Abstract
The green peach aphid (GPA), Myzus persicae, is an important pest of the peach crop. Three major dominant resistance genes have already been detected, Rm1 in the Weeping Flower Peach (WFP) clone, Rm2 in the Rubira clone, and Rm3 in the Fen Shouxing clone. In this study, after NGS resequencing of WFP and Rubira, we found that their genomic sequences in the Rm1 and Rm2 region were similar but very different from that of the susceptible reference peach Lovell. We constructed a BAC library for the GPA-resistant WFP and screened four BAC clones to sequence the target region. The new sequence was 61.7 Kb longer than Lovell and was annotated with four different TIR_NBS_LRR genes. Among them, the TNL1 gene was very overexpressed in WFP leaves 24 h after GPA infestation. This gene was also present and expressed in the Rubira clone and had the same sequence as the candidate Rm3 gene, supporting the hypothesis that the three genes share the same origin. In addition, we identified a second TNL, TNL2, located at 35.4 Kb from TNL1 and slightly overexpressed after GPA infestation. Kasp and size molecular markers were designed for use in marker-assisted selection and were validated in a peach segregating population.
1. Introduction
The identification of resistance genes to pests and pathogens in peach (Prunus persica L.) and other Prunus species is one of the main objectives of developing resistant peach and rootstock cultivars. Several mapping studies have been developed for locating Prunus resistance genes to pathogens [1,2] and only a few have been identified and functionally validated, such as the resistance genes to Root-knot nematodes Ma [3] and RMja [4] and the resistance gene to powdery mildew Vr3 [5]. Now, with the development of reference genomes and new sequencing technologies (NGS, long read RNAseq, Pacbio), there are powerful tools to carry out detailed molecular studies leading to the identification of new genes and genetic markers. In peach, the peach reference genome V1 [6], available in 2010, was generated from the double haploid peach “Lovell” PLov2-2n, and a second version of the peach genome (Peach v2.0) [7] from the same genotype Lovell was achieved in 2016 (https://www.rosaceae.org/species/prunus_persica/genome_v2.0.a1 accessed on 1 June 2020). It allowed us to continue the research on the resistance genes to the green peach aphids (GPA), a major pest of peach causing leaf curling, stunted shoots, and weakening of the tree. In 1982, Massonié et al. [8] developed inoculation tests and identified two varieties, the peach Weeping Flower Peach (WFP) S2678 and the peach rootstock Rubira S4359, where aphid colonies did not develop and where resistance was associated with a necrotic reaction. The WFP S2678 clone was introduced to INRAE in 1961 from Clemson University with no information on its origin. It has red double flowers, a weeping growth habit, and is also resistant to the aphid M. varians Davids [9]. Rubira is a red leaf peach seedling rootstock released by INRAE from a Californian peach seed lot.
Subsequently, several mapping studies were conducted by INRAE teams to identify the resistance genes of these two varieties, naming Rm1 the gene present in the WFP clone [10,11] and Rm2 the gene present in Rubira [12] and showing segregation of a single dominant gene.
Notably, the WFP Rm1 resistance gene to aphids causes strong antixenosis resistance [13]. This mechanism of resistance triggers hypersensitive-like necrotic reactions and protects the plant from aphid colonization. Based on this resistance mechanism, it can be expected that candidate genes belong to the family of the NLR (nucleotide-binding site-leucine-rich repeat). More recently, in China, a third single dominant GPA resistant gene, Rm3, was localized in the Chinese clone Fen Shouxing [14], and an NLR gene was identified as a candidate gene for GPA resistance [15]. As the two genes, Rm2 and Rm3, trigger the same reaction as Rm1 [14]), we hypothesized that these three genes probably belong to the same locus, but this hypothesis remains to be confirmed.
Rough mapping placed the three genes at the end of chromosome 1, the Rm1 locus in the genomic region (Pp01:43.62~46.50 Mb) [11], the Rm2 locus in the genomic region (Pp01:45.08~46.23 Mb) [16], and the Rm3 locus [17] within a 460-kb interval between the two SNP M1 SNP_Pp01_45.665389 and M2 SNP_Pp01_46.120950 (Figure 1a). In the hypothesis of a common origin and similarity of the three genes, the Rm1 and Rm2 genes should also be localized at the Rm3 locus in the interval between the two markers M1 and M2, which is the smallest and is include in the genomic region encompassing the two other genes.
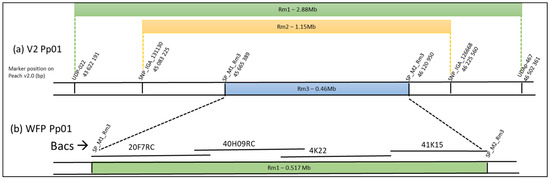
Figure 1.
(a) Schemes of the genomic region identified by fine mapping for the Rm1 [11], Rm2 [16], and Rm3 [17] genes. The position on the chromosome Pp01 peach V2 of the SSR and SNP markers framing the region are indicated. (b) Position of the four sequenced BAC clones screened from WFP leaves DNA.
In this M1M2 interval, 53 genes (Prupe.1G559400 to Prupe.1G564500) were annotated in the peach V2 reference genome and none of them as defense genes [9]. Four additional TNL genes (ppa000596m, ppa017041m, ppa015956m and ppa018622m) annotated on the peach V1.0 reference in the corresponding interval were reported on the Peach V2 by Van Ghelder and Esmenjaud [12] in a specific track on the GDR Jbrowse of the Peach V2 including all annotated TNL genes in peach.
In this work, we decided to resequence the genomics region, including the M1M2 interval of the resistant genotypes WFP and Rubira, which harbor Rm1 and Rm2, respectively, in order to look for additional TNL genes that may not be present in the peach reference V2. Using resequencing and expression data in WFP and Rubira, we identified four TNL genes in the targeted interval, two of which are overexpressed in aphid-infected plants.
