
Genome-Wide Association Study of Blood Mercury in European Pregnant Women and Children

 , , , , ,
, , , , ,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Overview
2.2. The Avon Longitudinal Study of Parents and Children (ALSPAC)
2.3. The Human Early Life Exposome (HELIX)
2.4. SNP Heritability
2.5. Genome-Wide Association Testing
2.6. In Silico Functional Analysis
3. Results
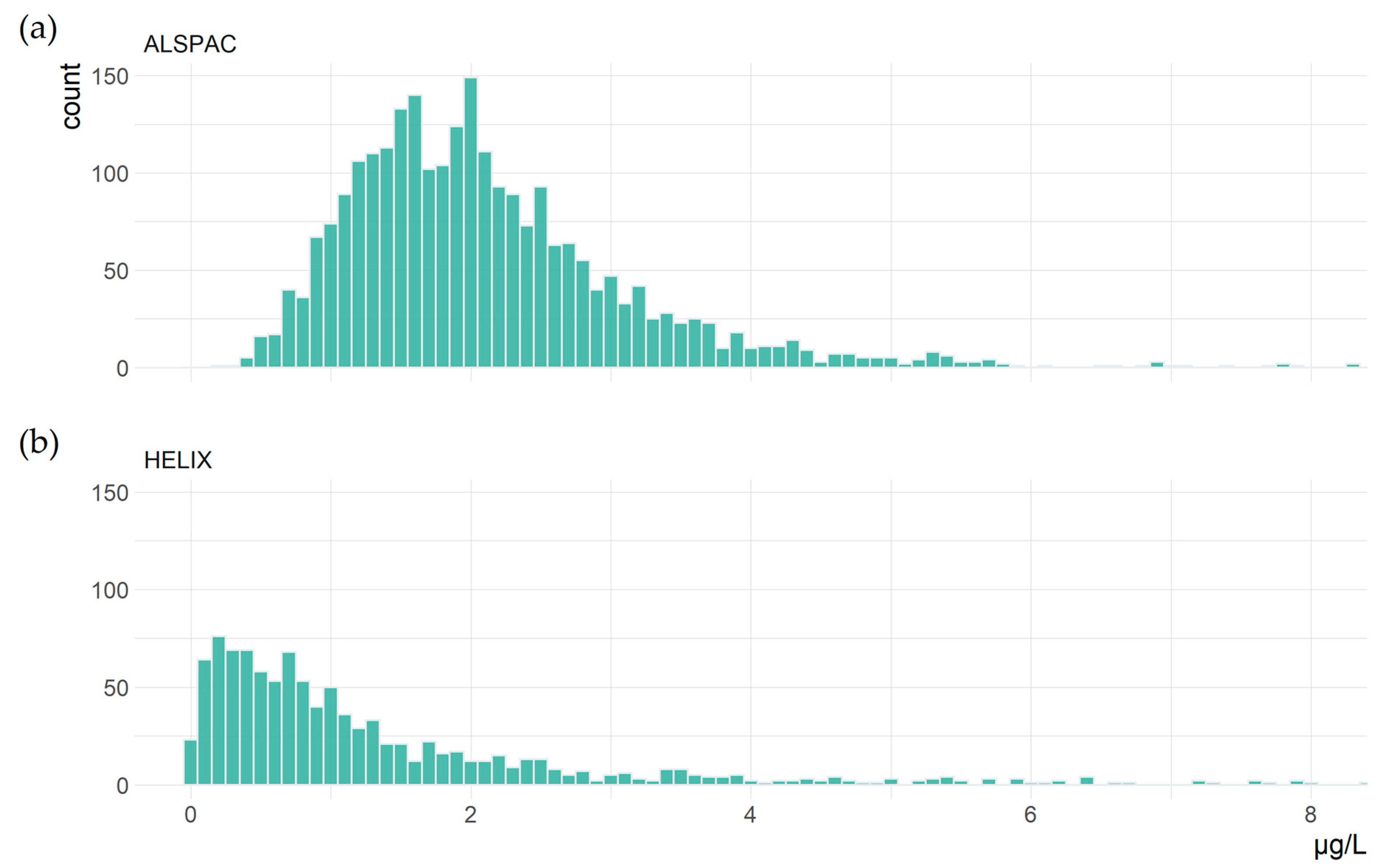
3.1. Study Characteristics
3.2. SNP Heritability
3.3. Genome-Wide Association Testing
3.4. In Silico Functional Analysis
3.5. Associations in Previous Candidate Variants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gworek, B.; Dmuchowski, W.; Baczewska-Dąbrowska, A.H. Mercury in the terrestrial environment: A review. Environ. Sci. Eur. 2020, 32, 128. [Google Scholar] [CrossRef]
- Sunderland, E.M.; Selin, N.E. Future trends in environmental mercury concentrations: Implications for prevention strategies. Environ. Health 2013, 12, 2. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, Z.; Huang, S.; Zhang, P.; Peng, Y.; Wu, P.; Gu, J.; Dutkiewicz, S.; Zhang, H.; Wu, S.; et al. Global health effects of future atmospheric mercury emissions. Nat. Commun. 2021, 12, 3035. [Google Scholar] [CrossRef]
- Broussard, L.A.; Hammett-Stabler, C.A.; Winecker, R.E.; Ropero-Miller, J.D. The Toxicology of Mercury. Lab. Med. 2002, 33, 614–625. [Google Scholar] [CrossRef]
- World Health Organization. Children’s Exposure to Mercury Compounds; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Driscoll, C.T.; Mason, R.P.; Chan, H.M.; Jacob, D.J.; Pirrone, N. Mercury as a Global Pollutant: Sources, Pathways, and Effects. Environ. Sci. Technol. 2013, 47, 4967–4983. [Google Scholar] [CrossRef] [PubMed]
- Selin, N.E.; Jacob, D.J.; Park, R.J.; Yantosca, R.M.; Strode, S.; Jaeglé, L.; Jaffe, D. Chemical cycling and deposition of atmospheric mercury: Global constraints from observations. J. Geophys. Res. Atmos. 2007, 112, 1–14. [Google Scholar] [CrossRef]
- Parks, J.M.; Johs, A.; Podar, M.; Bridou, R.; Hurt, R.A., Jr.; Smith, S.D.; Tomanicek, S.J.; Qian, Y.; Brown, S.D.; Brandt, C.C. The genetic basis for bacterial mercury methylation. Science 2013, 339, 1332–1335. [Google Scholar] [CrossRef]
- Hammerschmidt, C.R.; Fitzgerald, W.F. Methylmercury in Freshwater Fish Linked to Atmospheric Mercury Deposition. Environ. Sci. Technol. 2006, 40, 7764–7770. [Google Scholar] [CrossRef]
- Médieu, A.; Point, D.; Itai, T.; Angot, H.; Buchanan, P.J.; Allain, V.; Fuller, L.; Griffiths, S.; Gillikin, D.P.; Sonke, J.E.; et al. Evidence that Pacific tuna mercury levels are driven by marine methylmercury production and anthropogenic inputs. Proc. Natl. Acad. Sci. USA 2022, 119, e2113032119. [Google Scholar] [CrossRef]
- Golding, J.; Steer, C.D.; Hibbeln, J.R.; Emmett, P.M.; Lowery, T.; Jones, R. Dietary Predictors of Maternal Prenatal Blood Mercury Levels in the ALSPAC Birth Cohort Study. Environ. Health Perspect. 2013, 121, 1214–1218. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, O.; Zamorano, P.; Garcia, O.; Bastías, J.M. Arsenic, cadmium, mercury, sodium, and potassium concentrations in common foods and estimated daily intake of the population in Valdivia (Chile) using a total diet study. Food Chem. Toxicol. 2017, 109, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Cho, Y.-H.; Park, S.-O.; Kye, S.-H.; Kim, B.-H.; Hahm, T.-S.; Kim, M.; Lee, J.O.; Kim, C.-I. Dietary exposure of the Korean population to arsenic, cadmium, lead and mercury. J. Food Compos. Anal. 2006, 19, S31–S37. [Google Scholar] [CrossRef]
- Almerud, P.; Zamaratskaia, G.; Lindroos, A.K.; Bjermo, H.; Andersson, E.M.; Lundh, T.; Ankarberg, E.H.; Lignell, S. Cadmium, total mercury, and lead in blood and associations with diet, sociodemographic factors, and smoking in Swedish adolescents. Environ. Res. 2021, 197, 110991. [Google Scholar] [CrossRef]
- Berlin, M.; Zalups, R.K.; Fowler, B.A. Chapter 46–Mercury. In Handbook on the Toxicology of Metals, 4th ed.; Nordberg, G.F., Fowler, B.A., Nordberg, M., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 1013–1075. [Google Scholar]
- United Nations Environment Programme; World Health Organization. Guidance for Identifying Populations at Risk from Mercury Exposure; UNEP: Nairobi, Kenya, 2008. [Google Scholar]
- Bridges, C.C.; Zalups, R.K. Mechanisms involved in the transport of mercuric ions in target tissues. Arch. Toxicol. 2017, 91, 63–81. [Google Scholar] [CrossRef]
- Park, J.-D.; Zheng, W. Human Exposure and Health Effects of Inorganic and Elemental Mercury. J. Prev. Med. Public Health 2012, 45, 344–352. [Google Scholar] [CrossRef]
- Yin, Z.; Jiang, H.; Syversen, T.; Rocha, J.B.T.; Farina, M.; Aschner, M. The methylmercury-l-cysteine conjugate is a substrate for the L-type large neutral amino acid transporter. J. Neurochem. 2008, 107, 1083–1090. [Google Scholar] [CrossRef]
