
Laboratory-Controlled Experiments Reveal Microbial Community Shifts during Sediment Resuspension Events



Abstract
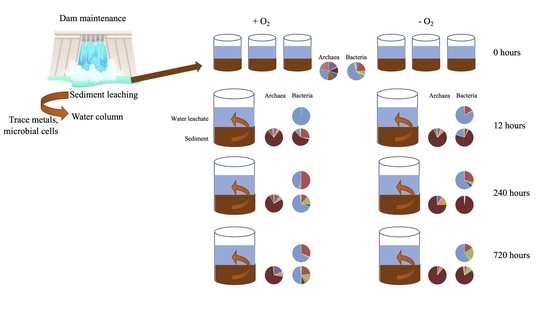
:
1. Introduction
2. Materials and Methods
2.1. Study Site, Sampling, and Incubation Setup
2.2. Chemical Analysis of Water Leachate
2.3. DNA Extraction, Sequencing and Sequence Analysis
2.4. Statistical Analyses
3. Results
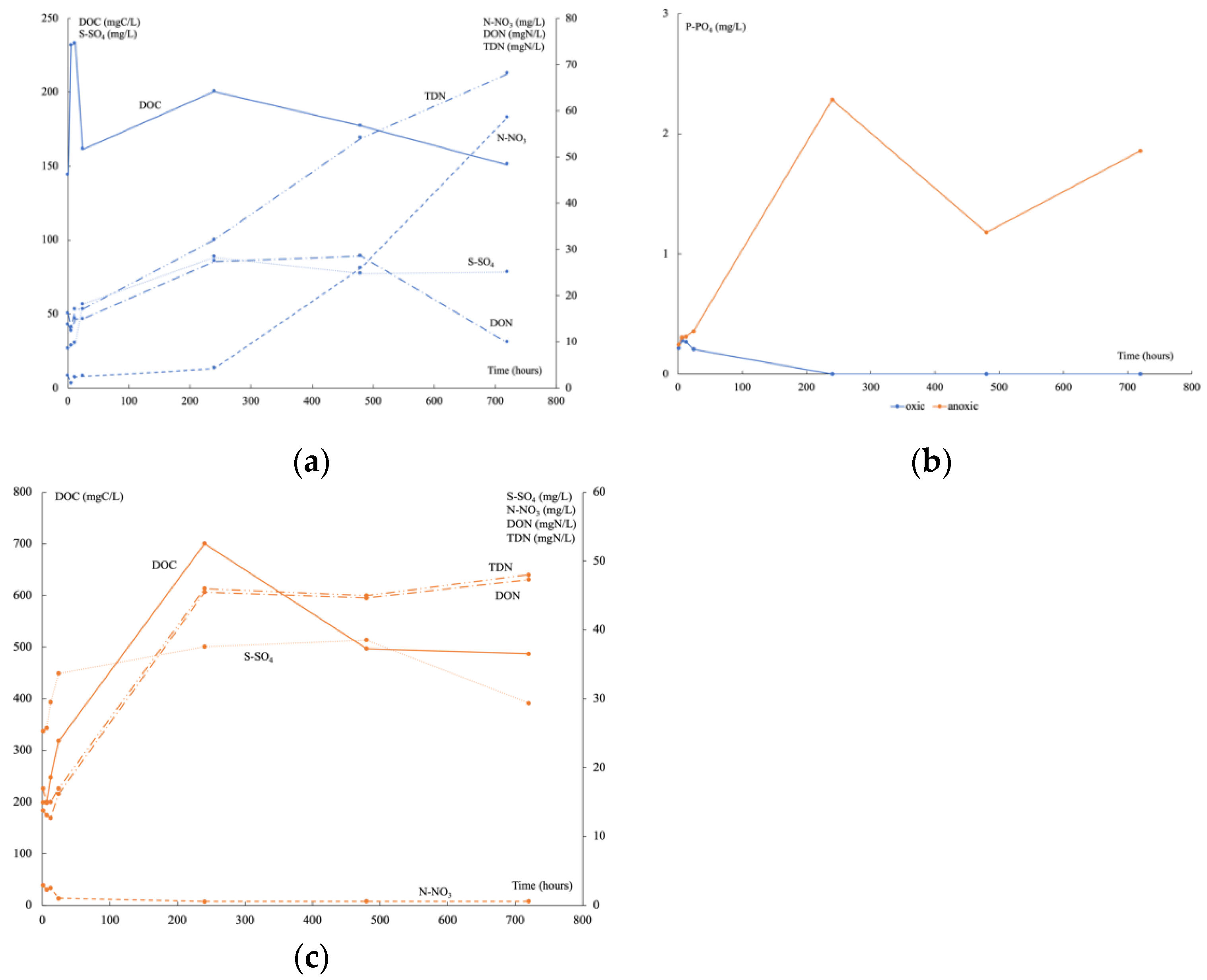
3.1. Geochemical Analyses in the Water Leachate over Time
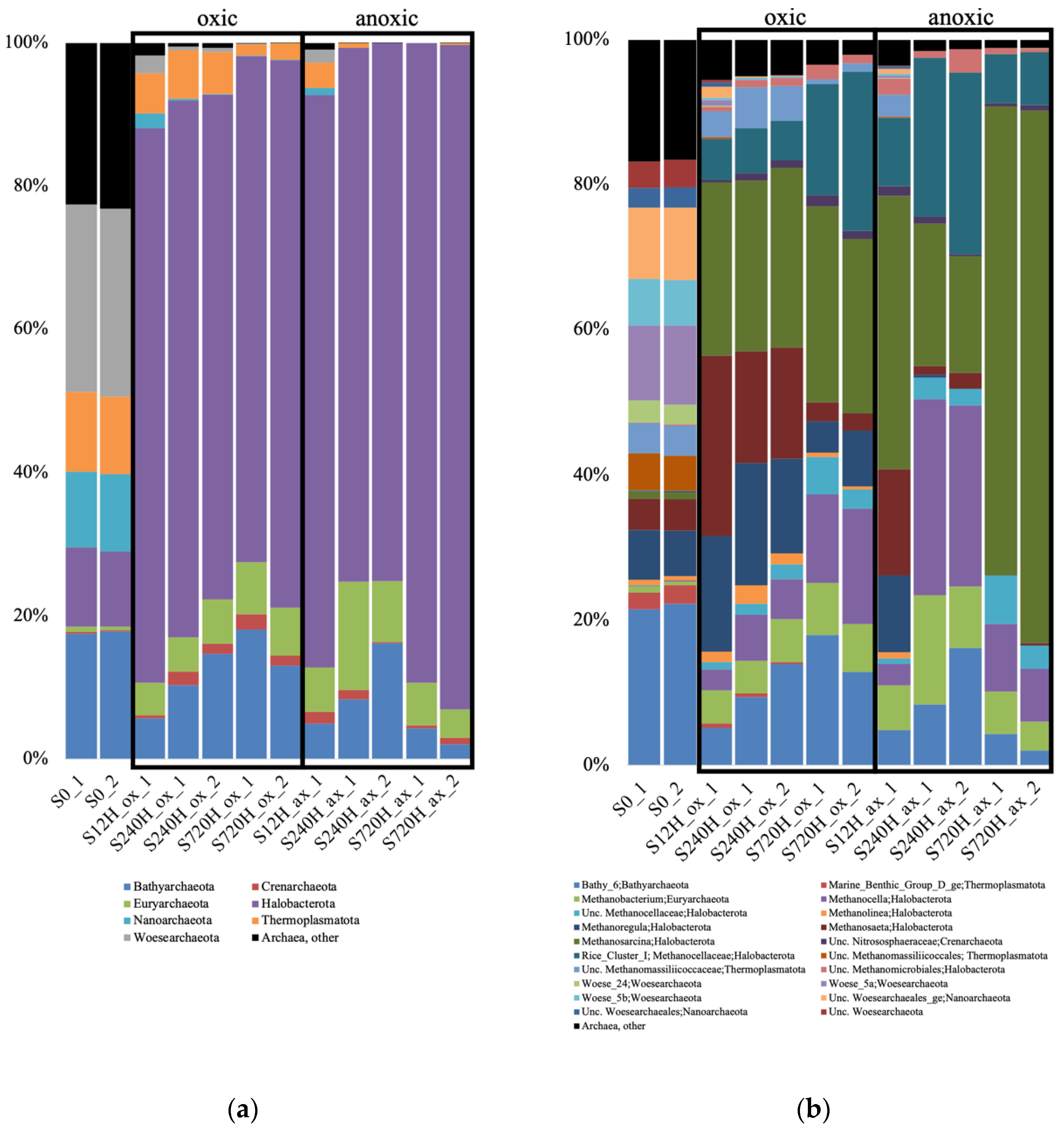
3.2. Archaeal 16S rRNA Gene Diversity in the Sediments and Leachate
3.3. Bacterial 16S rRNA Gene Diversity in the Sediments and Leachate
3.4. Archaeal and Bacterial α-Diversity Indices Comparison
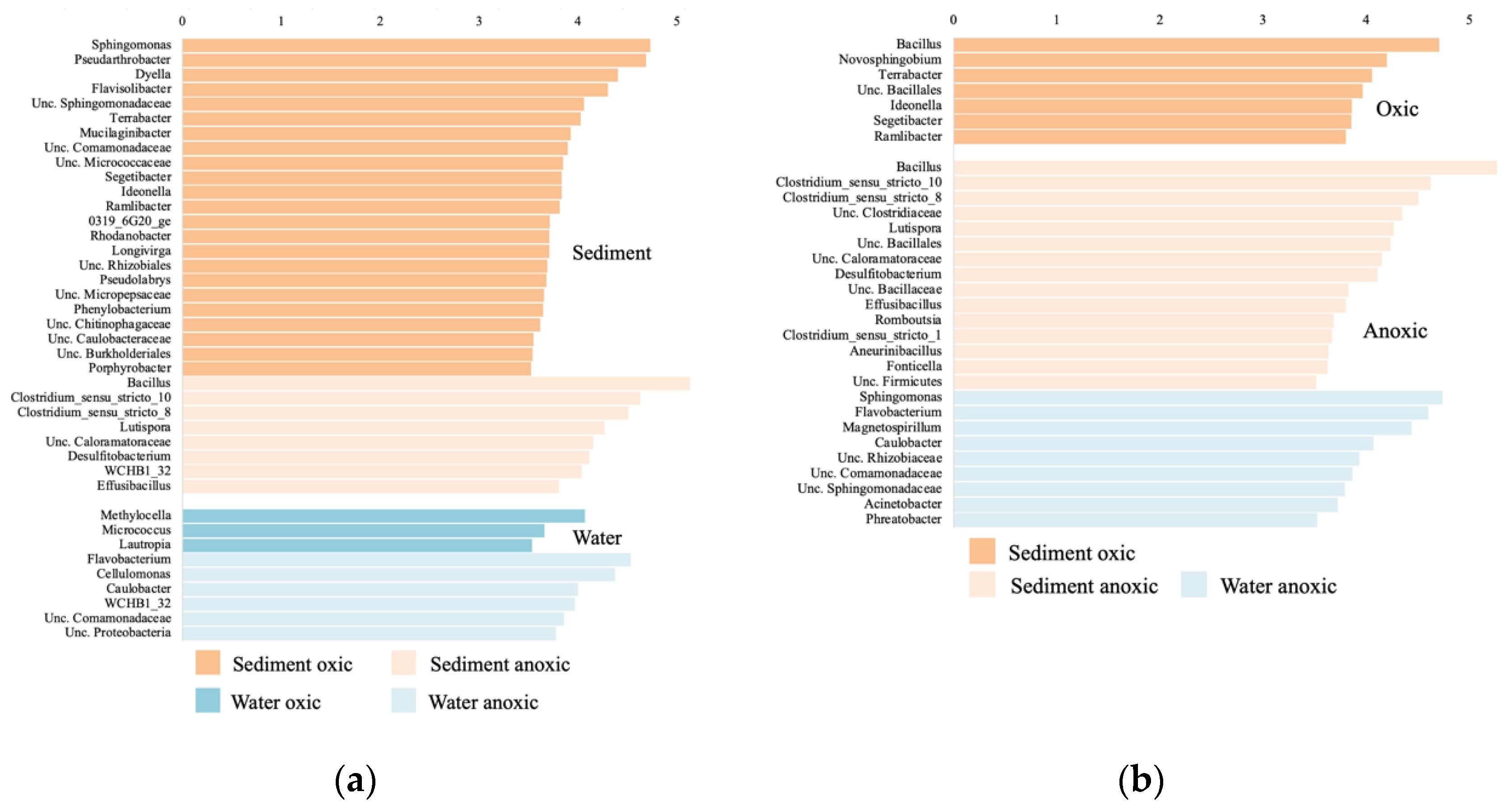
3.5. Bacterial Community Composition
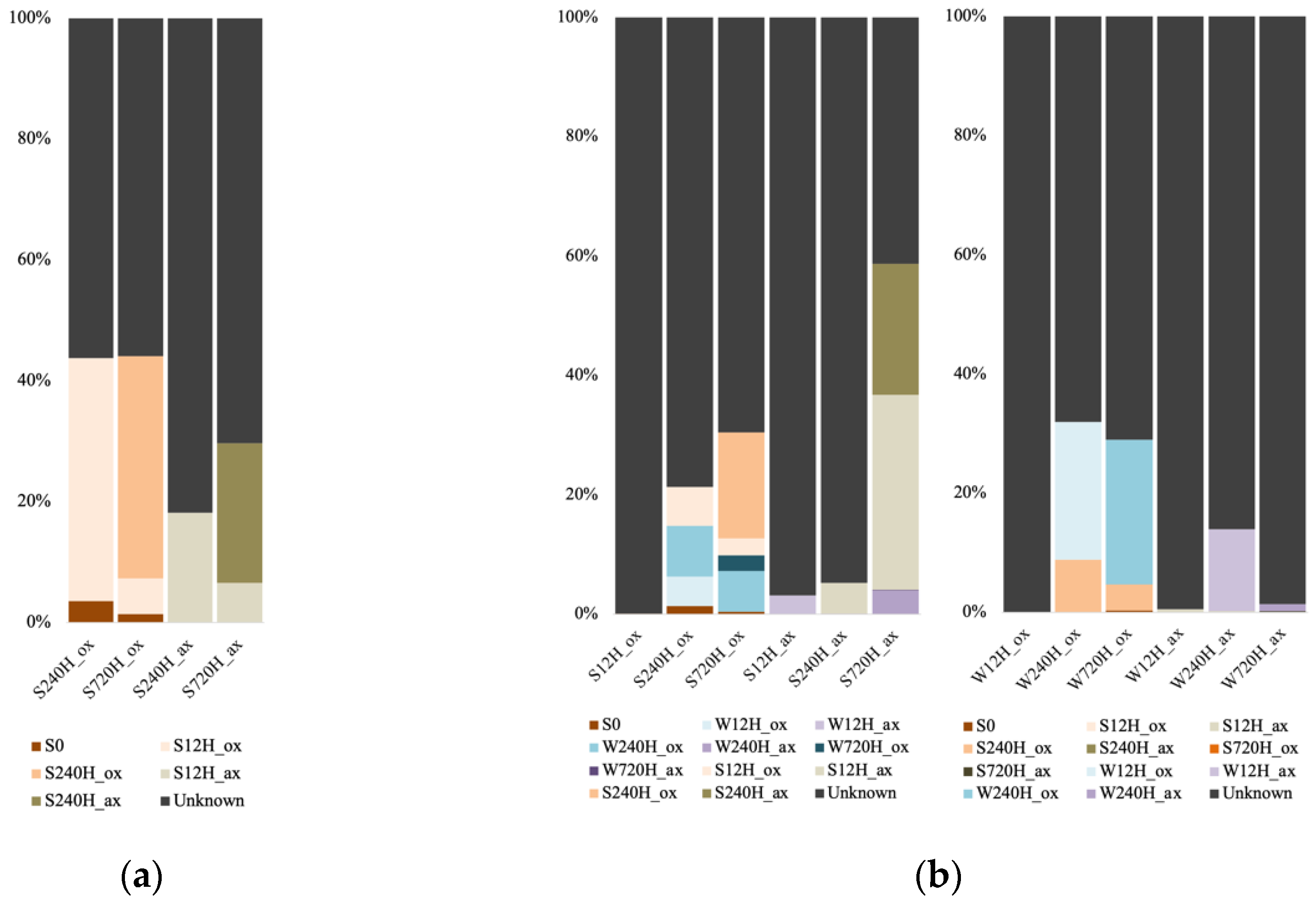
3.6. Source of Leachate and Sediment Archaeal and Bacterial Communities
4. Discussion
4.1. Organic Matter Degradation and Mineral Alteration
4.2. Microbial Community Changes in the Sediment
4.3. Potential Microbial Metabolisms in the Sediment
4.4. Leaching of Microbial Communities in the Water
4.5. Potential Microbial Metabolisms in the Water
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uroz, S.; Calvaruso, C.; Turpault, M.P.; Frey-Klett, P. Mineral weathering by bacteria: Ecology, actors and mechanisms. Trends Microbiol. 2009, 17, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Wild, B.; Daval, D.; Beaulieu, E.; Pierret, M.C.; Viville, D.; Imfelda, G. In-situ dissolution rates of silicate minerals and associated bacterial communities in the critical zone (Strengbach catchment, France). Geochim. Cosmochim. Acta 2019, 249, 95–120. [Google Scholar] [CrossRef]
