Single Nucleotide Polymorphisms of ALDH18A1 and MAT2A Genes and Their Genetic Associations with Milk Production Traits of Chinese Holstein Cows



Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Selection, Pedigree, and Phenotypic Data Collation
2.2. Genomic DNA Extraction from Frozen Semen and Blood Samples
2.3. Polymorphism Detection of Candidate Genes
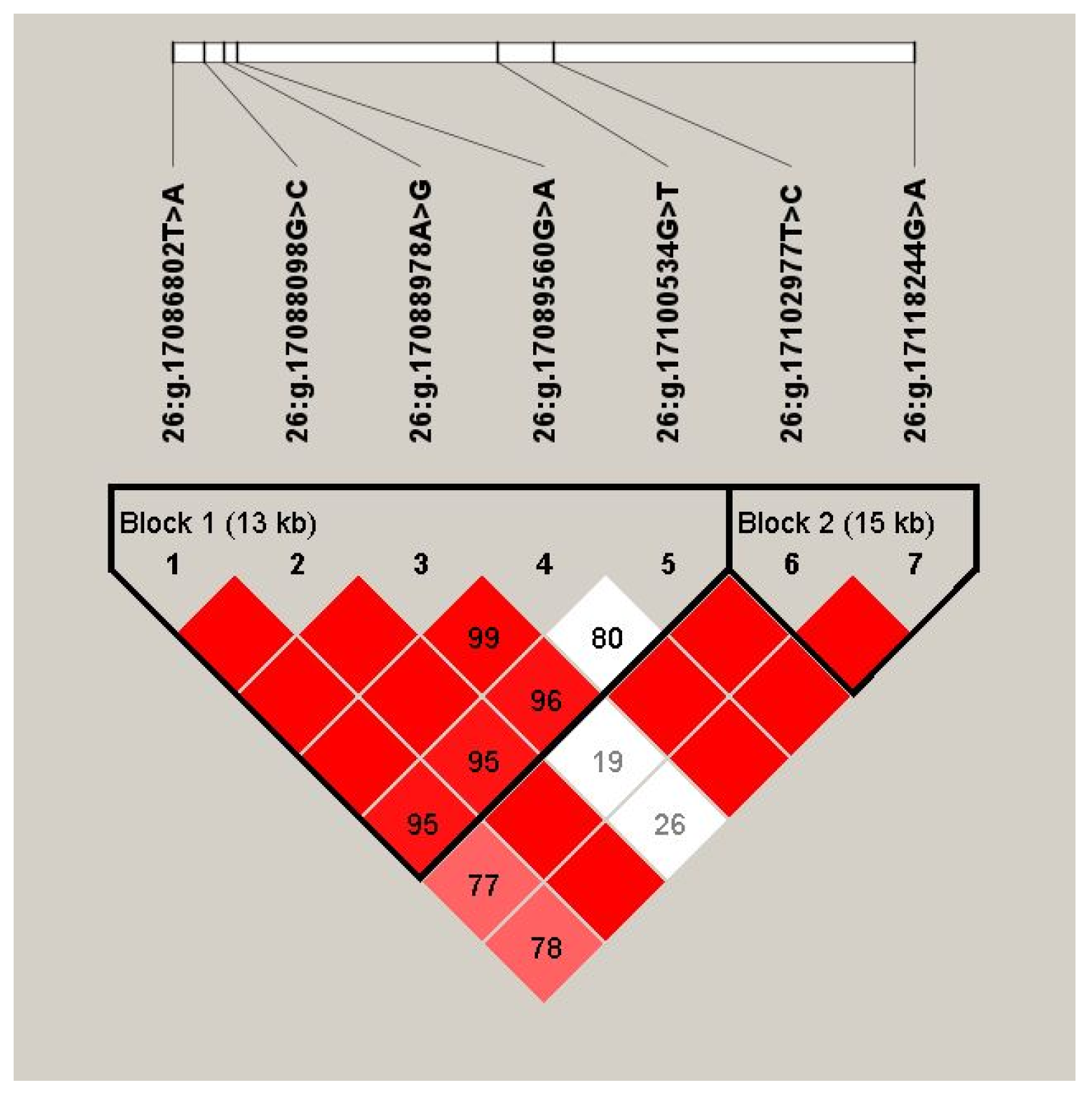
2.4. Linkage Disequilibrium (LD) Estimation
2.5. Association Analysis of Single Marker/Haplotype and Milk Production Traits
2.6. Biological Function Prediction
3. Results
3.1. SNPs Identification
3.2. Single Marker Association Analysis
3.3. Haplotype Association Analysis
3.4. Functional Variation Prediction Caused by SNPs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tunick, M.H.; Van Hekken, D.L. Dairy Products and Health: Recent Insights. J. Agric. Food Chem. 2015, 63, 9381–9388. [Google Scholar] [CrossRef] [PubMed]
- Nieman, K.M.; Anderson, B.D.; Cifelli, C.J. The Effects of Dairy Product and Dairy Protein Intake on Inflammation: A Systematic Review of the Literature. J. Am. Coll. Nutr. 2020, 40, 571–582. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yang, X.; Xia, J.; Zhao, L.; Yang, Y. Consumption of meat and dairy products in China: A review. Proc. Nutr. Soc. 2016, 75, 385–391. [Google Scholar] [CrossRef]
- Wang, Y.; Li, S. Worldwide trends in dairy production and consumption and calcium intake: Is promoting consumption of dairy products a sustainable solution for inadequate calcium intake? Food Nutr. Bull. 2008, 29, 172–185. [Google Scholar] [CrossRef]
- Hu, B. Development of China’s Dairy Industry in 2018 and Its Prospect for 2019; Agricultural Outlook: Beijing, China, 2019. [Google Scholar]
- Jin, S.; Yuan, R.; Zhang, Y.; Jin, X. Chinese Consumers’ Preferences for Attributes of Fresh Milk: A Best-Worst Approach. Int. J. Environ. Res. Public Health 2019, 16, 4286. [Google Scholar] [CrossRef]
- Andersson, L.; Georges, M. Domestic-animal genomics: Deciphering the genetics of complex traits. Nat. Rev. Genet. 2004, 5, 202–212. [Google Scholar] [CrossRef]
- Chen, Y.J.; Zhang, Z.Q.; Wang, M.W.; Qiu, Y.S.; Yuan, R.Y.; Dong, E.L.; Zhao, Z.; Zhou, H.T.; Wang, N.; Chen, W.J.; et al. Novel Compound Missense and Intronic Splicing Mutation in ALDH18A1 Causes Autosomal Recessive Spastic Paraplegia. Front. Neurol. 2021, 12, 627531. [Google Scholar] [CrossRef]
- Fu, Y.; Jia, R.; Xu, L.; Su, D.; Li, Y.; Liu, L.; Ma, Z.; Sun, D.; Han, B. Fatty acid desaturase 2 affects the milk-production traits in Chinese Holsteins. Anim. Genet. 2022, 53, 422–426. [Google Scholar] [CrossRef]
- Jia, R.; Fu, Y.; Xu, L.; Li, H.; Li, Y.; Liu, L.; Ma, Z.; Sun, D.; Han, B. Associations between polymorphisms of SLC22A7, NGFR, ARNTL and PPP2R2B genes and Milk production traits in Chinese Holstein. BMC Genom. Data 2021, 22, 47. [Google Scholar] [CrossRef]
- Napolitano, F.; Grandoni, F.; De Matteis, G.; Degano, L.; Vicario, D.; Buttazzoni, L. Novel SNPs and haplotypes identified in the CD4 gene and their influence on deregressed MACE EBV indexes of milk-related traits in Simmental breed. J. Dairy Res. 2021, 88, 368–373. [Google Scholar] [CrossRef]