2. Materials and Methods
2.1. Plant Materials
The trees of the two aphid-resistant peach clones, S2678 Weeping Flower Peach and S4359 Rubira, were maintained in the INRAE collection in 32l pots under insect-proof tunnels. The plants used for the GPA infestation tests were produced by grafting on the peach GF305 rootstock [8]. In this study, a population of 108 hybrids (Pamirskii × Rubira) × (Montclar × Nemared) PRMN previously described [18] were phenotyped for resistance to M. persicae and genotyped with specific molecular markers for the validation of the candidate genes. Thirty French peach accessions of the INRAE peach collections known as susceptible were genotyped with Kasp-specific markers (Table S1).
2.2. Phenotyping for Aphid Resistance
The test was carried out in a greenhouse under controlled conditions on young plants grown in small pots. After 3 months of growth, seedlings or grafted plants were infested by placing three adult apterous M. persicae on the terminal apex of each plant. The adult aphids were maintained and provided by the INRAE unit PSH (Plant and Horticultural System), which collected the aphid genotype from a peach tree in Avignon [13]. We observed the plants two weeks after infestation. Two classes of symptoms were observed and recorded. S: development of an aphid colony and leaf curling reactions, R: no aphid colony development, escape of aphids, and presence of necrotic spots (Figure 2).

Figure 2.
(a) Phenotype of the WFP resistant plant after infestation with GPA with the escape of aphids and presence of necrotic spots. (b) GPA colony development on a susceptible hybrid.
2.3. Nucleic Acid Extraction and Genotyping
The genomic DNA of all tested plants was extracted using 100 mg of frozen young leaves. Each sample was ground with a mixer mill in 330 µL of extraction buffer (sorbitol 0.35 M, Tris 0.1 M, EDTA 5 mM, 4 mg of sodium metabisulfite), 330 µL of lysis buffer (Tris 0.2 M, EDTA 50 mM, NaCl 2 M, CTAB 2%), and 130 µL of sarkosyl 5%. After a chloroform-isoamylalcohol (24:1) procedure, precipitation with isopropanol and three ethanol washes, the DNA content was eluted in water and treated with RNAse. PCRs were carried out using the MyTaq DNA polymerase kit or the Expand long-range kit for longer fragments, following the manufacturer’s instructions.
2.4. NGS Genome Sequencing of the Peach Genotypes
The two resistant peaches, WFP and Rubira, and the parents of the PRMN population, Pamirskij, Montclar, and Nemared, were resequenced. The DNA was extracted from the leaves of the trees and maintained in pots under insect-proof tunnels. NGS resequencing was achieved using the platform MGX-Montpellier. Tagmentation and PCR amplifications were carried out using the Illumina Nextera DNA sample preparation kit following the supplier’s instructions. The validation of the genomics libraries was performed with DNA quantification using an Agilent high sensitivity chip and by qPCR. The cluster was carried out inside a flow cell using the Illumina cluster generation kit. Then, paired-end 125 sequencing was achieved using an Illumina Hiseq 2500 with a sequence by synthesis (SBS) technique. Pre-treatment of the data was performed using Illumina software HCS and RTA. The quality of the samples was analyzed with the software FastQC.
2.5. Genomic Library and Bacterial Artificial Chromosome (BAC) Sequencing
High molecular weight (HMW) genomic DNA was prepared from young frozen leaves picked from a potted tree kept in the dark for a week from the resistant peach WFP as described by Peterson [19] and Gonthier [20]. Agarose-embedded HMW DNA was partially digested with HindIII and subjected to two size selection steps by pulsed-field electrophoresis using a CHEF Mapper system. DNA was eluted, ligated into the pIndigoBAC-5 HindIII-Cloning Ready vector, and transformed into Escherichia coli electrocompetent cells. Pulsed-field migration programs electrophoresis buffer and ligation desalting conditions were performed based on the protocol of Challoub [21]. BAC clones were spotted on a nylon membrane, screened with radioactively labelled probes, and revealed by the high-density filter reader program. Eight labelled probes were designed using Primer 3 software from parts of sequences of WFP conserved with Peach V2 in the targeted genomic interval (Table S1).
We verified the positive clones by real-time PCR using specific primers. In total, 2 µg of each individual BAC clone of interest was pooled for the construction of a SMRT® library using the standard Pacific Biosciences preparation protocol for a 10 kb library with PacBio® Barcoded Adapters.
The pool was then sequenced in one SMRT cell using P6 polymerase with C4 chemistry. Sequencing was performed on the PacBio RS II sequencer. After a demultiplexing step, the sequence assembly was performed following the HGAP PacBio workflow [22] and using SMRT® Analysis software version 2.3 October 2014 for HGAP implementation BAC end sequences confirmed the position of selected clones on the peach genome.
2.6. Construction and Sequencing Full-Length RNA-Seq Library
RNA extractions: We extracted total RNA from 2 apex leaf samples and 2 root samples using the NucleoSpin® RNA Plant kit (Macherey Nagel). RNA concentration was measured using a Nanodrop® by spectrophotometric absorbance at 260 and 280 nm. The quality of total RNA was evaluated using an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). Only samples with RNA integrity number (RIN) ≥ 8 were used for deep sequencing.
Library construction and RNA sequencing: The complementary DNA was prepared from 500 ng of total RNA using the NEB Next® Single Cell/Low Input cDNA Synthesis and Amplification Module kit. We used the SMRTbell® Express Template Prep Kit 2.0 to prepare libraries and multiplex up to 2 samples. PacBio Sequel® Systems was used to sequence transcriptomes. Full-length reads were selected and trimmed of 5′ and 3′ primers with specific adaptors (CGCACTCTGATATGTG for reverse primer of RNA apex, CTCACAGTCTGTGTGT for reverse primer of RNA roots).
The raw SMRT sequencing data for this work was mapped onto the Peach V2 reference sequence, to which a new scaffold corresponding to the de novo sequences from the previously produced BAC libraries was added. We used the Iso-Seq 3 bioinformatics workflow in SMRT Link Version 7.0.1.66975 to produce full-length transcript sequences and splice variants of good quality, with no assembly required. The Iso-Seq analysis workflow starts with the generation of high-fidelity reads using the circular consensus sequencing (CCS) method and the required trimmed poly(A) tail. The minimum accepted subread length was 50 bp, and we added the isoseq-mode option.