- Ullman, W.J.; Kirchman, D.L.; Welch, S.A.; Vandevivere, P. Laboratory evidence for microbially mediated silicate mineral dissolution in nature. Chem. Geol. 1996, 132, 11–17. [Google Scholar] [CrossRef]
- Barker, W.W.; Welch, S.A.; Chu, S.; Banfield, J.F. Experimental observations of the effects of bacteria on aluminosilicate weathering. Am. Mineral. 1998, 83, 1551–1563. [Google Scholar] [CrossRef]
- Liermann, L.J.; Kalinowski, B.E.; Brantley, S.L.; Ferry, J.G. Role of bacterial siderophores in dissolution of hornblende. Geochim. Cosmochim. 2000, 64, 587–602. [Google Scholar] [CrossRef]
- Norris, P.R. Acidophilic bacteria and their activity in mineral sulfide oxidation. In Microbial Mineral Recovery; Erlich, H.L., Brierler, C., Eds.; McGraw-Hill: New York, NK, USA, 1990. [Google Scholar]
- Mueller, B. Experimental interactions between clay minerals and bacteria: A review. Pedosphere 2015, 25, 799–810. [Google Scholar] [CrossRef]
- Rogers, J.R.; Bennett, P.C.; Choi, W.J. Feldspars as a source of nutrients for microorganisms. Am. Mineral. 1998, 83, 1532–1540. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial phosphorus solubilization and its potential use in sustainable agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef]
- Watteau, F.; Berthelin, J. Microbial dissolution of iron and aluminum from soil minerals: Efficiency and specificity of hydrowamate siderophores compared to aliphatic acids. Eur. J. Soil. Biol. 1994, 30, 1–9. [Google Scholar]
- Kalinowski, B.E.; Liermann, L.J.; Givens, S.; Brantley, S.L. Rates of bacteria-promoted solubilization of Fe from minerals, a review of problems and approaches. Chem. Geol. 2000, 169, 357–370. [Google Scholar] [CrossRef]
- Schippers, A.; Sand, W. Bacterial leaching of metal sulfides proceeds by two indirect mechanisms via thiosulfate or via polysulfides and sulfur. Appl. Environ. Microbiol. 1999, 65, 319–321. [Google Scholar] [CrossRef] [PubMed]