- Mortensen, M.E.; Caudill, S.P.; Caldwell, K.L.; Ward, C.D.; Jones, R.L. Total and methyl mercury in whole blood measured for the first time in the U.S. population: NHANES 2011–2012. Environ. Res. 2014, 134, 257–264. [Google Scholar] [CrossRef]
- Yokoo, E.M.; Valente, J.G.; Grattan, L.; Schmidt, S.L.; Platt, I.; Silbergeld, E.K. Low level methylmercury exposure affects neuropsychological function in adults. Environ. Health 2003, 2, 8. [Google Scholar] [CrossRef]
- Bridges, C.C.; Krasnikov, B.F.; Joshee, L.; Pinto, J.T.; Hallen, A.; Li, J.; Zalups, R.K.; Cooper, A.J. New insights into the metabolism of organomercury compounds: Mercury-containing cysteine S-conjugates are substrates of human glutamine transaminase K and potent inactivators of cystathionine γ-lyase. Arch. Biochem. Biophys. 2012, 517, 20–29. [Google Scholar] [CrossRef]
- Bernhoft, R.A. Mercury toxicity and treatment: A review of the literature. J. Environ. Public Health 2012, 2012, 460508. [Google Scholar] [CrossRef]
- Grotto, D.; Valentini, J.; Fillion, M.; Passos, C.J.S.; Garcia, S.C.; Mergler, D.; Barbosa, F., Jr. Mercury exposure and oxidative stress in communities of the Brazilian Amazon. Sci. Total Environ. 2010, 408, 806–811. [Google Scholar] [CrossRef]
- Syversen, T.; Kaur, P. The toxicology of mercury and its compounds. J. Trace Elem. Med. Biol. 2012, 26, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Zefferino, R.; Piccoli, C.; Ricciardi, N.; Scrima, R.; Capitanio, N. Possible Mechanisms of Mercury Toxicity and Cancer Promotion: Involvement of Gap Junction Intercellular Communications and Inflammatory Cytokines. Oxidative Med. Cell. Longev. 2017, 2017, 7028583. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Momtaz, S.; Abdollahi, M. The relationship between mercury exposure and epigenetic alterations regarding human health, risk assessment and diagnostic strategies. J. Trace Elements Med. Biol. 2019, 52, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Orr, S.E.; Barnes, M.C.; Joshee, L.; Uchakina, O.; McKallip, R.J.; Bridges, C.C. Potential mechanisms of cellular injury following exposure to a physiologically relevant species of inorganic mercury. Toxicol. Lett. 2019, 304, 13–20. [Google Scholar] [CrossRef]
- Ye, B.-J.; Kim, B.-G.; Jeon, M.-J.; Kim, S.-Y.; Kim, H.-C.; Jang, T.-W.; Chae, H.-J.; Choi, W.-J.; Ha, M.-N.; Hong, Y.-S. Evaluation of mercury exposure level, clinical diagnosis and treatment for mercury intoxication. Ann. Occup. Environ. Med. 2016, 28, 5. [Google Scholar] [CrossRef] [PubMed]
- Do, S.Y.; Lee, C.G.; Kim, J.Y.; Moon, Y.H.; Kim, M.S.; Bae, I.H.; Song, H.S. Cases of acute mercury poisoning by mercury vapor exposure during the demolition of a fluorescent lamp factory. Ann. Occup. Environ. Med. 2017, 29, 19. [Google Scholar] [CrossRef]
- Asano, S.; Eto, K.; Kurisaki, E.; Gunji, H.; Hiraiwa, K.; Sato, M.; Sato, H.; Hasuike, M.; Hagiwara, N.; Wakasa, H. Acute inorganic mercury vapor inhalation poisoning. Pathol. Int. 2000, 50, 169–174. [Google Scholar] [CrossRef]
- Clarkson, T.W. The three modern faces of mercury. Environ. Health Perspect. 2002, 110 (Suppl. 1), 11–23. [Google Scholar] [CrossRef]
- Bridges, C.C.; Zalups, R.K. Transport of Inorganic Mercury and Methylmercury in Target Tissues and Organs. J. Toxicol. Environ. Health Part B Crit. Rev. 2010, 13, 385–410. [Google Scholar] [CrossRef]
- Fisher, J.F.; World Health Organization. Elemental Mercury and Inorganic Mercury Compounds: Human Health Aspects; World Health Organization: Geneva, Switzerland, 2003. [Google Scholar]
- Jo, S.; Woo, H.D.; Kwon, H.-J.; Oh, S.-Y.; Park, J.-D.; Hong, Y.-S.; Pyo, H.; Park, K.S.; Ha, M.; Kim, H.; et al. Estimation of the Biological Half-Life of Methylmercury Using a Population Toxicokinetic Model. Int. J. Environ. Res. Public Health 2015, 12, 9054–9067. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.F.; Singh, K.; Chan, H.M. Mercury Exposure, Blood Pressure, and Hypertension: A Systematic Review and Dose–response Meta-analysis. Environ. Health Perspect. 2018, 126, 076002. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.F.; Lowe, M.; Chan, H.M. Mercury exposure, cardiovascular disease, and mortality: A systematic review and dose-response meta-analysis. Environ. Res. 2021, 193, 110538. [Google Scholar] [CrossRef]
- Bridges, C.C.; Zalups, R.K. The aging kidney and the nephrotoxic effects of mercury. J. Toxicol. Environ. Health Part B Crit. Rev. 2017, 20, 55–80. [Google Scholar] [CrossRef]
- Masley, S.C.; Masley, L.V.; Gualtieri, C. Effect of mercury levels and seafood intake on cognitive function in middle-aged adults. Integr. Med. 2012, 11, 32–40. [Google Scholar]
- National Research Council. Toxicological Effects of Methylmercury; The National Academies Press: Washington, DC, USA, 2000. [Google Scholar]