2.7. Gene Expression (qPCR)
Total RNA from apex leaves of WFP genotypes, from aphid-infected after 24 h and from healthy plants was extracted from frozen samples using the NucleoSpin®RNA Plant Kit (Macherey-Nagel, Dueren, Germany) according to the manufacturer’s instructions. RNA yield and purity were quantified using a Nanodrop® by spectrophotometric absorbance and agarose gel electrophoresis. In total, 500 mg of cDNAs were synthesized using the Affinity Script Multiple Temperature cDNA Synthesis Kit (Agilent, Santa Clara, CA, USA), following the instructions provided. Expression analysis was performed by quantitative real-time PCR (qRT-PCR) using a “Stratagen MX Pro 3005” and Brilliant III SYBR Green qPCR Master Mix with Low ROX (Agilent Technologies). The reaction was prepared with each primer (0.5 µL, 10 µM), 2 µL of 1:10 diluted cDNA, RNase-free water (5 µL), and Brilliant III SYBR Green qPCR Master Mix with High ROX (7 µL) in a total volume of 15 µL. PCR conditions were 95 °C for 10 min, followed by 40 cycles of 95 °C for 30 s and 60 °C for 1 min. All qPCR reactions were normalized using the Ct value corresponding to the EiF4Ɣ peach gene (Pp_Eif4Ɣ) and the 60 S L13 ribosomal protein gene (Pp_RPL13). Each sample was measured three times with three technical replicates. The list of the designed primers for the qPCR is in Table S1.
2.8. NGS and Gene Sequence Analysis
The sequence alignment on the peach genome V2 was obtained using BWA (v 0.7.12-r1039) with the BWA-MEM algorithm for mapping. The read alignment and index files (BAM and BAI files), genomic feature files (GFF3) on the reference Peach V2, and de novo sequences were visualized with the Integrative Genomics Viewer (IGV) [23]. We used the CLC workbench (Qiagen, Hilden, Germany) for the sequence trimmings, translation proteins, gene annotations, and motif research. The software used for the gene prediction was FGENESH [24], with Pp as organism-specific gene-finding parameters (http://www.softberry.com) (accessed on 1 June 2020). Protein analysis and domain research was carried out with Interproscan (http://www.ebi.ac.uk/interpro/) (accessed on 1 June 2020).
3. Results
3.1. Reconstruction of the Genomic Region Encompassing Rm1 in WFP
The Rm1 gene was previously located between the markers UDP022 and UDPAP467 and the Rm3 gene between the markers M1 SNP_Pp01:45.665389 and M2 SNP_Pp01_46.120950. To obtain the genomic sequence of WFP in this interval we resequenced WFP on an Illumina Hiseq 2500. We obtained 63.6 million short reads after including purifying filters for a depth of 42×. Only 2.4% of WFP reads were unmapped on the Peach V2. In the targeted M1M2 interval, there was deficient mapping to allow reconstruction of the complete WFP sequence in this interval. Indeed, in this interval, there were some parts of sequences conserved between WFP and Lovell, some dissimilar sequences detected when no WFP short-reads could align on the Lovell sequence and then the impossibility to reconstruct the WFP sequence.
To construct a physical map of the dissimilar region, we built a BAC library of the WFP accession with the aim of sequencing some BAC clones of this chromosome region. Our library contained 18,432 clones with an average insertion size of 119 kb and genome coverage of 7.6×. We screened 27 BAC clones, including the Rm1 regions, with the eight specific probes designed from the conserved sequences in the M1M2 interval. After aligning all the BAC ends on the Peach V2, we selected four BACS (20F07, 40H09, 4K22, and 41K15. See Figure 1b) and sequenced them using PacBio long-read sequencing. These BAC sequences covering the Rm1 region meet the quality criteria with a quality value of QV > 48, meaning high confidence in nucleotides. The fasta files were assembled by overlapping regions between the four BACs. With the mapping of the WFP NGS short-reads on the four BAC sequences, the few deletions or insertions in the overlapping BACs could be elucidated and resolved, and we obtained, between the two flanking markers M1 and M2, one unique consensus sequence “Rm1bac” of 517.362 kb (fasta File in supplemental data). This sequence from WFP is 61.7 kb longer compared to the same sequence interval in the Lovell peach genome that was sequenced in the reference V2 (455.560 Kb), suggesting it could include additional genes. We decided to look for potential resistance genes in this locus.
3.2. Structural Annotation of the TNL Genes in the Novel Rm1Bac Sequence from WFP
In Peach V2, 53 genes (Prupe.1G559400 to Prupe.1G564500) and 4 TNL genes (ppa000596m, ppa017041m, ppa015956m and ppa018622m) were annotated in the 460Kb M1M2 region (Figure 3a). To look for additional genes in the Rm1bac sequence, we used the predictive tool FGENESH and the prediction revealed 116 genes with 6 TNL genes, positioned as genes 17, 24, 56, 60, 92, and 93 (Figure 3b and Figure S1). Those TNL were compared with the four TNL annotated on the peach V2. TNL1 and TNL2 have 83.31 and 67.96 % nucleic identity with ppa000596m, TNL3 92.41% nucleic identity with ppa017041m, TNL4 99.49% nucleic identity with ppa015956m, TNL5 and TNL6 79.82%, and 80.66% nucleic identity with ppa018622m. TNL1, TNL2, TNL5, and TNL6 are, in a negative sense as the ppa000596 and ppa018622m genes, while TNL3, TNL4, ppa01704m1, and ppa015956m are positive genes.
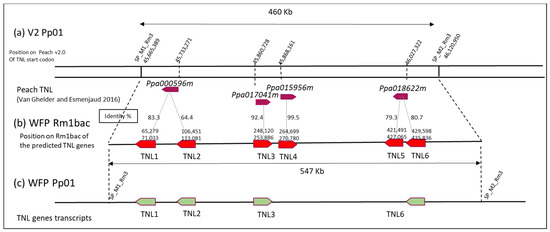
Figure 3.