- Bosecker, K. Bioleaching, metal solubilization by microorganisms. FEMS Microbiol. Rev. 1997, 20, 591–604. [Google Scholar] [CrossRef]
- Benzerara, K.; Menguy, N.; Guyot, F.; Chaïrat, C.; Oelkers, E.H.; McEldoney, S. Experimental colonization and weathering of orthopyroxenes by the pleomorphic bacteria Ramlibacter tatahouinensis. Geomicrobiol. J. 2004, 21, 341–349. [Google Scholar] [CrossRef]
- Welch, S.A.; Barker, W.W.; Banfield, J.F. Microbial extracellular polysaccharides and plagioclase dissolution. Geochim. Cosmochim. Acta 1999, 63, 1405–1419. [Google Scholar] [CrossRef]
- Zwirglmaier, K.; Keiz, K.; Engel, M.; Geist, J.; Geist, J. Seasonal and spatial patterns of microbial diversity along a trophic gradient in the interconnected lakes of Osterseen Lake District, Bavaria. Front. Microbiol. 2015, 6, 1168. [Google Scholar] [CrossRef] [PubMed]
- Wurzbacher, C.; Fuchs, A.; Attermeyer, K.; Frindte, K.; Grossart, H.-P.; Hupfer, M.; Casper, P.; Monaghan, M.T. Shifts among Eukaryota, Bacteria and Archaea define the vertical organization of a lake sediment. Microbiome 2017, 5, 41. [Google Scholar] [CrossRef]
- Gallon, C.; Tessier, A.; Gobeil, C.; La Torre, A.; Catalina, M. Modeling diagenesis of lead in sediments of a Canadian Shield lake. Geochim. Cosmochim. Acta 2004, 68, 3531–3545. [Google Scholar] [CrossRef]
- Chappaz, A.; Gobeil, C.; Tessier, A. Geochemical and anthropogenic enrichments of Mo in sediments from perennially oxic and seasonally anoxic lakes in Eastern Canada. Geochim. Cosmochim. Acta 2008, 72, 170–184. [Google Scholar] [CrossRef]
- Couture, R.M.; Gobeil, C.; Tessier, A. Arsenic, iron and sulfur co-diagenesis in lake sediments. Geochim. Cosmochim. Acta 2010, 74, 1238–1255. [Google Scholar] [CrossRef]