- Dack, K.; Fell, M.; Taylor, C.M.; Havdahl, A.; Lewis, S.J. Mercury and Prenatal Growth: A Systematic Review. Int. J. Environ. Res. Public Health 2021, 18, 7140. [Google Scholar] [CrossRef]
- Dack, K.; Fell, M.; Taylor, C.M.; Havdahl, A.; Lewis, S.J. Prenatal Mercury Exposure and Neurodevelopment up to the Age of 5 Years: A Systematic Review. Int. J. Environ. Res. Public Health 2022, 19, 1976. [Google Scholar] [CrossRef]
- Barcelos, G.R.M.; Grotto, D.; de Marco, K.C.; Valentini, J.; Lengert, A.v.H.; de Oliveira, A.S.; Garcia, S.C.; Braga, G.L.; Engström, K.S.; de Syllos Cólus, I.M.; et al. Polymorphisms in glutathione-related genes modify mercury concentrations and antioxidant status in subjects environmentally exposed to methylmercury. Sci. Total. Environ. 2013, 463–464, 319–325. [Google Scholar] [CrossRef]
- Engström, K.S.; Strömberg, U.; Lundh, T.; Johansson, I.; Vessby, B.; Hallmans, G.; Skerfving, S.; Broberg, K. Genetic Variation in Glutathione-Related Genes and Body Burden of Methylmercury. Environ. Health Perspect. 2008, 116, 734–739. [Google Scholar] [CrossRef]
- Gundacker, C.; Komarnicki, G.; Jagiello, P.; Gencikova, A.; Dahmen, N.; Wittmann, K.J.; Gencik, M. Glutathione-S-transferase polymorphism, metallothionein expression, and mercury levels among students in Austria. Sci. Total Environ. 2007, 385, 37–47. [Google Scholar] [CrossRef]
- Wang, Y.; Goodrich, J.M.; Gillespie, B.; Werner, R.; Basu, N.; Franzblau, A. An Investigation of Modifying Effects of Metallothionein Single-Nucleotide Polymorphisms on the Association between Mercury Exposure and Biomarker Levels. Environ. Health Perspect. 2012, 120, 530–534. [Google Scholar] [CrossRef]
- Ng, S.; Lin, C.-C.; Hwang, Y.-H.; Hsieh, W.-S.; Liao, H.-F.; Chen, P.-C. Mercury, APOE, and children’s neurodevelopment. NeuroToxicology 2013, 37, 85–92. [Google Scholar] [CrossRef]
- Julvez, J.; Smith, G.D.; Golding, J.; Ring, S.; Pourcain, B.S.; Gonzalez, J.R.; Grandjean, P. Prenatal Methylmercury Exposure and Genetic Predisposition to Cognitive Deficit at Age 8 Years. Epidemiology 2013, 24, 643–650. [Google Scholar] [CrossRef]
- Bjørklund, G.; Aaseth, J.; Ajsuvakova, O.P.; Nikonorov, A.A.; Skalny, A.V.; Skalnaya, M.G.; Tinkov, A.A. Molecular interaction between mercury and selenium in neurotoxicity. Coord. Chem. Rev. 2017, 332, 30–37. [Google Scholar] [CrossRef]
- Franciscato, C.; Moraes-Silva, L.; Duarte, F.; Oliveira, C.; Ineu, R.; Flores, E.; Dressler, V.; Peixoto, N.; Pereira, M. Delayed biochemical changes induced by mercury intoxication are prevented by zinc pre-exposure. Ecotoxicol. Environ. Saf. 2011, 74, 480–486. [Google Scholar] [CrossRef]
- Mesquita, M.; Pedroso, T.F.; Oliveira, C.S.; Oliveira, V.A.; Santos, R.F.D.; Bizzi, C.A.; Pereira, M.E. Effects of zinc against mercury toxicity in female rats 12 and 48 hours after HgCl2 exposure. EXCLI J. 2016, 15, 256–267. [Google Scholar] [CrossRef]
- Singh, N.; Gupta, V.K.; Kumar, A.; Sharma, B. Synergistic Effects of Heavy Metals and Pesticides in Living Systems. Front. Chem. 2017, 5, 70. [Google Scholar] [CrossRef]
- Hernandez, A.F.; Buha, A.; Constantin, C.; Wallace, D.R.; Sarigiannis, D.; Neagu, M.; Antonijevic, B.; Hayes, A.W.; Wilks, M.F.; Tsatsakis, A. Critical assessment and integration of separate lines of evidence for risk assessment of chemical mixtures. Arch. Toxicol. 2019, 93, 2741–2757. [Google Scholar] [CrossRef]
- Warrington, N.M.; Zhu, G.; Dy, V.; Heath, A.C.; Madden, P.A.; Hemani, G.; Kemp, J.P.; Mcmahon, G.; Pourcain, B.S.; Timpson, N.J.; et al. Genome-wide association study of blood lead shows multiple associations near ALAD. Hum. Mol. Genet. 2015, 24, 3871–3879. [Google Scholar] [CrossRef]
- Evans, D.M.; Zhu, G.; Dy, V.; Heath, A.C.; Madden, P.A.F.; Kemp, J.P.; McMahon, G.; St Pourcain, B.; Timpson, N.J.; Golding, J.; et al. Genome-wide association study identifies loci affecting blood copper, selenium and zinc. Hum. Mol. Genet. 2013, 22, 3998–4006. [Google Scholar] [CrossRef]
- Cornelis, M.C.; Fornage, M.; Foy, M.; Xun, P.; Gladyshev, V.N.; Morris, S.; Chasman, D.I.; Hu, F.B.; Rimm, E.B.; Kraft, P.; et al. Genome-wide association study of selenium concentrations. Hum. Mol. Genet. 2015, 24, 1469–1477. [Google Scholar] [CrossRef]
- Davies, N.M.; Holmes, M.V.; Smith, G.D. Reading Mendelian randomisation studies: A guide, glossary, and checklist for clinicians. BMJ 2018, 362, k601. [Google Scholar] [CrossRef]
- Duncan, L.E.; Ostacher, M.; Ballon, J. How genome-wide association studies (GWAS) made traditional candidate gene studies obsolete. Neuropsychopharmacology 2019, 44, 1518–1523. [Google Scholar] [CrossRef]
- Tam, V.; Patel, N.; Turcotte, M.; Bossé, Y.; Paré, G.; Meyre, D. Benefits and limitations of genome-wide association studies. Nat. Rev. Genet. 2019, 20, 467–484. [Google Scholar] [CrossRef]
- Bell, S.; Rigas, A.S.; Ferkingstad, E.; Allara, E.; Bjornsdottir, G.; Ramond, A.; Sørensen, E.; Halldorsson, G.H.; Paul, D.S.; Burgdorf, K.S.; et al. A genome-wide meta-analysis yields 46 new loci associating with biomarkers of iron homeostasis. Commun. Biol. 2021, 4, 156. [Google Scholar] [CrossRef]