TNL genes on the Pp01 M1M2 interval: (a) on Peach V2.0 with the 4 TNL genes located by Van Ghelder and Esmenjaud [25]; (b) FGENESH prediction of TNL genes on the WFP genomic sequence (c) Expressed transcripts identified in the WFP long-read RNAseq.
We can conclude that TNL3 close to ppa017041m and TNL4 identical to ppa015956m are quite conserved in both genomes, while TNL1 and especially TNL2 are different from ppa000596m. TNL5 and TNL6, different from ppa018622m, are less conserved even if they are in the same sense. Thus, we can determine that the four genes TNL1, TNL2, TNL5, and TNL6, which are present in the resistant clone WFP and not in the susceptible clone Lovel, could have a role in aphid resistance.
To see if these TNL genes were expressed and to validate their predicted intron/exon structure, we carried out a long-read RNAseq with iso-seq and PacBio technology.
RNAseq data were mapped on the Rm1bac genomic sequence showing that only four TNL genes were expressed, TNL1 (gene17), TNL2 (gene24), TNL3 (gene56), and TNL6 (gene93) (Figure 3c and Figure S3). The structure of the expressed genes, including the number and location of exons, number and location of introns, CDS region, and location of promoter and terminal regions (PolyA), was exactly the same as the FGENESH prediction for the TNL3 gene but differed slightly for the three other genes in exon five for TNL1, and in the exon seven for TNL2 and TNL6. The structure of the gene TNL1 consisted of a sequence of 5755 bp with six exons and five introns, while the three other genes had seven exons and six introns and are, respectively, 6630, 4512, and 4518 bp long. In the long-read transcripts of the four genes, we found a single open reading frame encoding four proteins of 1497, 1768, 1504, and 1506 amino acid residues (Figure 4).
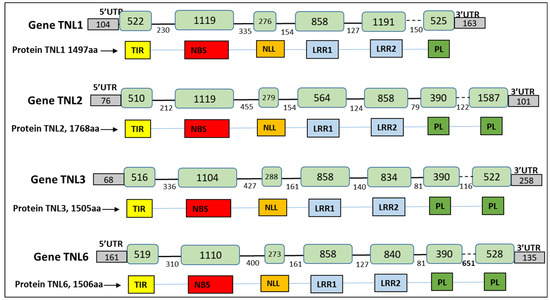
Figure 4.
Structure of the four genes TNL1, TNL2, TNL3, and TNL6: size (bp) of exons, introns, and UTR, predicted domains of the TNL protein. Exons are represented in green boxes, introns by a black lane. The last intron in the dotted line contains one SSR (single sequence repeat).
Using Interproscan analysis of the protein sequences, we confirmed that the four genes belong to the family of TIR-NLR (TNL) genes with the presence of a Toll-interleukin-1 receptor domain in the N-terminal part of the protein and display a complete structure with full-length domains. The three first exons correspond to a specific protein domain. The first exon contains the typical TIR Domain motifs described for Arabidopsis by Meyers [26], TIR1, TIR3, and TIR5; the second exon contains the NB domain motifs P-loop, GLPL, and MHDV, and the third exon contains the motifs NLL1, LRR1a, and PL-1 described by Van Ghelder and Esmenjaud [25] (Figures S3 and S4). The four other exons contain LRR and post-LRR (Leucine-Rich Repeat) domains, but with one less post-LRR exon for TNL1.
Our results showed that four TNL genes are expressed, and among them, three genes, TNL1, TNL2, and TNL6, are very different from the four TNL of the susceptible reference peach Lovell. In order to search which genes are involved in the resistance, we looked to see if they were overexpressed in GPA-infested plants, in the hypothesis of overexpression of the resistant gene in the presence of the pathogen, as has been shown for GmKR3, a TIR–NLR gene, conferring resistance to multiple viruses in soybean [27].
3.3. Quantitative Expression Analysis of the Transcript TNL Genes
We analyzed the relative normalized expression profiles of the four genes TNL1, TNL2, TNL3, and TNL6 from WFP trees infested with aphids for 24 h and from non-infested trees, using three specific primers for every gene (Table S1). The expression analysis of infected and uninfected tissues showed that TNL1 was highly overexpressed during infestation with a relative expression (RE) of 47.15 and significantly differentially expressed between infected or non-infested plants (p > 0.01) (Figure 5). TNL2 was slightly more expressed in infested plants (RE of 2.58) compared to non-infested plants (RE of 0.38), and in contrast, TNL3 and TNL6 were very weakly expressed in infected and uninfected plants with no statistical differences, with the RE of 0.02/0.17 and 0.001/0.004.
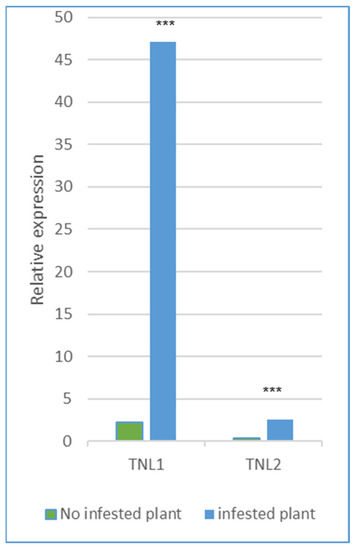
Figure 5.
Expression profiling of the two genes, TNL1 and TNL2, without aphid infestation or after green peach aphid infestation in the resistant peach Weeping Flower (WFP). Data represented the mean of three replicates. *** Significative differences between no infested and infested.
In the analysis, only the TNL1 gene expression showed a much higher value in infected plants of the resistant genotype, making the aphid resistance-associated TNL1 gene a strong candidate gene as the gene encoding the Rm1 protein. On the contrary, TNL3 and TNL6 are expressed very weakly in control and infected leaves and are therefore unlikely to be involved in aphid resistance. For the TNL2 gene, being also slightly more expressed in infected plants, we could not conclude whether it has a function in resistance or not.