- Cooke, G.D.; Welch, E.B.; Peterson, S.; Nichols, S.A. Restoration and Management of Lakes and Reservoirs, 2nd ed.; Lewis Publishers: Boca Raton, FL, USA, 1993. [Google Scholar]
- Carr, M.K.; Sadeghian, A.; Lindenschmidt, K.A.; Rinke, K.; Morales-Marin, L. Impacts of varying dam outflow elevations on water temperature, dissolved oxygen, and nutrient distribution in a large prairie reservoir. Environ. Eng. Sci. 2020, 37, 78–97. [Google Scholar] [CrossRef]
- Gassama, N.; Cocirta, C.; Kasper, H.U. Use of major and selected trace elements to describe mixing processes in water reservoir. Comptes-Rendus Géosci. 2012, 344, 25–32. [Google Scholar] [CrossRef]
- Mishra, A.; Alnahit, A.; Campbell, B. Impact of land uses, drought, flood, wildfire, and cascading events on water quality and microbial communities, a review and analysis. J. Hydrol. 2021, 596, 125707. [Google Scholar] [CrossRef]
- Espa, P.; Castelli, E.; Crosa, G.; Gentili, G. Environmental effects of storage preservation practices: Controlled flushing of fine sediment deom a small hydropower reservoir. Environ. Manag. 2013, 52, 261–276. [Google Scholar] [CrossRef]
- Frémion, F.; Courtin-Nomade, A.; Bordas, F.; Lenain, J.F.; Jugé, P.; Kestens, T.; Mourier, B. Impact of sediments resuspension on metal solubilisation and water quality during recurrent reservoir sluicing management. Sci. Total Environ. 2016, 562, 201–215. [Google Scholar] [CrossRef]
- Newcombe, C.P.; MacDonald, D.D. Effects of suspended sediments on aquatic ecosystems. N. Am. J. Fish. Manag. 1991, 11, 72–82. [Google Scholar] [CrossRef]
- Lors, C.; Tiffreau, C.; Laboudigue, A. Effects of bacterial activities on the releases of heavy metals from contaminated dredged sediments. Chemosphere 2004, 56, 619–630. [Google Scholar] [CrossRef]