- Ng, E.; Lind, P.M.; Lindgren, C.; Ingelsson, E.; Mahajan, A.; Morris, A.; Lind, L. Genome-wide association study of toxic metals and trace elements reveals novel associations. Hum. Mol. Genet. 2015, 24, 4739–4745. [Google Scholar] [CrossRef]
- Fraser, A.; Macdonald-Wallis, C.; Tilling, K.; Boyd, A.; Golding, J.; Smith, G.D.; Henderson, J.; Macleod, J.; Molloy, L.; Ness, A.; et al. Cohort Profile: The Avon Longitudinal Study of Parents and Children: ALSPAC mothers cohort. Eur. J. Endocrinol. 2012, 42, 97–110. [Google Scholar] [CrossRef]
- Boyd, A.; Golding, J.; Macleod, J.; A Lawlor, D.; Fraser, A.; Henderson, J.; Molloy, L.; Ness, A.; Ring, S.; Smith, G.D. Cohort Profile: The ‘Children of the 90s’—The index offspring of the Avon Longitudinal Study of Parents and Children. Eur. J. Endocrinol. 2012, 42 (Suppl. 3), 111–127. [Google Scholar] [CrossRef]
- Taylor, C.M.; Golding, J.; Hibbeln, J.; Emond, A.M. Environmental Factors Predicting Blood Lead Levels in Pregnant Women in the UK: The ALSPAC Study. PLoS ONE 2013, 8, e72371. [Google Scholar] [CrossRef]
- Hornung, R.W.; Reed, L.D. Estimation of Average Concentration in the Presence of Nondetectable Values. Appl. Occup. Environ. Hyg. 1990, 5, 46–51. [Google Scholar] [CrossRef]
- Pembrey, M. The Avon Longitudinal Study of Parents and Children (ALSPAC): A resource for genetic epidemiology. Eur. J. Endocrinol. 2004, 151 (Suppl. 3), U125–U129. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Steinthorsdottir, V.; McGinnis, R.; Williams, N.O.; Stefansdottir, L.; Thorleifsson, G.; Shooter, S.; Fadista, J.; Sigurdsson, J.K.; Auro, K.M.; Berezina, G.; et al. Genetic predisposition to hypertension is associated with preeclampsia in European and Central Asian women. Nat. Commun. 2020, 11, 5976. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Morris, T.T.; Davies, N.M.; Smith, G.D. Can education be personalised using pupils’ genetic data? eLife 2020, 9, e49962. [Google Scholar] [CrossRef] [PubMed]
- ALSPAC Team. ALSPAC OMICs Data Catalogue. Available online: https://alspac.github.io/omics_documentation/alspac_omics_data_catalogue.html (accessed on 16 October 2023).
- Delaneau, O.; Marchini, J.; Zagury, J.-F. A linear complexity phasing method for thousands of genomes. Nat. Methods 2012, 9, 179–181. [Google Scholar] [CrossRef]
- Das, S.; Forer, L.; Schönherr, S.; Sidore, C.; Locke, A.E.; Kwong, A.; Vrieze, S.I.; Chew, E.Y.; Levy, S.; McGue, M.; et al. Next-generation genotype imputation service and methods. Nat. Genet. 2016, 48, 1284–1287. [Google Scholar] [CrossRef]
- Vrijheid, M.; Slama, R.; Robinson, O.; Chatzi, L.; Coen, M.; Hazel, P.v.D.; Thomsen, C.; Wright, J.; Athersuch, T.J.; Avellana, N.; et al. The Human Early-Life Exposome (HELIX): Project Rationale and Design. Environ. Health Perspect. 2014, 122, 535–544. [Google Scholar] [CrossRef]
- Maitre, L.; De Bont, J.; Casas, M.; Robinson, O.; Aasvang, G.M.; Agier, L.; Andrušaitytė, S.; Ballester, F.; Basagaña, X.; Borràs, E.; et al. Human Early Life Exposome (HELIX) study: A European population-based exposome cohort. BMJ Open 2018, 8, e021311. [Google Scholar] [CrossRef]
- Magnus, P.; Birke, C.; Vejrup, K.; Haugan, A.; Alsaker, E.; Daltveit, A.K.; Handal, M.; Haugen, M.; Høiseth, G.; Knudsen, G.P.; et al. Cohort Profile Update: The Norwegian Mother and Child Cohort Study (MoBa). Int. J. Epidemiol. 2016, 45, 382–388. [Google Scholar] [CrossRef]
- Haug, L.S.; Sakhi, A.K.; Cequier, E.; Casas, M.; Maitre, L.; Basagana, X.; Andrusaityte, S.; Chalkiadaki, G.; Chatzi, L.; Coen, M.; et al. In-utero and childhood chemical exposome in six European mother-child cohorts. Environ. Int. 2018, 121, 751–763. [Google Scholar] [CrossRef]
- Rodushkin, I.; Axelsson, M.D. Application of double focusing sector field ICP-MS for multielemental characterization of human hair and nails. Part I. Analytical methodology. Sci. Total. Environ. 2000, 250, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Serra, B.; Maitre, L.; E Lau, C.-H.; Siskos, A.P.; Gützkow, K.B.; Andrušaitytė, S.; Casas, M.; Cadiou, S.; Chatzi, L.; González, J.R.; et al. Urinary metabolite quantitative trait loci in children and their interaction with dietary factors. Hum. Mol. Genet. 2021, 29, 3830–3844. [Google Scholar] [CrossRef]
- Bulik-Sullivan, B.K.; Loh, P.R.; Finucane, H.K.; Ripke, S.; Yang, J.; Schizophrenia Working Group of the Psychiatric Genomics Consortium; Patterson, N.; Daly, M.J.; Price, A.L.; Neale, B.M. LD Score regression distinguishes confounding from polygenicity in genome-wide association studies. Nat. Genet. 2015, 47, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Bulik-Sullivan, B.; Finucane, H.K.; Anttila, V.; Gusev, A.; Day, F.R.; Loh, P.-R.; Duncan, L.; Perry, J.R.B.; Patterson, N.; Robinson, E.B.; et al. An atlas of genetic correlations across human diseases and traits. Nat. Genet. 2015, 47, 1236–1241. [Google Scholar] [CrossRef] [PubMed]