3.4. Genomic Sequence of Rm2 in Rubira
The Rm2 gene of Rubira triggers the same type of reaction as the Rm1 gene of WFP and is located in the same region. One hypothesis is that Rm2 would be the same NLR gene as Rm1. To check that, we analyzed the Rm2 locus in Rubira. The NGS Rubira short reads were aligned onto the WFP Rm1bac sequence showing that the Rubira sequence was identical to the WFP Rm1bac sequence in this region, with the same four TNL genes as shown on the screenshot of the IGV viewer for TNL1 (Figure S2) with the addition of short reads of the susceptible peach Montclar. We then investigated the expression of TNL1 by RT-PCR. Transcription of the TNL1 gene of Rubira was confirmed by PCR of leaf cDNA from GPA-infected Rubira plants, using two specific primers pairs designed from the TNL1 sequence (Figure 6). The expression of TNL1 in WFP and Rubira confirmed that the TNL1 gene is a good candidate for the Rm2 gene in Rubira, as Rm1 was in WFP.
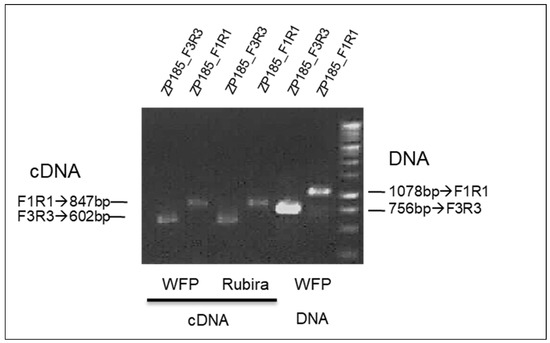
Figure 6.
Genotyping with the two primers ZP185F1R1 and ZP185F3R3 specific of the TNL1 gene, of the Rubira cDNA and Weeping flower peach (WFP) cDNA and DNA.
3.5. Marker Development for Marker Assisted Selection
Since TNL1 and, to a lesser extent, TNL2 are strong candidates for GPA-resistant genes, we decided to design dominant intragenic markers to identify other resistant peach genotypes that carry the TNL1 and TNL2 genes. We selected one size marker (ZP185-F3) for TNL1 and another (ZP186-F4) for TNL2 with an amplicon size of 756 bp and 669 bp for genomic DNA and 602 pb and 515 bp for cDNA, respectively. The presence or the absence of the amplicon by PCR reveals whether the gene is present or not. The gel with the resistant genotypes (WFP, Rubira) and three susceptible genotypes (Pamir, Montclar, and Nemared) is represented in Figure 7. These markers were validated on the progeny of the double cross (Pamir × Rubira) × (Montclar × Nemared) (PRMN), including the resistant parent, Rubira. In the 108 hybrids of the PRMN population, 63 were genotyped and phenotyped as resistant, while 45 hybrids were genotyped and phenotyped as susceptible.
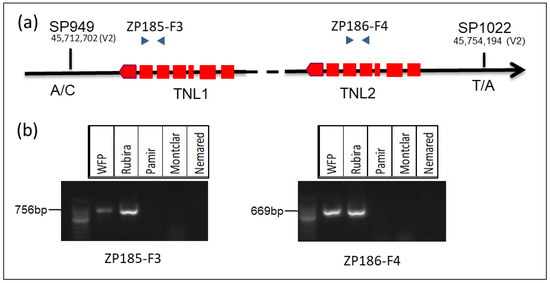
Figure 7.
Genotyping for the TNL1 and TNL2 genes of resistant and susceptible peach parents. (a) Position of the SNP and specific size markers (b) genotyping with the size markers ZP185-F3 and ZP186-F4; the presence of amplicon for resistant and absence of amplicon for susceptible genotypes.
We also developed Kasp markers with SNPs flanking the two genes TNL1 and TNL2. The two Kasp markers that were the most efficient and replicable were SP949, located 18 kb before TNL1, and SP1022, located 20 kb after TNL2 (Figure 7a). The Kasp marker genotyping of the 108 PRMN hybrids was perfectly consistent with the phenotyping. Likewise, the 30 susceptible French peach accessions of the INRAE collection were genotyped as susceptible with the two Kasp markers. These two types of markers are good markers which can be useful in peach breeding programs to integrate the two genes TNL1 and TNL2.
4. Discussion
Genetic resistance to aphids in peach varieties is a preferable control method to chemical control with insecticides. For this reason, Massonié et al. [8] introduced a wide variety of peaches from different origins and developed inoculation tests to find resistant varieties. After identifying the Weeping Peach Varieties S2678 (WFP) and the rootstock Rubira, several mapping studies were conducted by INRAE teams to localize the region of the Rm1 and Rm2 genes. In another study [14], a third similar gene, Rm3, was localized in the same region in a narrower interval. In this work, we built a BAC library of the resistant variety WFP to obtain the genomic sequence of this new targeted interval and found that additional TNL genes typical of dominant resistances are present in this genotype and are different from those present in the susceptible peach reference Lovell. Long-read RNAseq analysis showed that four TNL genes (TNL1, TNL2, TNL3, and TNL6) were expressed, and another expression analysis by qPCR showed that the TNL1 gene had a very high expression level in aphid-infected plants compared to the non-infested plants; TNL2 was slightly more expressed and the other two genes were not overexpressed. In addition, with the NGS resequencing of the resistant Rubira, we could see that WFP and Rubira have the same TNL genes in this region with an identical genomic sequence, and we could confirm that TNL1 and TNL2 were well expressed in Rubira as in WFP.