- Fang, W.; Wei, Y.; Liu, J.; Kosson, D.S.; van der Sloot, H.A.; Zhang, P. Effects of aerobic and anaerobic biological processes on leaching of heavy metals from soil amended with sewage sludge compost. Waste Manag. 2016, 58, 324–334. [Google Scholar] [CrossRef]
- Wu, L.; Jacobson, A.D.; Chen, H.-C.; Hausner, M. Characterization of elemental release during microbe-basalt interaction at T = 28 °C. Geochim. Cosmochim. Acta 2007, 71, 2224–2239. [Google Scholar] [CrossRef]
- Wang, Q.; Gao, S.; Ma, X.; Mao, X.; He, L.; Sheng, X. Distinct mineral weathering effectiveness and metabolic activity between mineral-weathering bacteria Burkholderia metallica F22 and Burkholderia phytofirmans G34. Chem. Geol. 2018, 489, 38–45. [Google Scholar] [CrossRef]
- Dhivert, E.; Grosbois, C.; Coynel, A.; Lefèvre, I.; Desmet, M. Influences of major flood sediment inputs on sedimentary and geochemical signals archived in a reservoir core (Upper Loire Basin, France). Catena 2015, 126, 75–85. [Google Scholar] [CrossRef]
- Dhivert, E.; Grosbois, C.; Courtin-Nomade, A.; Bourrain, M.D. Dynamics of metallic contaminants at a basin scale—Spatial and temporal reconstruction from four sediment cores (Loire fluvial system, France). Sci. Total Environ. 2016, 541, 1504–1515. [Google Scholar] [CrossRef]
- Liu, S.; Yu, H.; Yu, Y.; Huang, J.; Zhou, Z.; Zeng, J.; Chen, P.; Xiao, F.; He, Z.; Yan, Q. Ecological stability of microbial communities in lake Donghue regulated by keystone taxa. Ecol. Indic. 2022, 136, 108695. [Google Scholar] [CrossRef]
- Gantner, S.; Andersson, A.F.; Alonso-Saez, L.; Bertilsson, S. Novel primers for 16S rRNA-based archaeal community analyses in environmental samples. J. Microbiol. Methods 2011, 84, 8–12. [Google Scholar] [CrossRef]