- Potu, H.; FA, L.; K, A.; MT, D.; TV, A. High resolution melt curve analysis identifies a novel SNP (G21A) in butyrophilin gene having significant association with milk production traits in Holstein Friesian crossbreds of Kerala. Anim. Biotechnol. 2022, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Liang, Y.; Gao, Q.; Guo, J.; Tang, C.; Shi, K.; Yang, Z.; Mao, Y. AGPAT3 Gene polymorphisms are associated with milk production traits in Chinese Holstein cows. J Dairy Res. 2021, 88, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Ober, U.; Erbe, M.; Zhang, H.; Gao, N.; He, J.; Li, J.; Simianer, H. Improving the accuracy of whole genome prediction for complex traits using the results of genome wide association studies. PLoS ONE 2014, 9, e93017. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Erbe, M.; He, J.; Ober, U.; Gao, N.; Zhang, H.; Simianer, H.; Li, J. Accuracy of whole-genome prediction using a genetic architecture-enhanced variance-covariance matrix. G3 (Bethesda) 2015, 5, 615–627. [Google Scholar] [CrossRef]
- de Las Heras-Saldana, S.; Lopez, B.I.; Moghaddar, N.; Park, W.; Park, J.E.; Chung, K.Y.; Lim, D.; Lee, S.H.; Shin, D.; van der Werf, J.H.J. Use of gene expression and whole-genome sequence information to improve the accuracy of genomic prediction for carcass traits in Hanwoo cattle. Genet Sel. Evol. 2020, 52, 54. [Google Scholar] [CrossRef]
- Ibeagha-Awemu, E.M.; Kgwatalala, P.; Xin, Z. A critical analysis of production-associated DNA polymorphisms in the genes of cattle, goat, sheep, and pig. Mamm. Genome 2008, 19, 591–617. [Google Scholar] [CrossRef]
- Li, F.; Cai, C.; Qu, K.; Liu, J.; Lei, C. DGAT1 K232A polymorphism is associated with milk production traits in Chinese cattle. Anim. Biotechnol. 2021, 32, 427–431. [Google Scholar] [CrossRef]
- Cruz, V.A.R.; Oliveira, H.R.; Brito, L.F.; Fleming, A.; Larmer, S.; Miglior, F.; Schenkel, F.S. Genome-Wide Association Study for Milk Fatty Acids in Holstein Cattle Accounting for the DGAT1 Gene Effect. Animals 2019, 9, 997. [Google Scholar] [CrossRef]
- Bobbo, T.; Tiezzi, F.; Penasa, M.; Marchi, M.D.; Cassandro, M. Short communication: Association analysis of diacylglycerol acyltransferase (DGAT1) mutation on chromosome 14 for milk yield and composition traits, somatic cell score, and coagulation properties in Holstein bulls. J. Dairy Sci. 2018, 101, 8087–8091. [Google Scholar] [CrossRef]
- Xu, L.; Shi, L.; Liu, L.; Liang, R.; Li, Q.; Li, J.; Han, B.; Sun, D. Analysis of Liver Proteome and Identification of Critical Proteins Affecting Milk Fat, Protein, and Lactose Metabolism in Dariy Cattle with iTRAQ. Proteomics 2019, 19, e1800387. [Google Scholar] [CrossRef]
- Hu, C.A.A.; Lin, W.W.; Obie, C.; Valle, D. Molecular Enzymology of Mammalian Δ1-Pyrroline-5-carboxylate Synthase. J. Biol. Chem. 1999, 274, 6754–6762. [Google Scholar] [CrossRef] [PubMed]
- Tong, B.C.; Barbul, A. Cellular and physiological effects of arginine. Mini. Rev. Med. Chem. 2004, 4, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Ramani, K.; Yang, H.; Kuhlenkamp, J.; Tomasi, L.; Tsukamoto, H.; Mato, J.M.; Lu, S.C. Changes in the expression of methionine adenosyltransferase genes and S-adenosylmethionine homeostasis during hepatic stellate cell activation. Hepatology 2010, 51, 986–995. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Chen, X.; Wu, W.; Wang, W.; Pang, W.; Yang, G. MAT2B promotes adipogenesis by modulating SAMe levels and activating AKT/ERK pathway during porcine intramuscular preadipocyte differentiation. Exp. Cell Res. 2016, 344, 11–21. [Google Scholar] [CrossRef]