- Ni, G.; Moser, G.; Wray, N.R.; Lee, S.H.; Ripke, S.; Neale, B.M.; Corvin, A.; Walters, J.T.; Farh, K.-H.; Holmans, P.A.; et al. Estimation of Genetic Correlation via Linkage Disequilibrium Score Regression and Genomic Restricted Maximum Likelihood. Am. J. Hum. Genet. 2018, 102, 1185–1194. [Google Scholar] [CrossRef]
- Marchini, J.; Howie, B.; Myers, S.; McVean, G.; Donnelly, P. A new multipoint method for genome-wide association studies by imputation of genotypes. Nat. Genet. 2007, 39, 906–913. [Google Scholar] [CrossRef]
- Pagès, H.S. Hsapiens. dbSNPGRCh37: SNP Locations for Homo Sapiens (dbSNP Build 144). R Packag. version 0.99. 2017, Volume 20. Available online: https://bioconductor.org/packages/release/data/annotation/html/SNPlocs.Hsapiens.dbSNP144.GRCh37.html (accessed on 21 November 2023).
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating high-throughput genomic analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef]
- Turner, S.D. qqman: An R package for visualizing GWAS results using Q-Q and manhattan plots. J. Open Source Softw. 2018, 3, 731. [Google Scholar] [CrossRef]
- Hemani, G.; Zheng, J.; Elsworth, B.; Wade, K.H.; Haberland, V.; Baird, D.; Laurin, C.; Burgess, S.; Bowden, J.; Langdon, R.; et al. The MR-Base platform supports systematic causal inference across the human phenome. eLife 2018, 7, e34408. [Google Scholar] [CrossRef]
- Watanabe, K.; Taskesen, E.; van Bochoven, A.; Posthuma, D. Functional mapping and annotation of genetic associations with FUMA. Nat. Commun. 2017, 8, 1826. [Google Scholar] [CrossRef]
- Rangwala, S.H.; Kuznetsov, A.; Ananiev, V.; Asztalos, A.; Borodin, E.; Evgeniev, V.; Joukov, V.; Lotov, V.; Pannu, R.; Rudnev, D.; et al. Accessing NCBI data using the NCBI Sequence Viewer and Genome Data Viewer (GDV). Genome Res. 2021, 31, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-H.; Brown, D.W.; Machiela, M.J. LDtrait: An Online Tool for Identifying Published Phenotype Associations in Linkage Disequilibrium. Cancer Res. 2020, 80, 3443–3446. [Google Scholar] [CrossRef] [PubMed]
- Staley, J.R.; Blackshaw, J.; Kamat, M.A.; Ellis, S.; Surendran, P.; Sun, B.B.; Paul, D.S.; Freitag, D.; Burgess, S.; Danesh, J.; et al. PhenoScanner: A database of human genotype–phenotype associations. Bioinformatics 2016, 32, 3207–3209. [Google Scholar] [CrossRef] [PubMed]
- A Kamat, M.; A Blackshaw, J.; Young, R.; Surendran, P.; Burgess, S.; Danesh, J.; Butterworth, A.S.; Staley, J.R. PhenoScanner V2: An expanded tool for searching human genotype–phenotype associations. Bioinformatics 2019, 35, 4851–4853. [Google Scholar] [CrossRef] [PubMed]
- Safran, M.; Rosen, N.; Twik, M.; BarShir, R.; Stein, T.I.; Dahary, D.; Fishilevich, S.; Lancet, D. The GeneCards Suite. In Practical Guide to Life Science Databases; Abugessaisa, I., Kasukawa, T., Eds.; Springer: Singapore, 2021; pp. 27–56. [Google Scholar]
- Stanfill, A.G.; Cao, X. Enhancing Research through the Use of the Genotype-Tissue Expression (GTEx) Database. Biol. Res. Nurs. 2021, 23, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Carithers, L.J.; Ardlie, K.; Barcus, M.; Branton, P.A.; Britton, A.; Buia, S.A.; Compton, C.C.; DeLuca, D.S.; Peter-Demchok, J.; Gelfand, E.T.; et al. A Novel Approach to High-Quality Postmortem Tissue Procurement: The GTEx Project. Biopreservation Biobanking 2015, 13, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Hamosh, A.; Scott, A.F.; Amberger, J.; Valle, D.; McKusick, V.A. Online Mendelian Inheritance in Man (OMIM). Hum. Mutat. 2000, 15, 57–61. [Google Scholar] [CrossRef]
- Gaunt, T.R.; Shihab, H.A.; Hemani, G.; Min, J.L.; Woodward, G.; Lyttleton, O.; Zheng, J.; Duggirala, A.; McArdle, W.L.; Ho, K.; et al. Systematic identification of genetic influences on methylation across the human life course. Genome Biol. 2016, 17, 61. [Google Scholar] [CrossRef]
- Bonder, M.J.; Luijk, R.; Zhernakova, D.; Moed, M.; Deelen, P.; Vermaat, M.; Van Iterson, M.M.; Van Dijk, F.; Van Galen, M.; Bot, J.; et al. Disease variants alter transcription factor levels and methylation of their binding sites. Nat. Genet. 2017, 49, 131–138. [Google Scholar] [CrossRef]
- Lappalainen, T.; Sammeth, M.; Friedländer, M.R.; ‘t Hoen, P.A.C.; Monlong, J.; Rivas, M.A.; Gonzàlez-Porta, M.; Kurbatova, N.; Griebel, T.; Ferreira, P.G.; et al. Transcriptome and genome sequencing uncovers functional variation in humans. Nature 2013, 501, 506–511. [Google Scholar] [CrossRef]
- Aydemir, T.B.; Cousins, R.J. The Multiple Faces of the Metal Transporter ZIP14 (SLC39A14). J. Nutr. 2018, 148, 174–184. [Google Scholar] [CrossRef]