- Stahl, D.A.; Amann, R. Development and application of nucleic acid probes in bacterial systematics. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 1991; pp. 205–248. [Google Scholar]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analaysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Schloss, P. Reintroducing mothur, 10 years later. Appl. Environ. Microbiol. 2020, 86, e02343-19. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNE gene database project, improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Liu, X.; Li, M.; Castelle, C.J.; Probst, A.J.; Zhou, Z.P.; Banfield, J.F.; Gu, J. Insights into the ecology, evolution, and metabolism of the widespread Woesearchaeotal lineages. Microbiome 2018, 6, 102. [Google Scholar] [CrossRef]
- Zhou, Z.; Pan, J.; Wang, F.; Gu, J.; Li, M. Bathyarchaeota, globally distributed metabolic generalists in anoxic Environments. FEMS Microbiol. Rev. 2018, 42, 639–655. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 1 August 2020).
- Suzuki, R.; Shimodaira, H. Pvclust: An R package for assessing the uncertainty in hierarchical clustering. Bioinform 2006, 22, 1540–1542. [Google Scholar] [CrossRef]
- Shenhav, L.; Thompson, M.; Joseph, T.A.; Briscoe, L.; Furman, O.; Bogumil, D.; Mizrahi, I.; Péer, I.; Halperin, E. FEAST, fast expectation-maximization for microbial source tracking. Nat. Methods 2019, 16, 627–632. [Google Scholar] [CrossRef]
- Javanaud, C.; Michotey, V.; Guasco, S.; Garcia, N.; Anschutz, P.; Canton, M.; Bonin, P. Anaerobic ammonium oxidation mediated by Mn-oxides: From sediment to strain level. Res. Microbiol. 2011, 162, 848–857. [Google Scholar] [CrossRef]