- Martínez-Chantar, M.L.; Latasa, M.U.; Varela-Rey, M.; Lu, S.C.; García-Trevijano, E.R.; Mato, J.M.; Avila, M.A. L-methionine availability regulates expression of the methionine adenosyltransferase 2A gene in human hepatocarcinoma cells: Role of S-adenosylmethionine. J. Biol. Chem. 2003, 278, 19885–19890. [Google Scholar] [CrossRef]
- Zhao, C.; Wu, H.; Qimuge, N.; Pang, W.; Li, X.; Chu, G.; Yang, G. MAT2A promotes porcine adipogenesis by mediating H3K27me3 at Wnt10b locus and repressing Wnt/beta-catenin signaling. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 132–142. [Google Scholar] [CrossRef]
- Stram, D.O. Multi-SNP Haplotype Analysis Methods for Association Analysis. Methods Mol. Biol. 2017, 1666, 485–504. [Google Scholar]
- Vadva, Z.; Larsen, C.E.; Propp, B.E.; Trautwein, M.R.; Alford, D.R.; Alper, C.A. A New Pedigree-Based SNP Haplotype Method for Genomic Polymorphism and Genetic Studies. Cells 2019, 8, 835. [Google Scholar] [CrossRef]
- Pérez-Enciso, M.; Rincón, J.C.; Legarra, A. Sequence- vs. chip-assisted genomic selection: Accurate biological information is advised. Genet. Sel. Evol. 2015, 47, 43. [Google Scholar] [CrossRef]
- Habier, D.; Fernando, R.L.; Dekkers, J. Genomic Selection Using Low-Density Marker Panels. Genetics 2009, 182, 343–353. [Google Scholar] [CrossRef]
- Mustafa, H. Performance of bovine high density SNPs genotyping array in indigenous Pakistani cattle breeds. Pure Appl. Biol. 2018, 7, 221–226. [Google Scholar] [CrossRef]
- Grossi, D.A.; Brito, L.F.; Jafarikia, M.; Schenkel, F.S.; Feng, Z. Genotype imputation from various low-density SNP panels and its impact on accuracy of genomic breeding values in pigs. Animal 2018, 12, 2235–2245. [Google Scholar] [CrossRef] [PubMed]
- Bolormaa, S.; Gore, K.; Van, D.; Hayes, B.J.; Daetwyler, H.D. Design of a low-density SNP chip for the main Australian sheep breeds and its effect on imputation and genomic prediction accuracy. Anim. Genet. 2015, 46, 544–556. [Google Scholar] [CrossRef] [PubMed]
- Zuk, O.; Schaffner, S.F.; Samocha, K.; Do, R.; Hechter, E.; Kathiresan, S.; Daly, M.J.; Neale, B.M.; Sunyaev, S.R.; Lander, E.S. Searching for missing heritability: Designing rare variant association studies. Proc. Natl. Acad. Sci. USA 2014, 111, E455–E464. [Google Scholar] [CrossRef]
- Calkhoven, C.F.; Ab, G. Multiple steps in the regulation of transcription-factor level and activity. Biochem. J. 1996, 317, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Kasowski, M.; Grubert, F.; Heffelfinger, C.; Hariharan, M.; Asabere, A.; Waszak, S.M.; Habegger, L.; Rozowsky, J.; Shi, M.; Urban, A.E.; et al. Variation in transcription factor binding among humans. Science 2010, 328, 232–235. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, C.; Zheng, X.; Tian, Z.; Zhang, J. HMBOX1, homeobox transcription factor, negatively regulates interferon-γ production in natural killer cells. Int. Immunopharmacol. 2011, 11, 1895–1900. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Wu, L.; Zhang, C.; Zheng, X.; Tian, Z.; Zhang, J. Recombinant expression of a novel human transcriptional repressor HMBOX1 and preparation of anti-HMBOX1 monoclonal antibody. Cell Mol. Immunol. 2009, 6, 261–268. [Google Scholar] [CrossRef][Green Version]