- Tuschl, K.; Meyer, E.; Valdivia, L.E.; Zhao, N.; Dadswell, C.; Abdul-Sada, A.; Hung, C.Y.; Simpson, M.A.; Chong, W.K.; Jacques, T.S.; et al. Mutations in SLC39A14 disrupt manganese homeostasis and cause childhood-onset parkinsonism–dystonia. Nat. Commun. 2016, 7, 11601. [Google Scholar] [CrossRef]
- Xin, Y.; Gao, H.; Wang, J.; Qiang, Y.; Imam, M.U.; Li, Y.; Wang, J.; Zhang, R.; Zhang, H.; Yu, Y.; et al. Manganese transporter Slc39a14 deficiency revealed its key role in maintaining manganese homeostasis in mice. Cell Discov. 2017, 3, 17025. [Google Scholar] [CrossRef] [PubMed]
- Jenkitkasemwong, S.; Akinyode, A.; Paulus, E.; Weiskirchen, R.; Hojyo, S.; Fukada, T.; Giraldo, G.; Schrier, J.; Garcia, A.; Janus, C.; et al. SLC39A14 deficiency alters manganese homeostasis and excretion resulting in brain manganese accumulation and motor deficits in mice. Proc. Natl. Acad. Sci. USA 2018, 115, E1769–E1778. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.; Morgan, H.; Johnson, A.; Nicholson, R. Structure–function analysis of a novel member of the LIV-1 subfamily of zinc transporters, ZIP14. FEBS Lett. 2005, 579, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Liuzzi, J.P.; Aydemir, F.; Nam, H.; Knutson, M.D.; Cousins, R.J. Zip14 (Slc39a14) mediates non-transferrin-bound iron uptake into cells. Proc. Natl. Acad. Sci. USA 2006, 103, 13612–13617. [Google Scholar] [CrossRef]
- Widhalm, R.; Ellinger, I.; Granitzer, S.; Forsthuber, M.; Bajtela, R.; Gelles, K.; Hartig, P.-Y.; Hengstschläger, M.; Zeisler, H.; Salzer, H.; et al. Human placental cell line HTR-8/SVneo accumulates cadmium by divalent metal transporters DMT1 and ZIP14. Metallomics 2020, 12, 1822–1833. [Google Scholar] [CrossRef]
- Boczonadi, V.; King, M.S.; Smith, A.C.; Olahova, M.; Bansagi, B.; Roos, A.; Eyassu, F.; Borchers, C.; Ramesh, V.; Lochmüller, H.; et al. Mitochondrial oxodicarboxylate carrier deficiency is associated with mitochondrial DNA depletion and spinal muscular atrophy–like disease. Anesthesia Analg. 2018, 20, 1224–1235. [Google Scholar] [CrossRef]
- Wahlberg, K.; Love, T.M.; Pineda, D.; Engström, K.; Watson, G.E.; Thurston, S.W.; Yeates, A.J.; Mulhern, M.S.; McSorley, E.M.; Strain, J.; et al. Maternal polymorphisms in glutathione-related genes are associated with maternal mercury concentrations and early child neurodevelopment in a population with a fish-rich diet. Environ. Int. 2018, 115, 142–149. [Google Scholar] [CrossRef]
- de Oliveira, A.S.; de Souza, M.F.; Lengert, A.v.H.; de Oliveira, M.T.; Camargo, R.B.d.O.G.; Braga, G.L.; Cólus, I.M.d.S.; Barbosa, F., Jr.; Barcelos, G.R.M. Genetic Polymorphisms in Glutathione (GSH-) Related Genes Affect the Plasmatic Hg/Whole Blood Hg Partitioning and the Distribution between Inorganic and Methylmercury Levels in Plasma Collected from a Fish-Eating Population. BioMed Res. Int. 2014, 2014, 940952. [Google Scholar] [CrossRef]
- Buck, K.A.; Varian-Ramos, C.W.; Cristol, D.A.; Swaddle, J.P. Blood Mercury Levels of Zebra Finches Are Heritable: Implications for the Evolution of Mercury Resistance. PLoS ONE 2016, 11, e0162440. [Google Scholar] [CrossRef]
- Paape, T.; Heiniger, B.; Domingo, M.S.; Clear, M.R.; Lucas, M.M.; Pueyo, J.J. Genome-Wide Association Study Reveals Complex Genetic Architecture of Cadmium and Mercury Accumulation and Tolerance Traits in Medicago truncatula. Front. Plant Sci. 2022, 12, 806949. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, K.; Yasutake, A. Sex and age differences in mercury distribution and excretion in methylmercury-administered mice. J. Toxicol. Environ. Health 1986, 18, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.T. The influence of age on the gastrointestinal absorption of mercuric chloride and methyl mercury chloride in the rat. Environ. Res. 1982, 27, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Eide, D.J. The SLC39 family of zinc transporters. Mol. Asp. Med. 2013, 34, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.M.; Dnyanmote, A.V.; Bush, K.T.; Wu, W.; Nigam, S.K. Deletion of Multispecific Organic Anion Transporter Oat1/Slc22a6 Protects against Mercury-induced Kidney Injury. J. Biol. Chem. 2011, 286, 26391–26395. [Google Scholar] [CrossRef] [PubMed]
- Borges, V.C.; Santos, F.W.; Rocha, J.B.T.; Nogueira, C.W. Heavy Metals Modulate Glutamatergic System in Human Platelets. Neurochem. Res. 2007, 32, 953–958. [Google Scholar] [CrossRef]
- Albrecht, J.; Matyja, E. Glutamate: A potential mediator of inorganic mercury neurotoxicity. Metab. Brain Dis. 1996, 11, 175–184. [Google Scholar] [CrossRef]
- Peixoto, N.; Serafim, A.; Flores, E.; Bebianno, M.; Pereira, M. Metallothionein, zinc, and mercury levels in tissues of young rats exposed to zinc and subsequently to mercury. Life Sci. 2007, 81, 1264–1271. [Google Scholar] [CrossRef]
- Lozano, M.; Yousefi, P.; Broberg, K.; Soler-Blasco, R.; Miyashita, C.; Pesce, G.; Kim, W.J.; Rahman, M.; Bakulski, K.M.; Haug, L.S.; et al. DNA methylation changes associated with prenatal mercury exposure: A meta-analysis of prospective cohort studies from PACE consortium. Environ. Res. 2022, 204, 112093. [Google Scholar] [CrossRef]