- Fernandes, S.O.; Javanaud, C.; Aigle, A.; Garcia, N.; Anschutz, P.; Canton, M.; Bonin, P. Anaerobic nitrification-denitrification mediated by Mn-oxides in meso-tidal sediments: Implications for N2 and N2O production. J. Mar. Syst. 2015, 144, 1–8. [Google Scholar] [CrossRef]
- Jetten, M.S.M.; Strous, M.; van de Pas-Schooten, K.T.; Schalk, J.; van Dongen, U.G.J.M.; van de Graaf, A.A.; Logemann, S.; Muyzer, G.; van Loosdrecht, M.C.M.; Kuenen, J.G. The anaerobic oxidation of ammonium. FEMS Microbiol. Rev. 1998, 22, 421–437. [Google Scholar] [CrossRef]
- Dalsgaard, T.; Thamdrup, B.; Canfield, D.E. Anaerobic ammonium oxidation (anammox) in the marine environment. Res. Microbiol. 2005, 156, 457–464. [Google Scholar] [CrossRef]
- Yabuuchi, E.; Yano, I.; Oyaizu, H.; Hashimoto, Y.; Ezaki, T.; Yamamoto, H. Proposals of Sphingomonas paucimobilis gen. nov. and comb. Nov., Sphingomonas paracpaucimobilis sp. Nov., Sphingomonas yanoikuyae sp. Nov., Sphingomonas adhaesiva sp. Nov., Sphingomonas capsulate comb. Nov., and two genospecies of the genus Sphingomonas. Microbiol. Immunol. 1990, 34, 99–119. [Google Scholar] [CrossRef]
- Du, Y.; Yu, X.; Wang, G. Massilia tieshanensis sp. Nov., isolated from mining soil. Int. J. Syst. Evol. 2012, 62, 2356–2362. [Google Scholar] [CrossRef]
- Busse, H.J. Review of the taxonomy of the genus Arthrobacter, emendation of the genus Athrobacter sensus lato, proposal to reclassify selected species of the genus Arthrobacter in the novel genera Glutamicibacter gen. nov., Paeniglutamicibacter gen. nov. and Pseudoarthrobacter gen. nov., and emended description of Arthrobacter roseus. Int. J. Syst. Evol. 2016, 66, 9–37. [Google Scholar]
- Utkin, I.; Woese, K.; Wiegel, J. Isolation and characterization of Desulfitobacterium dehalogenans gen. nov., sp. nov., an anaerobic bacterium which reductively dechlorinates chlorophenolic compounds. Int. J. Syst. Evol. 1994, 44, 612–619. [Google Scholar] [CrossRef]