- Chen, S.; Saiyin, H.; Zeng, X.; Xi, J.; Liu, X.; Li, X.; Yu, L. Isolation and functional analysis of human HMBOX1, a homeobox containing protein with transcriptional repressor activity. Cytogenet. Genome Res. 2006, 114, 131–136. [Google Scholar] [CrossRef]
- Luo, W.; Zhu, X.; Liu, W.; Ren, Y.; Bei, C.; Qin, L.; Miao, X.; Tang, F.; Tang, G.; Tan, S. MYC associated zinc finger protein promotes the invasion and metastasis of hepatocellular carcinoma by inducing epithelial mesenchymal transition. Oncotarget 2016, 7, 86420–86432. [Google Scholar] [CrossRef]
- Su, J.M.; Lai, X.M.; Lan, K.H.; Li, C.P.; Chao, Y.; Yen, S.H.; Chang, F.Y.; Lee, S.D.; Lee, W.P. X protein of hepatitis B virus functions as a transcriptional corepressor on the human telomerase promoter. Hepatology 2007, 46, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.X.; Zhao, W.P.; Kishi, H.; Dokan, J.; Jin, Z.X.; Wei, X.C.; Yokoyama, K.K.; Muraguchi, A. Activation of mouse RAG-2 promoter by Myc-associated zinc finger protein. Biochem. Biophys. Res. Commun. 2004, 317, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.D.; Li, Y.J. Expression of telomerase, p53 and CK19 and their significance in hepatocellular carcinoma. Chin. J. Clin. Exp. Pathol. 2012, 28, 862–866. [Google Scholar]


{kind=link}
| Gene | SNP Name | RS ID | Position (UMD3.1) | Gene Region | Genotype | Genotypic Frequency | Allele | Allelic Frequency | Hardy–Weinberg Equilibrium |
|---|---|---|---|---|---|---|---|---|---|
| ALDH18A1 | 26:g.17130318C>A | rs109124430 | Chr26: | 5′ flanking region | AA | 0.6840 | A | 0.8274 | T |
| 17130318 | AC | 0.2868 | C | 0.1726 | |||||
| CC | 0.0292 | ||||||||
| 26:g.17118244G>A | rs133295794 | Chr26: | Exon (synonymous) | AA | 0.0195 | A | 0.1239 | T | |
| 17118244 | AG | 0.2089 | G | 0.8761 | |||||
| GG | 0.7716 | ||||||||
| 26:g.17102977T>C | rs136218403 | Chr26: | Intron | CC | 0.0216 | C | 0.1261 | T | |
| 17102977 | CT | 0.2089 | T | 0.8739 | |||||
| TT | 0.7695 | ||||||||
| 26:g.17100534G>T | rs110706194 | Chr26: | Exon (synonymous) | GG | 0.8680 | G | 0.9324 | T | |
| 17100534 | GT | 0.1288 | T | 0.0676 | |||||
| TT | 0.0032 | ||||||||
| 26:g.17089560G>A | rs458180458 | Chr26: | Exon (synonymous) | AA | 0.0119 | A | 0.0947 | T | |
| 17089560 | AG | 0.1656 | G | 0.9053 | |||||
| GG | 0.8225 | ||||||||
| 26:g.17088978A>G | rs41255559 | Chr26: | 3′UTR (untranslated region) | AA | 0.0606 | A | 0.2262 | T | |
| 17088978 | AG | 0.3312 | G | 0.7738 | |||||
| GG | 0.6082 | ||||||||
| 26:g.17088098G>C | rs208352883 | Chr26: | 3′ flanking region | CC | 0.3506 | C | 0.5812 | T | |
| 17088098 | CG | 0.4610 | G | 0.4188 | |||||
| GG | 0.1883 | ||||||||
| 26:g.17086802T>A | rs109201383 | Chr26: | 3′ flanking region | AA | 0.3712 | A | 0.6012 | T | |
| 17086802 | AT | 0.4600 | T | 0.3988 | |||||
| TT | 0.1688 | ||||||||
| MAT2A | 11:g.49472723G>C | rs109079969 | Chr26: | 5′ flanking region | CC | 0.6396 | C | 0.8003 | T |
| 49472723 | CG | 0.3214 | G | 0.1997 | |||||
| GG | 0.0390 | ||||||||
| 11:g.49465032C>T | rs110124316 | Chr26: | 3′ flanking region | CC | 0.4913 | C | 0.6813 | F | |
| 49465032 | CT | 0.3798 | T | 0.3187 | |||||
| TT | 0.1288 |
| Genes | SNPs | Lactation | Genotype (No.) | Milk Yield (kg) | Fat Yield (kg) | Fat Percentage (%) | Protein Yield (kg) | Protein Percentage (%) |