We hypothesized that the three genes Rm1, Rm2, and Rm3, being localized in the same region, were closely related. This was confirmed by the latest work on Rm3 by Pan et al. [15], who found a candidate gene for the aphid-resistant gene Rm3 in peach “Fen Shouxing” that provides the same necrotic reaction as Rm1 and Rm2 when infected with M. persicae. They used the same methodology as ours to identify the Rm3 gene using a BAC library and PacBio sequencing. They also found two new NLR genes, NLR1 and NLR2, and identified NRL1 as a strong candidate gene for Rm3. The comparison of the genomic sequence and transcripts of TNL1 and NRL1 shows that they are identical. They did not publish the sequence of their second gene, NLR2, but TNL2 and NRL2 could be different because NRL2 is oriented in the positive strand while our TNL2 gene is oriented in the negative strand; NLR2 was not overexpressed in the infested plants, while TNL2 was slightly overexpressed. Our results and those obtained on the Rm3 gene support the hypothesis that the three loci have the same origin and the TNL1 gene (=NLR1) is the same candidate gene for Rm1, Rm2, and Rm3. The role of TNL2 is less clear, and it has either no action in GPA resistance or perhaps an action of regulation.
Sauge et al. [28] found a slower effect of antixenosis in Rubira than in WFP. In their experiments, they counted the percentage of aphids remaining on plants after placement on the plant and in their 2006 experiments (10 aphids by experiment), after 96 h, all aphids escaped from WFP and 75% for Rubira. However, in the data of the same experiment in 2002 [29] (40 aphids by experiment), they observed that after 96 h, the aphids were almost all gone from Rubira plants, and they did not test WFP. Despite this slight difference in the escape dynamic of aphids, we showed that the two accessions carry the same TNL1 gene, suggesting that the observed difference may be linked to the level of expression of the gene TNL1, regulated by another gene such as TNL2, which has been shown for some resistances mediated by NLR pairs [30]. Another explanation may be the role of the genetic background [31], which could have an influence on the expression level for plant viruses, where the genetic background may impair effector-triggered dominant resistances at several stages by modifying the NLR response pathway.
Only two major aphid resistance genes have been cloned so far, the melon Vat gene, which confers resistance to the melon–cotton aphid Aphis gossypii [32] and the Mi-1 gene, which confers resistance to the potato aphid Macrosiphum euphorbiae [33]. The Vat gene also confers resistance to some viruses when they are transmitted by A. gossypii [34], and the gene Mi-1 also confers resistance to root-knot nematodes [35] and other insects, psyllids and whiteflies [36]. These two genes belong to the Coil–coil (CC)-NLR family, encoding a protein with a coiled-coil domain in the N-terminal extremity of the NBS region, while the genes TNL1 and TNL2 are resistance genes of the sub-family Toll/interleukin-1 receptor (TIR)-NLR. This family is thought to have appeared earlier than the CNL family [37], and many TNL genes control diverse plant pathogens such as viruses, bacteria [38], nematodes [3], or fungi. Rubira is also known as a peach rootstock for its tolerance against Pseudomonas syringae [39], and it would be interesting to check if there is an effect of the TNL1 gene on tolerance to the bacterial canker.
In this study, we identified the gene TNL1 as a major candidate gene for the two known aphid resistance genes, Rm1 in WFP and Rm2 in Rubira, identical to the NRL1 candidate gene Rm3. A second gene, TNL2, located at 35 kb of TNL1, was shown to also be weakly expressed in WFP, but its role in aphid resistance is still unclear. It would be necessary to create a recombinant peach hybrid bearing the TNL1 gene and not the TNL2 gene to know the role of each one. One other possibility would be to clone the TNL1 gene and transform a susceptible peach with a TNL1 construction. In vitro peach regeneration and genetic transfer with Agrobacterium is difficult, but the first transgenic peach plant carrying the cry1Ab gene for insect resistance was successfully produced recently [40].
Using the sequence of the two genes and the region of the locus, we developed specific Kasp markers and intragenic markers to integrate these major aphid resistance genes in new peach rootstocks and peach varieties, which will require several backcrosses to improve the poor fruit quality of the two peach varieties WFP and Rubira. These markers would be indispensable if we want to cumulate aphid-resistant QTLs such as those detected in the wild peach, Prunus davidiana [41], with major resistance genes.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/genes13081489/s1, Table S1: list and sequences of primers; Table S2; list of French peach accessions genotyped; Figure S1: Output of FGENESH TNL data in the FGENESH analysis of the sequence Rm1bac, Figure S2: mapping view of long-read and NGS read on IGV; Figure S3: TNL1 protein sequence with annotations. Figure S4: TNL2 protein sequence with annotations.
Author Contributions
Conceptualization, H.D.; Data curation, L.H., N.D. and C.C.; Formal analysis, J.L.; Methodology, H.D., N.D. and J.L.; Software, H.D., L.H., C.C. and J.L.; Supervision, J.L.; Writing—original draft, H.D.; Writing—review and editing, H.D., L.H. and N.D. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the INRAE Plant biology and Breeding Division.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Sequencing data for genome assemblies and contigs sequences were deposited in the NCBI database under the following numbers: Bioprojects PRJNA854381 and Biosample SAMN29438676 for Weeping Flower Peach, BioProject PRJNA854381 and Biosample SAMN29438677 for Rubira. All other relevant data are available on reasonable request to the Corresponding Author.
