.740 | 0.997 | 0.093 |
| G6 | 0.587 | 0.730 | 0.302 | 0.362 | 0.959 | 0.995 | 0.821 | 0.864 | 0.533 |
| G7 | 0.321 | 0.734 | 0.135 | 0.216 | 0.829 | 0.890 | 0.279 | 0.647 | 0.600 |
| G8 | 0.861 | 0.586 | 0.813 | 0.648 | 0.347 | 0.582 | 0.453 | 0.757 | 0.871 |
| G9 | 0.503 | 0.979 | 0.267 | 0.425 | 0.463 | 0.953 | 0.410 | 0.509 | 0.296 |
| G10 | 0.783 | 0.485 | 0.799 | 0.582 | 0.652 | 0.423 | 0.961 | 0.685 | 0.599 |
| G11 | 0.422 | 0.428 | 0.476 | 0.934 | 0.943 | 0.831 | 0.680 | 0.979 | 0.517 |
| G12 | 0.454 | 0.246 | 0.955 | 0.565 | 0.625 | 0.446 | 0.847 | 0.768 | 0.462 |
| G13 | 0.131 | 0.356 | 0.046 | 0.057 | 0.912 | 0.728 | 0.730 | 0.663 | 0.025 |
| G14 | 0.878 | 0.623 | 0.765 | 0.637 | 0.884 | 0.839 | 0.790 | 0.929 | 0.638 |
| G15 | 0.121 | 0.171 | 0.333 | 0.966 | 0.141 | 0.111 | 0.504 | 0.505 | 0.834 |
| G16 | 0.283 | 0.964 | 0.140 | 0.308 | 0.221 | 0.873 | 0.345 | 0.261 | 0.049 |
| PANSS cognitive | 0.296 | 0.284 | 0.152 | 0.635 | 0.349 | 0.574 | 0.106 | 0.862 | 0.700 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sagud, M.; Tudor, L.; Nedic Erjavec, G.; Nikolac Perkovic, M.; Uzun, S.; Mimica, N.; Madzarac, Z.; Zivkovic, M.; Kozumplik, O.; Konjevod, M.; et al. Genotypic and Haplotypic Association of Catechol-O-Methyltransferase rs4680 and rs4818 Gene Polymorphisms with Particular Clinical Symptoms in Schizophrenia. Genes 2023, 14, 1358. https://doi.org/10.3390/genes14071358
Sagud M, Tudor L, Nedic Erjavec G, Nikolac Perkovic M, Uzun S, Mimica N, Madzarac Z, Zivkovic M, Kozumplik O, Konjevod M, et al. Genotypic and Haplotypic Association of Catechol-O-Methyltransferase rs4680 and rs4818 Gene Polymorphisms with Particular Clinical Symptoms in Schizophrenia. Genes. 2023; 14(7):1358. https://doi.org/10.3390/genes14071358
Chicago/Turabian StyleSagud, Marina, Lucija Tudor, Gordana Nedic Erjavec, Matea Nikolac Perkovic, Suzana Uzun, Ninoslav Mimica, Zoran Madzarac, Maja Zivkovic, Oliver Kozumplik, Marcela Konjevod, and et al. 2023. "Genotypic and Haplotypic Association of Catechol-O-Methyltransferase rs4680 and rs4818 Gene Polymorphisms with Particular Clinical Symptoms in Schizophrenia" Genes 14, no. 7: 1358. https://doi.org/10.3390/genes14071358
APA StyleSagud, M., Tudor, L., Nedic Erjavec, G., Nikolac Perkovic, M., Uzun, S., Mimica, N., Madzarac, Z., Zivkovic, M., Kozumplik, O., Konjevod, M., Svob Strac, D., & Pivac, N. (2023). Genotypic and Haplotypic Association of Catechol-O-Methyltransferase rs4680 and rs4818 Gene Polymorphisms with Particular Clinical Symptoms in Schizophrenia. Genes, 14(7), 1358. https://doi.org/10.3390/genes14071358