|---|---|---|---|---|---|---|---|---|
| ALDH18A1 | 26:g.17130318C>A | 1 | AA (632) | 10,312 ± 59.78 | 339.4 ± 2.66 | 3.31 ± 0.02 | 305.9 ± 1.94 | 2.98 ± 0.02 |
| AC (265) | 10,145 ± 69.33 | 338.84 ± 3.01 | 3.37 ± 0.03 | 302.59 ± 2.19 | 3.00 ± 0.02 | |||
| CC (27) | 10,211 ± 149.06 a | 339.89 ± 6.11 | 3.36 ± 0.06 | 305.07 ± 4.45 | 3.00 ± 0.04 | |||
| 8.76 | 0.07 | 6.86 | 3.92 | 2.15 | ||||
| p value | 0.0128 | 0.8714 | 0.0329 | 0.1405 | 0.3414 | |||
| Corrected p | 0.1024 | 6.9712 | 0.2632 | 1.124 | 2.7312 | |||
| 2 | AA (437) | 10,850 ± 61.29 | 388.69 ± 2.73 Aa | 3.59 ± 0.03 | 322 ± 1.99 Aa | 2.97 ± 0.02 | ||
| AC (181) | 10,694 ± 76.68 | 383.71 ± 3.31 Aa | 3.59 ± 0.03 | 315.51 ± 2.41 ABb | 2.95 ± 0.02 | |||
| CC (17) | 10,331 ± 194.55 | 349.48 ± 7.96 Bb | 3.40 ± 0.08 | 299.99 ± 5.80 Bb | 2.91 ± 0.05 | |||
| 10.81 | 26.7 | 6.09 | 22.03 | 3.14 | ||||
| p value | 0.0047 | <0.0001 | 0.0485 | <0.0001 | 0.2088 | |||
| Corrected p | 0.0376 | <0.0001 | 0.388 | <0.0001 | 1.6704 | |||
| 26:g.17118244G>A | 1 | AA (18) | 10,232 ± 179.35 | 344.82 ± 7.31 | 3.39 ± 0.07 | 304.58 ± 5.33 | 2.99 ± 0.04 | |
| AG (193) | 10,292 ± 75.23 | 342.37 ± 3.24 | 3.36 ± 0.03 | 306.49 ± 2.36 | 3.00 ± 0.02 | |||
| GG (713) | 10,251 ± 58.52 | 338.26 ± 2.62 | 3.32 ± 0.02 | 304.47 ± 1.90 | 2.98 ± 0.02 | |||
| 0.46 | 3.36 | 2.64 | 1.24 | 0.76 | ||||
| p value | 0.796 | 0.187 | 0.2675 | 0.5381 | 0.6851 | |||
| Corrected p | 6.368 | 1.496 | 2.14 | 4.3048 | 5.4808 | |||
| 2 | AA (12) | 10,527 ± 227.41 ab | 362.45 ± 9.26 a | 3.46 ± 0.09 | 307.38 ± 6.75 | 2.93 ± 0.05 | ||
| AG (133) | 10,997 ± 84.80 a | 391.85 ± 3.63 b | 3.57 ± 0.03 | 325.40 ± 2.64 | 2.96 ± 0.02 | |||
| GG (490) | 10,751 ± 60.15 b | 385.76 ± 2.69 ab | 3.59 ± 0.02 | 318.67 ± 1.96 | 2.97 ± 0.02 | |||
| 11.76 | 11.5 | 2.25 | 12.73 | 0.71 | ||||
| p value | 0.003 | 0.0034 | 0.3248 | 0.0019 | 0.7024 | |||
| Corrected p | 0.024 | 0.0272 | 2.5984 | 0.0152 | 5.6192 | |||
| 26:g.17102977T>C | 1 | CC (20) | 10,240 ± 171.14 | 343.5 ± 6.98 | 3.37 ± 0.07 | 305.74 ± 5.09 | 3.00 ± 0.04 | |
| CT (193) | 10,278 ± 75.15 | 342.12 ± 3.23 | 3.36 ± 0.03 | 306 ± 2.36 | 3.00 ± 0.02 | |||
| TT (711) | 10,255 ± 58.58 | 338.33 ± 2.62 | 3.32 ± 0.02 | 304.55 ± 1.91 | 2.98 ± 0.02 | |||
| 0.16 | 2.73 | 2.72 | 0.67 | 0.8 | ||||
| p value | 0.9253 | 0.2564 | 0.2577 | 0.7137 | 0.6691 | |||
| Corrected p | 7.4024 | 2.0512 | 2.0616 | 5.7096 | 5.3528 | |||
| 2 | CC (12) | 10,524 ± 227.4 | 362.32 ± 9.26 | 3.46 ± 0.09 | 307.29 ± 6.75 | 2.93 ± 0.05 | ||
| CT (134) | 10,978 ± 84.37 | 390.97 ± 3.61 | 3.57 ± 0.03 | 324.86 ± 2.63 | 2.96 ± 0.02 | |||
| TT (489) | 10,755 ± 60.20 | 385.96 ± 2.69 B | 3.59 ± 0.02 | 318.78 ± 1.96 | 2.97 ± 0.02 | |||
| 10.11 | 10.31 | 2.45 | 11.22 | 0.71 | ||||
| p value | 0.0067 | 0.0061 | 0.2946 | 0.0039 | 0.7021 | |||
| Corrected p | 0.0536 | 0.0488 | 2.3568 | 0.0312 | 5.6168 | |||
| 26:g.17100534G>T | 1 | GG (802) | 10,306 ± 58.10 Aa | 340.22 ± 2.60 | 3.33 ± 0.02 | 305.91 ± 1.89 | 2.98 ± 0.02 | |
| GT (119) | 9977.38 ± 87.89 Bb | 333.57 ± 3.72 | 3.38 ± 0.04 | 298.81 ± 2.71 | 3.01 ± 0.02 | |||
| TT (3) | 10,230 ± 421.67 ab | 337.7 ± 17.04 | 3.33 ± 0.17 | 305.44 ± 12.43 | 3.00 ± 0.10 | |||
| 17.74 | 4.41 | 2.48 | 9.48 | 2.19 | ||||
| p value | 0.0002 | 0.1109 | 0.2903 | 0.009 | 0.3343 | |||