Acknowledgments
We thank Hugo Hussié for his help with hybrid phenotyping and the technical staff of the INRAE A2M team for the maintenance of trees in tunnels. We are grateful to the genotoul bioinformatics platform Toulouse Occitanie (Bioinfo Genotoul, https://doi.org/10.15454/1.5572369328961167E12) (accessed on 1 January 2018) for providing computing, storage resources, the MGX Platform—Montpellier GenomiX for the NGS sequencing and the INRAE Gentyane platform for the long-read RNAseq sequencing. We thank Catherine Dogimont, Jean-Luc Gallois, and Véronique Chovelon for their helpful discussion and critical reading of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aranzana, M.J.; Decroocq, V.; Dirlewanger, E.; Eduardo, I.; Gao, Z.S.; Gasic, K.; Iezzoni, A.; Jung, S.; Peace, C.; Prieto, H.; et al. Prunus genetics and applications after de novo genome sequencing: Achievements and prospects. Hortic. Res. 2019, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Gradziel, T.M. Exotic genes for solving emerging peach production challenges. Sci. Hortic. 2022, 295, 110801. [Google Scholar] [CrossRef]
- Claverie, M.; Dirlewanger, E.; Bosselut, N.; Van Ghelder, C.; Voisin, R.; Kleinhentz, M.; Lafargue, B.; Abad, P.; Rosso, M.-N.; Chalhoub, B.; et al. The Ma Gene for Complete-Spectrum Resistance to Meloidogyne Species in Prunus Is a TNL with a Huge Repeated C-Terminal Post-LRR Region. Plant Physiol. 2011, 156, 779–792. [Google Scholar] [CrossRef] [PubMed]
- Van Ghelder, C.; Esmenjaud, D.; Callot, C.; Dubois, E.; Mazier, M.; Duval, H. Ma Orthologous Genes in Prunus spp. Shed Light on a Noteworthy NBS-LRR Cluster Conferring Differential Resistance to Root-Knot Nematodes. Front. Plant Sci. 2018, 9, 1269. [Google Scholar] [CrossRef]
- Marimon, N.; Luque, J.; Arús, P.; Eduardo, I. Fine mapping and identification of candidate genes for the peach powdery mildew resistance gene Vr3. Hortic. Res. 2020, 7, 175. [Google Scholar] [CrossRef]
- Verde, I.; Abbott, A.; Scalabrin, S.; Jung, S.; Shu, S.; Marroni, F.; Zhebentyayeva, T.; Dettori, M.; Grimwood, J.; Cattonaro, F.; et al. The high-quality draft genome of peach (Prunus persica) identifies unique patterns of genetic diversity, domestication and genome evolution. Nat. Genet. 2013, 45, 487–494. [Google Scholar] [CrossRef]
- Verde, I.; Jenkins, J.; Dondini, L.; Micali, S.; Pagliarani, G.; Vendramin, E.; Paris, R.; Aramini, V.; Gazza, L.; Rossini, L.; et al. The Peach v2.0 release: High-resolution linkage mapping and deep resequencing improve chromosome-scale assembly and contiguity. BMC Genom. 2017, 18, 225. [Google Scholar] [CrossRef]
- Massonié, G.; Maison, P.; Monet, R.; Grasselly, C. Resistance to green peach aphid Myzus-Persicae Sulzer in Prunus persica (L) Batsch and others Prunus. Agronomie 1982, 2, 63–70. [Google Scholar] [CrossRef]
- Massonie, G.; Maison, P. Preliminary-Results on the Study of the Resistance of Prunus-Persica L (batsch) Varieties to Myzus-Persicae Sulz and Myzus-Varians Davids. Ann. Zool. Ecol. Anim. 1979, 11, 479–485. [Google Scholar]
- Monet, R.; Massonie, G. Genetic Determinism of Resistance to the Green Aphid (myzus-Persicae) in the Peach—Further Results. Agronomie 1994, 14, 177–182. [Google Scholar] [CrossRef][Green Version]
- Pascal, T.; Aberlenc, R.; Confolent, C.; Hoerter, M.; Lecerf, E.; Tuero, C.; Lambert, P. Mapping of new resistance (Vr2, Rm1) and ornamental (Di2, pl) Mendelian trait loci in peach. Euphytica 2017, 213, 132. [Google Scholar] [CrossRef]
- Lambert, P.; Pascal, T. Mapping Rm2 gene conferring resistance to the green peach aphid (Myzus persicae Sulzer) in the peach cultivar “Rubira (R)”. Tree Genet. Genomes 2011, 7, 1057–1068. [Google Scholar] [CrossRef]
- Sauge, M.H.; Kervella, J.; Pascal, T. Settling behaviour and reproductive potential of the green peach aphid Myzus persicae on peach varieties and a related wild Prunus. Entomol. Exp. Appl. 1998, 89, 233–242. [Google Scholar] [CrossRef]
- Niu, L.; Lu, Z.; Zeng, W.; Cui, G.; Pan, L.; Xu, Q.; Li, G.; Wang, Z. Inheritance analysis of resistance to green peach aphids(Myzus persicae Sulzer)for peach cultivar‘Fen Shouxing’(Prunus persica var. densa). J. Fruit Sci. 2016, 33, 578–584. [Google Scholar]
- Pan, L.; Lu, Z.; Yan, L.; Zeng, W.; Shen, Z.; Yu, M.; Bu, L.; Cui, G.; Niu, L.; Wang, Z. NLR1 is a strong candidate for the Rm3 dominant green peach aphid (Myzus persicae) resistance trait in peach. J. Exp. Bot. 2022, 73, 1357–1369. [Google Scholar] [CrossRef]
- Lambert, P.; Campoy, J.A.; Pacheco, I.; Mauroux, J.-B.; Linge, C.D.S.; Micheletti, D.; Bassi, D.; Rossini, L.; Dirlewanger, E.; Pascal, T.; et al. Identifying SNP markers tightly associated with six major genes in peach [Prunus persica (L.) Batsch] using a high-density SNP array with an objective of marker-assisted selection (MAS). Tree Genet. Genomes 2016, 12, 121. [Google Scholar] [CrossRef]
- Zhang, N.; Lu, Z.; Cui, G.; Pan, L.; Zeng, W.; Niu, L.; Wang, Z. Gene mapping of aphid-resistant for peach using SNP markers. Sci. Agric. Sin. 2017, 50, 4613–4621. [Google Scholar]
- Duval, H.; Hoerter, M.; Polidori, J.; Confolent, C.; Masse, M.; Moretti, A.; Van Ghelder, C.; Esmenjaud, D. High-resolution mapping of the RMia gene for resistance to root-knot nematodes in peach. Tree Genet. Genomes 2014, 10, 297–306. [Google Scholar] [CrossRef]