- Pierce, B.L.; Kibriya, M.G.; Tong, L.; Jasmine, F.; Argos, M.; Roy, S.; Paul-Brutus, R.; Rahaman, R.; Rakibuz-Zaman, M.; Parvez, F.; et al. Genome-Wide Association Study Identifies Chromosome 10q24.32 Variants Associated with Arsenic Metabolism and Toxicity Phenotypes in Bangladesh. PLoS Genet. 2012, 8, e1002522. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, M.F.H.; Grotto, D.; Barbosa, F., Jr. Inorganic and Methylmercury Levels in Plasma are Differentially Associated with Age, Gender, and Oxidative Stress Markers in a Population Exposed to Mercury through Fish Consumption. J. Toxicol. Environ. Health Part A 2014, 77, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-G.; Jo, E.-M.; Kim, G.-Y.; Kim, D.-S.; Kim, Y.-M.; Kim, R.-B.; Suh, B.-S.; Hong, Y.-S. Analysis of Methylmercury Concentration in the Blood of Koreans by Using Cold Vapor Atomic Fluorescence Spectrophotometry. Ann. Lab. Med. 2012, 32, 31–37. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| The Avon Longitudinal Study of Parents and Children | The Human Early Life Exposome | |
|---|---|---|
| Abbreviation | ALSPAC | HELIX |
| Date of recruitment | April 1991–December 1992 | 1999–2010 |
| Population | Pregnant women | Children aged 7 to 9 |
| Location | Former Avon Health Authority area, UK | UK, France, Spain, Lithuania, Norway, and Greece |
| Total participants | 14,833 | Approximately 32,000 |
| Hg source | Whole blood | Whole blood |
| Hg sample timing | Early pregnancy | 6–11 years old |
| Hg analysis method | ICP-DRC-MS | ICP-SFMS |
| Genotyping method | Illumina Human660W-Quad Array | Infinium Global Screening Array (GSA) (Illumina) |
| Imputation panel | Haplotype Reference Consortium (HRC r1.1) | Haplotype Reference Consortium (HRC r1.1) |
| Country | Sample Size | |
|---|---|---|
| Children (HELIX) | - | 1042 |
| Born in Bradford (BiB) | United Kingdom | 90 |
| Étude des Déterminants pré et postnatals du développement et de la santé de l’ENfant (EDEN) | France | 135 |
| INfancia y Medio Ambiente (INMA) | Spain | 198 |
| Kaunas birth cohort (KANC) | Lithuania | 196 |
| The Norwegian Mother, Father and Child cohort study (MoBa) | Norway | 237 |
| The Rhea Mother–Child Cohort in Crete (Rhea) | Greece | 186 |
| Pregnant women (ALSPAC) | United Kingdom | 2983 |
| ALSPAC | HELIX | |||
|---|---|---|---|---|
| SNP | N | SNP | N | |
| Direct genotyped | 557,124 | 10,015 | 692,367 | 1397 |
| Direct genotyped after QC 1 | 526,688 | 8196 | 509,344 | 1304 |
| Imputed (HRC r1.1) 2 | 39,117,141 | 8196 | 40,405,505 | 1304 |
| Imputed after QC | 6,649,782 | 8196 | 6,143,757 | 1304 |
| Blood Hg measurements | - | 4014 | - | 1301 |
| GWAS | 6,649,782 | 2893 | 6,143,757 | 1042 |
| GWAS after QC | 6,620,135 | 2893 | 6,138,843 | 1042 |
| Pregnant Women, ALSPAC (n = 2893, nSNP = 6,620,135) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SNP | Chr | Position 1 | Gene 2 | Effect Allele | Other Allele | MAF 3 | β | SE 4 | p-Value | HELIX p-Value 5 |
| rs4853739 | 2 | 191698516 | - | T | C | 0.24 | 0.10 | 0.02 | 2.32 × 10−6 | - |
| rs11709754 | 3 | 149615531 | RNF13 | T | A | 0.21 | −0.10 | 0.02 | 9.39 × 10−6 | - |
| rs361166 | 4 | 152792791 | - | A | G | 0.41 | 0.09 | 0.02 | 5.78 × 10−6 | 0.53 |
| rs6859392 | 5 | 63241481 | - | G | C | 0.30 | −0.09 | 0.02 | 3.31 × 10−6 | - |
| rs1372504 | 5 | 103749428 | RP11-6N13.1 | A | G | 0.38 | −0.09 | 0.02 | 1.70 × 10−6 | 0.53 |
| rs2246509 | 6 | 156058999 | - | G | A | 0.46 | 0.08 | 0.02 | 8.69 × 10−6 | 0.98 |
| rs1845418 | 7 | 24095415 | - | T | C | 0.14 | 0.14 | 0.03 | 8.68 × 10−8 | 0.48 |
| rs146099921 | 8 | 22254947 | SLC39A14 | G | T | 0.14 | −0.12 | 0.03 | 8.21 × 10−6 | 0.14 |
| rs7900717 | 10 | 122034207 | - | C | T | 0.25 | −0.09 | 0.02 | 6.02 × 10−6 | 0.31 |
| rs7301395 | 12 | 116402976 | MED13L | A | G | 0.02 | 0.33 | 0.07 | 5.06 × 10−6 | - |
| rs12874443 | 13 | 38304884 | TRPC4 | T | G | 0.45 | −0.08 | 0.02 | 7.73 × 10−6 | 0.99 |
| rs113202356 | 16 | 27891845 | GSG1L | A | G | 0.10 | −0.14 | 0.03 | 8.45 × 10−6 | - |
| rs74450576 | 16 | 73918519 | - | A | G | 0.04 | 0.21 | 0.05 | 7.40 × 10−6 | 0.80 |
| rs11643897 | 16 | 78221370 | WWOX | C | T | 0.32 | 0.08 | 0.02 | 9.70 × 10−6 | 0.87 |
| rs35522803 | 19 | 3592734 | GIPC3 | T | C | 0.08 | −0.15 | 0.03 | 3.83 × 10−6 | 0.82 |
| rs60192794 | 20 | 52317186 | - | C | T | 0.07 | −0.16 | 0.04 | 5.66 × 10−6 | - |
| Children, HELIX (n = 1042, nSNP = 6,138,843) | ||||||||||
| SNP | Chr | Position | Gene | Effect allele | Other allele | MAF | β | SE | p-value | ALSPAC p-value |
| rs7526817 | 1 | 28195486 | - | A | T | 0.09 | −0.67 | 0.15 | 5.02 × 10−6 | 0.06 |
| rs79810835 | 1 | 146999434 | - | A | G | 0.02 | 1.37 | 0.31 | 6.83 × 10−6 | - |
| rs59436870 | 2 | 101830050 | TBC1D8 | T | C | 0.18 | −0.52 | 0.11 | 5.16 × 10−6 | 0.74 |