| Corrected p | 0.0016 | 0.8872 | 2.3224 | 0.072 | 2.6744 | |||
| 2 | GG (550) | 10,878 ± 59.18 Aa | 389.39 ± 2.66 Aa | 3.58 ± 0.02 | 322.33 ± 1.93 Aa | 2.97 ± 0.02 | ||
| GT (82) | 10,222 ± 104.36 Bb | 366.03 ± 4.39 Bb | 3.58 ± 0.04 | 301.48 ± 3.20 Bb | 2.95 ± 0.03 | |||
| TT (3) | 10,129 ± 433.12 ab | 361.25 ± 17.55 ab | 3.59 ± 0.17 | 302.88 ± 12.802 ab | 2.97 ± 0.10 | |||
| 44.84 | 34.64 | 0 | 51.09 | 0.58 | ||||
| p value | <0.0001 | <0.0001 | 0.9985 | <0.0001 | 0.7473 | |||
| Corrected p | <0.0001 | <0.0001 | 7.988 | <0.0001 | 5.9784 | |||
| 26:g.17089560G>A | 1 | AA (11) | 10,748 ± 231.33 | 354.55 ± 9.42 | 3.30 ± 0.09 | 321.51 ± 6.87 | 2.98 ± 0.06 | |
| AG (153) | 10,285 ± 84.85 | 339.32 ± 3.63 | 3.33 ± 0.03 | 304.86 ± 2.65 | 2.98 ± 0.02 | |||
| GG (760) | 10,253 ± 57.75 | 339.19 ± 2.58 | 3.33 ± 0.02 | 304.83 ± 1.88 | 2.99 ± 0.02 | |||
| 4.77 | 2.9 | 0.15 | 6.47 | 0.22 | ||||
| p value | 0.0928 | 0.2354 | 0.9278 | 0.0398 | 0.8947 | |||
| Corrected p | 0.7424 | 1.8832 | 7.4224 | 0.3184 | 7.1576 | |||
| 2 | AA (8) | 10,490 ± 295.13 | 378.43 ± 12.00 | 3.61 ± 0.12 | 319.19 ± 8.7487 | 3.03 ± 0.07 | ||
| AG (103) | 10,640 ± 95.67 | 378.74 ± 4.06 | 3.57 ± 0.04 | 315.39 ± 2.9595 | 2.97 ± 0.02 | |||
| GG (524) | 10,827 ± 59.57 | 387.96 ± 2.67 | 3.59 ± 0.02 | 320.55 ± 1.944 | 2.96 ± 0.02 | |||
| 5.16 | 6.55 | 0.28 | 3.73 | 0.97 | ||||
| p value | 0.0766 | 0.0387 | 0.8696 | 0.1559 | 0.6172 | |||
| Corrected p | 0.6128 | 0.3096 | 6.9568 | 1.2472 | 4.9376 | |||
| 26:g.17088978A>G | 1 | AA (56) | 10,161 ± 117.76 | 341.96 ± 4.89 | 3.39 ± 0.05 | 305.17 ± 3.57 | 3.01 ± 0.03 | |
| AG (306) | 10,250 ± 67.89 | 338.46 ± 2.96 | 3.33 ± 0.03 | 305.17 ± 2.16 | 2.99 ± 0.02 | |||
| GG (562) | 10,273 ± 60.16 | 339.35 ± 2.67 | 3.33 ± 0.02 | 304.71 ± 1.95 | 2.98 ± 0.02 | |||
| 1.02 | 0.65 | 1.71 | 0.09 | 1.44 | ||||
| p value | 0.6 | 0.7229 | 0.4256 | 0.9564 | 0.487 | |||
| Corrected p | 4.8 | 5.7832 | 3.4048 | 7.6512 | 3.896 | |||
| 2 | AA (42) | 10,538 ± 139.32 AB | 362.98 ± 5.77 Aa | 3.42 ± 0.06 Aa | 309.2 ± 4.21 Aa | 2.93 ± 0.03 | ||
| AG (206) | 10,581 ± 74.38 A | 383.18 ± 3.22 Bb | 3.63 ± 0.03 Bb | 314.45 ± 2.35 Aa | 2.97 ± 0.02 | |||
| GG (387) | 10,927 ± 63.06 B | 390.75 ± 2.80 Bb | 3.58 ± 0.03 ab | 323.48 ± 2.04 Bb | 2.97 ± 0.02 | |||
| 27.58 | 25.8 | 14.13 | 25.1 | 1.74 | ||||
| p value | <0.0001 | <0.0001 | 0.0009 | <0.0001 | 0.4186 | |||
| Corrected p | <0.0001 | <0.0001 | 0.0072 | <0.0001 | 3.3488 | |||
| 26:g.17088098G>C | 1 | CC (324) | 10,333 ± 66.66 | 341.06 ± 2.93 | 3.32 ± 0.03 | 305.57 ± 2.13 | 2.97 ± 0.02 | |
| CG (426) | 10,215 ± 62.67 | 337.21 ± 2.76 | 3.33 ± 0.03 | 304.24 ± 2.01 | 2.99 ± 0.02 | |||
| GG (174) | 10,225 ± 78.95 | 341.27 ± 3.38 | 3.37 ± 0.03 | 305.25 ± 2.46 | 3.00 ± 0.02 | |||
| 4.57 | 3.89 | 2.16 | 0.73 | 3.49 | ||||
| p value | 0.1025 | 0.1436 | 0.3401 | 0.696 | 0.175 | |||
| Corrected p | 0.82 | 1.1488 | 2.7208 | 5.568 | 1.4 | |||
| 2 | CC (233) | 10,970 ± 71.62 Aa | 396.06 ± 3.13 Aa | 3.61 ± 0.03 | 324.66 ± 2.28 Aa | 2.97 ± 0.02 | ||
| CG (277) | 10,708 ± 68.62 ABb | 383.33 ± 3.01 Bb | 3.59 ± 0.03 | 317.99 ± 2.19 ABb | 2.97 ± 0.02 | |||
| GG (125) | 10,641 ± 87.94 Bb | 374.74 ± 3.74 Bb | 3.52 ± 0.04 | 314.05 ± 2.73 Bb | 2.95 ± 0.02 | |||
| 17.34 | 34.84 | 6.37 | 16.71 | 1.12 | ||||
| p value | 0.0002 | <0.0001 | 0.0423 | 0.0003 | 0.5723 | |||