- Peterson, D.G.; Tomkins, J.P.; Frisch, D.A.; Paterson, A.H. Construction of plant bacterial artificial chromosome (bac) libraries: An illustrated guide. J. Agric. Genom. 2000, 5, 1–100. [Google Scholar]
- Gonthier, L.; Bellec, A.; Blassiau, C.; Prat, E.; Helmstetter, N.; Rambaud, C.; Huss, B.; Hendriks, T.; Berges, H.; Quillet, M.-C. Construction and characterization of two BAC libraries representing a deep-coverage of the genome of chicory (Cichorium intybus L., Asteraceae). BMC Res. Notes 2010, 3, 225. [Google Scholar] [CrossRef]
- Chalhoub, B.; Belcram, H.; Caboche, M. Efficient cloning of plant genomes into bacterial artificial chromosome (BAC) libraries with larger and more uniform insert size. Plant Biotechnol. J. 2004, 2, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.; Alexander, D.; Marks, P.; Klammer, A.; Drake, J.; Heiner, C.; Clum, A.; Copeland, A.; Huddleston, J.; Eichler, E.; et al. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat. Methods 2013, 10, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative Genomics Viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [PubMed]
- Solovyev, V.; Kosarev, P.; Seledsov, I.; Vorobyev, D. Automatic annotation of eukaryotic genes, pseudogenes and promoters. Genome Biol. 2006, 7, S10. [Google Scholar] [CrossRef]
- Van Ghelder, C.; Esmenjaud, D. TNL genes in peach: Insights into the post-LRR domain. BMC Genom. 2016, 17, 317. [Google Scholar] [CrossRef]
- Meyers, B.; Kozik, A.; Griego, A.; Kuang, H.; Michelmore, R. Genome-wide analysis of NBS-LRR-encoding genes in Arabidopsis. Plant Cell 2003, 15, 809–834. [Google Scholar] [CrossRef]
- Xun, H.; Yang, X.; He, H.; Wang, M.; Guo, P.; Wang, Y.; Pang, J.; Dong, Y.; Feng, X.; Wang, S.; et al. Over-expression of GmKR3, a TIR–NBS–LRR type R gene, confers resistance to multiple viruses in soybean. Plant Mol. Biol 2019, 99, 95–111. [Google Scholar] [CrossRef]
- Sauge, M.; Mus, F.; Lacroze, J.; Pascal, T.; Kervella, J.; Poessel, J. Genotypic variation in induced resistance and induced susceptibility in the peach—Myzus persicae aphid system. Oikos 2006, 113, 305–313. [Google Scholar] [CrossRef]
- Sauge, M.H.; Lacroze, J.P.; Poessel, J.L.; Pascal, T.; Kervella, J. Induced resistance by Myzus persicae in the peach cultivar “Rubira. ” Entomol. Exp. Appl. 2002, 102, 29–37. [Google Scholar] [CrossRef]
- Eitas, T.K.; Dangl, J.L. NB-LRR proteins: Pairs, pieces, perception, partners, and pathways. Curr. Opin. Plant Biol. 2010, 13, 472–477. [Google Scholar] [CrossRef]
- Gallois, J.; Moury, B.; German-Retana, S. Role of the Genetic Background in Resistance to Plant Viruses. Int. J. Mol. Sci. 2018, 19, 2856. [Google Scholar] [CrossRef] [PubMed]
- Dogimont, C.; Chovelon, V.; Pauquet, J.; Boualem, A.; Bendahmane, A. The Vat locus encodes for a CC-NBS-LRR protein that confers resistance to Aphis gossypii infestation and A. gossypii-mediated virus resistance. Plant J. 2014, 80, 993–1004. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Goggin, F.; Milligan, S.; Kaloshian, I.; Ullman, D.; Williamson, V. The nematode resistance gene Mi of tomato confers resistance against the potato aphid. Proc. Natl. Acad. Sci. USA 1998, 95, 9750–9754. [Google Scholar] [CrossRef] [PubMed]
- Boualem, A.; Dogimont, C.; Bendahmane, A. The battle for survival between viruses and their host plants. Curr. Opin. Virol. 2016, 17, 32–38. [Google Scholar] [CrossRef]
- Milligan, S.; Bodeau, J.; Yaghoobi, J.; Kaloshian, I.; Zabel, P.; Williamson, V. The root knot nematode resistance gene Mi from tomato is a member of the leucine zipper, nucleotide binding, leucine-rich repeat family of plant genes. Plant Cell 1998, 10, 1307–1319. [Google Scholar] [CrossRef] [PubMed]
- Nombela, G.; Williamson, V.; Muniz, M. The root-knot nematode resistance gene Mi-1.2 of tomato is responsible for resistance against the whitefly Bemisia tabaci. Mol. Plant-Microbe Interact. 2003, 16, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.; Meyers, B.; Chen, J.; Tian, D.; Yang, S. Tracing the origin and evolutionary history of plant nucleotide-binding site-leucine-rich repeat (NBS-LRR) genes. New Phytol. 2012, 193, 1049–1063. [Google Scholar] [CrossRef]
- Gassmann, W.; Hinsch, M.; Staskawicz, B. The Arabidopsis RPS4 bacterial-resistance gene is a member of the TIR-NBS-LRR family of disease-resistance genes. Plant J. 1999, 20, 265–277. [Google Scholar] [CrossRef]
- Reighard, G.L.; Newall, W.C.; Beckman, T.G.; Okie, W.R.; Zehr, E.I.; Nyczepir, A.P. Field performance of Prunus rootstock cultivars and selections on replant soils in South Carolina. In Proceedings of the International Symposium on Integrating Canopy, Rootstocks and Environmental Physiology in Orchard Systems, Proceedings—Vols 1 and 2, Stellenbosch, South Africa, 3–6 December 2012; Barritt, B.H., Kappel, F., Eds.; International Society Horticultural Science: Leuven, Belgium, 1997; pp. 243–249. [Google Scholar]
- Kadasa, N.M.; Metwali, E.M.R.; Soliman, H.I.A.; Alshehri, W.A. Creation of borer pests resistance genetically engineering peach (Prunus persica L.) plants by constitutively overexpressing the cry1Ab gene. Plant Cell Tiss Organ Cult. 2022, 148, 465–477. [Google Scholar] [CrossRef]
- Sauge, M.-H.; Lambert, P.; Pascal, T. Co-localisation of host plant resistance QTLs affecting the performance and feeding behaviour of the aphid Myzus persicae in the peach tree. Heredity 2012, 108, 292–301. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).