| rs9852537 | 3 | 98658085 | CTD-2021J15.1 | T | C | 0.03 | 1.29 | 0.29 | 7.62 × 10−6 | 0.66 |
| rs186276942 | 3 | 115279279 | - | G | A | 0.01 | 2.87 | 0.63 | 5.29 × 10−6 | - |
| rs62287513 | 3 | 184104050 | CHRD | A | G | 0.31 | 0.44 | 0.09 | 7.69 × 10−7 | - |
| rs28618224 | 4 | 21041710 | KCNIP4 | C | A | 0.03 | 1.47 | 0.32 | 5.03 × 10−6 | 0.11 |
| rs115812569 | 4 | 42726300 | - | G | A | 0.01 | 1.82 | 0.40 | 6.42 × 10−6 | 0.56 |
| rs2904271 | 4 | 90284516 | - | C | A | 0.37 | −0.41 | 0.09 | 1.84 × 10−6 | 0.79 |
| rs113384484 | 5 | 151015719 | - | A | G | 0.04 | 1.03 | 0.22 | 1.50 × 10−6 | 0.02 |
| rs79340261 | 7 | 35559836 | - | A | C | 0.02 | 1.93 | 0.40 | 1.08 × 10−6 | - |
| rs75847252 | 9 | 71483481 | PIP5K1B | C | T | 0.02 | 1.22 | 0.26 | 2.96 × 10−6 | - |
| rs73510541 | 11 | 69313573 | - | A | G | 0.13 | 0.54 | 0.12 | 3.72 × 10−6 | 0.50 |
| rs9510838 | 13 | 24317031 | - | T | G | 0.17 | 0.58 | 0.12 | 3.43 × 10−6 | 0.88 |
| rs9563673 | 13 | 34239526 | RP11-141M1.3 | C | T | 0.07 | −0.90 | 0.19 | 5.90 × 10−6 | 0.21 |
| rs17106291 | 14 | 37559401 | SLC25A21 | G | A | 0.02 | 2.32 | 0.52 | 7.53 × 10−6 | - |
| rs7154700 | 14 | 63201771 | KCNH5 | T | C | 0.01 | 1.61 | 0.34 | 2.15 × 10−6 | - |
| rs116971963 | 17 | 14961821 | - | A | G | 0.01 | 1.57 | 0.34 | 4.91 × 10−6 | - |
| rs145982353 | 19 | 54257385 | - | G | C | 0.03 | 1.27 | 0.27 | 4.07 × 10−6 | - |
| rs6075980 | 20 | 288233 | - | G | A | 0.38 | 0.42 | 0.09 | 2.31 × 10−6 | 0.24 |
| rs148653405 | 20 | 5594216 | - | A | G | 0.01 | 1.77 | 0.40 | 8.35 × 10−6 | 0.34 |
| SNP | Gene | Source Study, Direction of Effect, and Biological Matrix | Effect Allele | Other Allele | Study | β | SE | p-Value |
|---|---|---|---|---|---|---|---|---|
| Hg—gene association studies | ||||||||
| rs3811647 | TF | A allele, lower cord tissue Hg [48] | A | G | ALSPAC | 0.00 | 0.02 | 0.83 |
| HELIX | −0.14 | 0.09 | 0.11 | |||||
| rs761142 | GCLC | GG vs. TT, lower hair Hg [107] | C | A | ALSPAC | −0.01 | 0.02 | 0.63 |
| HELIX | 0.06 | 0.10 | 0.57 | |||||
| rs41307970 | GCLM | CT + TT vs. TT, lower blood Hg [108] | C | G | ALSPAC | 0.00 | 0.05 | 0.93 |
| HELIX | 0.20 | 0.25 | 0.41 | |||||
| rs10636 | MT2A | C allele, higher urine Hg [46] | C | G | ALSPAC | 0.05 | 0.02 | 0.01 |
| HELIX | −0.06 | 0.10 | 0.53 | |||||
| rs2270837 1 | MT1M | A allele, higher urine Hg [46] | G | A | ALSPAC | 0.01 | 0.03 | 0.57 |
| rs8052394 | MT1A | A allele interacts with fish intake for higher hair Hg [46] | G | A | ALSPAC | 0.00 | 0.03 | 0.96 |
| HELIX | 0.05 | 0.13 | 0.69 | |||||
| rs9936741 | MT1M | T allele interacts with fish intake for higher hair Hg [46] | C | T | ALSPAC | −0.08 | 0.06 | 0.17 |
| HELIX | −0.95 | 0.37 | 0.01 | |||||
| Lead—GWAS | ||||||||
| rs1805313 | ALAD | G allele, lower blood lead [54] | G | A | ALSPAC | 0.01 | 0.02 | 0.57 |
| HELIX | 0.10 | 0.09 | 0.25 | |||||
| Selenium—GWAS | ||||||||
| rs672413 | ARSB | G allele, lower blood selenium [56] | G | A | ALSPAC | 0.00 | 0.02 | 0.92 |
| HELIX | −0.01 | 0.09 | 0.95 | |||||
| rs705415 1 | Intergenic | T allele, lower blood selenium [56] | T | C | HELIX | 0.15 | 0.17 | 0.37 |
| rs6586282 | CBS | T allele, lower blood selenium [56] | T | C | ALSPAC | −0.03 | 0.02 | 0.27 |
| HELIX | −0.06 | 0.11 | 0.61 | |||||
| Zinc—GWAS | ||||||||
| rs2120019 | PPCDC | C allele, lower blood zinc [55] | C | T | ALSPAC | 0.00 | 0.02 | 0.94 |
| HELIX | 0.07 | 0.10 | 0.50 | |||||
| rs1532423 | CA1 | G allele, lower blood zinc [55] | G | A | ALSPAC | −0.03 | 0.02 | 0.07 |
| HELIX | −0.08 | 0.09 | 0.36 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dack, K.; Bustamante, M.; Taylor, C.M.; Llop, S.; Lozano, M.; Yousefi, P.; Gražulevičienė, R.; Gutzkow, K.B.; Brantsæter, A.L.; Mason, D.; et al. Genome-Wide Association Study of Blood Mercury in European Pregnant Women and Children. Genes 2023, 14, 2123. https://doi.org/10.3390/genes14122123
Dack K, Bustamante M, Taylor CM, Llop S, Lozano M, Yousefi P, Gražulevičienė R, Gutzkow KB, Brantsæter AL, Mason D, et al. Genome-Wide Association Study of Blood Mercury in European Pregnant Women and Children. Genes. 2023; 14(12):2123. https://doi.org/10.3390/genes14122123
Chicago/Turabian StyleDack, Kyle, Mariona Bustamante, Caroline M. Taylor, Sabrina Llop, Manuel Lozano, Paul Yousefi, Regina Gražulevičienė, Kristine Bjerve Gutzkow, Anne Lise Brantsæter, Dan Mason, and et al. 2023. "Genome-Wide Association Study of Blood Mercury in European Pregnant Women and Children" Genes 14, no. 12: 2123. https://doi.org/10.3390/genes14122123
APA StyleDack, K., Bustamante, M., Taylor, C. M., Llop, S., Lozano, M., Yousefi, P., Gražulevičienė, R., Gutzkow, K. B., Brantsæter, A. L., Mason, D., Escaramís, G., & Lewis, S. J. (2023). Genome-Wide Association Study of Blood Mercury in European Pregnant Women and Children. Genes, 14(12), 2123. https://doi.org/10.3390/genes14122123