| Corrected p | 0.0016 | <0.0001 | 0.3384 | 0.0024 | 4.5784 | |||
| 26:g.17086802T>A | 1 | AA (343) | 10,321 ± 65.81 | 341.52 ± 2.89 | 3.33 ± 0.03 | 305.33 ± 2.11 | 2.97 ± 0.02 | |
| AT (425) | 10,217 ± 62.63 | 336.33 ± 2.76 | 3.32 ± 0.03 | 304.26 ± 2.01 | 2.99 ± 0.02 | |||
| TT (156) | 10,234 ± 81.80 | 343.07 ± 3.49 | 3.38 ± 0.03 | 305.75 ± 2.54 | 3.00 ± 0.02 | |||
| 3.65 | 8.43 | 4.55 | 0.73 | 3.27 | ||||
| p value | 0.1615 | 0.0151 | 0.1034 | 0.6944 | 0.1954 | |||
| Corrected p | 1.292 | 0.1208 | 0.8272 | 5.5552 | 1.5632 | |||
| 2 | AA (247) | 11,016 ± 70.14 Aa | 396.72 ± 3.07 Aa | 3.60 ± 0.03 | 325.69 ± 2.23 Aa | 2.96 ± 0.02 | ||
| AT (272) | 10,666 ± 69.12 Bb | 382.36 ± 3.03 Bb | 3.59 ± 0.03 | 316.75 ± 2.20 Bb | 2.97 ± 0.02 | |||
| TT (116) | 10,580 ± 90.67 Bb | 372.28 ± 3.85 Bb | 3.52 ± 0.04 | 312.89 ± 2.80 Bb | 2.96 ± 0.02 | |||
| 31.11 | 45.41 | 5.8 | 26.55 | 0.65 | ||||
| p value | <0.0001 | <0.0001 | 0.0559 | <0.0001 | 0.7242 | |||
| Corrected p | <0.0001 | <0.0001 | 0.4472 | <0.0001 | 5.7936 | |||
| MAT2A | 11:g.49472723G>C | 1 | CC (591) | 10,307 ± 60.23 a | 339.22 ± 2.68 | 3.31 ± 0.02 | 304.92 ± 1.95 | 2.97 ± 0.02 a |
| CG (297) | 10,150 ± 67.98 b | 338.26 ± 2.96 | 3.36 ± 0.03 | 304.07 ± 2.15 | 3.01 ± 0.02 b | |||
| GG (36) | 10,376 ± 137.72 ab | 348.4 ± 5.68 | 3.39 ± 0.06 | 311.23 ± 4.14 | 3.01 ± 0.03 ab | |||
| 9.05 | 3.54 | 6.34 | 3.33 | 10.16 | ||||
| p value | 0.0111 | 0.1707 | 0.0426 | 0.1899 | 0.0064 | |||
| Corrected p | 0.0222 | 0.3414 | 0.0852 | 0.3798 | 0.0128 | |||
| 2 | CC (410) | 10,829 ± 62.7056 | 387.57 ± 2.79 | 3.58 ± 0.03 | 320.29 ± 2.03 | 2.96 ± 0.02 | ||
| CG (201) | 10,707 ± 74.7301 | 384.71 ± 3.24 | 3.59 ± 0.03 | 318.45 ± 2.36 | 2.97 ± 0.02 | |||
| GG (24) | 11,006 ± 168.94 | 383.24 ± 6.93 | 3.48 ± 0.07 | 322.02 ± 5.05 | 2.92 ± 0.04 | |||
| 4.96 | 1.22 | 2.7 | 1.05 | 2.13 | ||||
| p value | 0.085 | 0.5426 | 0.2596 | 0.5909 | 0.3455 | |||
| Corrected p | 0.17 | 1.0852 | 0.5192 | 1.1818 | 0.691 | |||
| 11:g.49465032C>T | 1 | CC (454) | 10,313 ± 62.47 a | 340.38 ± 2.76 | 3.33 ± 0.03 | 306.92 ± 2.01 a | 2.99 ± 0.02 | |
| CT (351) | 10,167 ± 64.95 b | 336.71 ± 2.85 | 3.34 ± 0.03 | 302.05 ± 2.07 b | 2.98 ± 0.02 | |||
| TT (119) | 10,327 ± 86.35 ab | 342.48 ± 3.66 | 3.33 ± 0.04 | 305.58 ± 2.67 ab | 2.97 ± 0.02 | |||
| 8.8 | 4.57 | 0.14 | 9.83 | 1.07 | ||||
| p value | 0.0126 | 0.1022 | 0.9317 | 0.0075 | 0.5869 | |||
| Corrected p | 0.0252 | 0.2044 | 1.8634 | 0.015 | 1.1738 | |||
| 2 | CC (316) | 10,914 ± 66.32 A | 387.23 ± 2.92 ab | 3.55 ± 0.03 | 323.86 ± 2.13 Aa | 2.97 ± 0.02 | ||
| CT (235) | 10,763 ± 71.46 AB | 389.58 ± 3.11 a | 3.62 ± 0.03 | 316.58 ± 2.27 Bb | 2.95 ± 0.02 | |||
| TT (84) | 10,496 ± 99.15 B | 376.99 ± 4.17 b | 3.60 ± 0.04 | 314.32 ± 3.04 Bb | 3.00 ± 0.03 | |||
| 18.81 | 9.26 | 6.05 | 18.36 | 5.1 | ||||
| p value | <0.0001 | 0.0102 | 0.0495 | 0.0001 | 0.0791 | |||
| Corrected p | <0.0001 | 0.0204 | 0.099 | 0.0002 | 0.1582 |
| Block | Lactation | Haplotype Combination | Milk Yield (kg) | Fat Yield (kg) | Fat Percentage (%) | Protein Yield (kg) | Protein Percentage (%) |
|---|---|---|---|---|---|---|---|
| Block1 | 1 | H1H1 (322) | 10,425 ± 67.38 a | 344.75 ± 2.98 | 3.32 ± 0.03 | 308.87 ± 2.17 ab | 2.97 ± 0.02 |
| H1H2 (174) | 10,260 ± 77.58 ab | 336.27 ± 3.36 | 3.30 ± 0.032 | 305.59 ± 2.45 ab | 2.99 ± 0.02 | ||
| H1H3 (92) | 10,466 ± 96.58 a | 343.07 ± 4.10 | 3.29 ± 0.04 | 311.62 ± 2.99 ab | 2.99 ± 0.03 | ||
| H1H4 (68) | 10,422 ± 105.67 ab | 347.38 ± 4.43 | 3.34 ± 0.04 | 313.52 ± 3.23 a | 3.02 ± 0.03 | ||
| H1H5 (71) | 10,097 ± 105.9 b | 334.61 ± 4.46 | 3.34 ± 0.04 | 301.61 ± 3.25 b | 3.00 ± 0.03 | ||
| 15.59 | 14.34 | 2.01 | 14.25 | 4.59 | |||
| p value | 0.0039 | 0.0067 | 0.7333 | 0.007 | 0.3327 | ||
| Corrected p | 0.0078 | 0.0134 | 1.4666 | 0.014 | 0.6654 | ||
| 2 | H1H1 (231) | 11,060 ± 71.76 | 398.4 ± 3.15 | 3.60 ± 0.03 | 328.22 ± 2.29 | 2.97 ± 0.02 | |
| H1H2 (108) | 10,964 ± 90.72 | 388.92 ± 3.86 | 3.55 ± 0.04 | 327.25 ± 2.81 | 2.98 ± 0.02 | ||
| H1H3 (60) | 10,704 ± 115.33 | 381.95 ± 4.83 | 3.59 ± 0.05 | 319.31 ± 3.52 | 2.99 ± 0.03 | ||
| H1H4 (44) | 10,447 ± 132.22 | 379.36 ± 5.51 | 3.62 ± 0.05 | 310.99 ± 4.02 | 2.98 ± 0.03 | ||
| H1H5 (50) | 10,320 ± 128.01 | 373.52 ± 5.33 | 3.62 ± 0.05 | 307.29 ± 3.89 | 2.98 ± 0.03 | ||
| 50.87 | 35.23 | 2.6 | 47.84 | 0.53 | |||
| p value | <0.0001 | <0.0001 | 0.769 | <0.0001 | 0.9707 | ||
| Corrected p | <0.0001 | <0.0001 | 1.538 | <0.0001 | 1.9414 | ||
| Block2 | 1 | H1H1 (711) | 10,351 ± 58.33 | 343.72 ± 2.61 | 3.34 ± 0.02 | 307.27 ± 1.90 | 2.97 ± 0.02 |
| H1H2 (191) | 10,342 ± 75.13 | 343.81 ± 3.24 | 3.35 ± 0.03 | 306.99 ± 2.36 | 2.98 ± 0.02 | ||
| 0.03 | 0 | 0.39 | 0.02 | 0.08 | |||
| p value | 0.8736 | 0.9716 | 0.5314 | 0.879 | 0.7734 | ||
| Corrected p | 1.7472 | 1.9432 | 1.0628 | 1.758 | 1.5468 | ||
| 2 | H1H1 (489) | 10,804 ± 61.05 Aa | 386.12 ± 2.73 a | 3.5771 ± 0.03 | 321.12 ± 1.99 Aa | 2.97 ± 0.02 | |
| H1H2 (133) | 11,083 ± 83.35 Bb | 393.98 ± 3.55 b | 3.5669 ± 0.03388 | 328.79 ± 2.59 Bb | 2.96 ± 0.02 | ||
| 12.98 | 6.29 | 0.11 | 11.25 | 0.11 | |||
| p value | 0.0003 | 0.0125 | 0.7416 | 0.0009 | 0.7456 | ||
| Corrected p | 0.0006 | 0.025 | 1.4832 | 0.0018 | 1.4912 |
| Gene | SNPs | Allele | Transcription Factor | Relative Score (≥0.80; Jasper) | Predicted Binding Site Sequence (Genomatix) |
|---|---|---|---|---|---|
| ALDH18A1 | 26:g.17130318C>A | C | HMBOX1 | 0.92 | ACTAGTTTAG |
| A | |||||
| MAT2A | 11:g.49472723G>C | G | |||
| C | MAZ | 0.84 | CGCGGCTCCCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, W.; Xu, L.; Li, Y.; Liu, L.; Ma, Z.; Sun, D.; Han, B. Single Nucleotide Polymorphisms of ALDH18A1 and MAT2A Genes and Their Genetic Associations with Milk Production Traits of Chinese Holstein Cows. Genes 2022, 13, 1437. https://doi.org/10.3390/genes13081437
Ye W, Xu L, Li Y, Liu L, Ma Z, Sun D, Han B. Single Nucleotide Polymorphisms of ALDH18A1 and MAT2A Genes and Their Genetic Associations with Milk Production Traits of Chinese Holstein Cows. Genes. 2022; 13(8):1437. https://doi.org/10.3390/genes13081437
Chicago/Turabian StyleYe, Wen, Lingna Xu, Yanhua Li, Lin Liu, Zhu Ma, Dongxiao Sun, and Bo Han. 2022. "Single Nucleotide Polymorphisms of ALDH18A1 and MAT2A Genes and Their Genetic Associations with Milk Production Traits of Chinese Holstein Cows" Genes 13, no. 8: 1437. https://doi.org/10.3390/genes13081437
APA StyleYe, W., Xu, L., Li, Y., Liu, L., Ma, Z., Sun, D., & Han, B. (2022). Single Nucleotide Polymorphisms of ALDH18A1 and MAT2A Genes and Their Genetic Associations with Milk Production Traits of Chinese Holstein Cows. Genes, 13(8), 1437. https://doi.org/10.3390/genes13081437