- Holmes, D.E.; Nevin, K.P.; Woodard, T.L.; Peacock, A.D.; Lovley, D.R. Prolixibacter bellariivorans gen. nov., sp. nov., a sugar-fermenting, psychrotolerant anaerobe of the phylum Bacteroidetes, isolated from a marine-sediment fuel cell. Int. J. Syst. Evol. 2007, 57, 701–707. [Google Scholar] [CrossRef]
- Shiratori, H.; Ohiwa, H.; Ikeno, H.; Ayame, S.; Kataoka, N.; Miya, A.; Beppu, T.; Ueda, K. Lutispora thermophila gen. nov., sp. nov., a thermophilic, spore-forming bacterium isolated from a thermophilic methanogenic bioreactor digesting municipal solid wastes. Int. J. Syst. Evol. 2008, 58, 964–969. [Google Scholar] [CrossRef]
- Ogg, C.D.; Patel, B.K.C. Caloramator australicus sp. nov., a thermophilic, anaerobic bacterium from the Great Artesian Basin of Australia. Int. J. Syst. Evol. 2009, 59, 95–101. [Google Scholar] [CrossRef]
- Sowers, K.R.; Baron, S.F.; Ferry, J.G. Methanosarcina acetivorans sp. nov., an acetotrophic methane-producing bacterium isolated from marine sediments. Appl. Environ. Microbiol. 1984, 47, 971–978. [Google Scholar] [CrossRef]
- Patel, G.B.; Sprott, G.D. Methanosaeta concilii gen. nov., sp. nov. (“Methanothrix concilii”) and Methanosaeta thermoacetophila nom. rev., com. nov. Int. J. Syst. Evol. 1990, 40, 78–82. [Google Scholar]
- Sakai, S.; Imachi, H.; Hanada, S.; Ohashi, A.; Harada, H.; Kamagata, Y. Methanocella paludicola gen. nov., sp. nov., a methane-producing archaeon, the first isolate of the lineage ‘Rice Cluster I’, and proposal of the new archaeal order Methanocellales ord. nov. Int. J. Syst. Evol. 2008, 58, 929–936. [Google Scholar] [CrossRef]
- Bräuer, S.L.; Cadillo-Quiroz, H.; Ward, R.J.; Yavitt, J.B.; Zinde, S.H. Methanoregula boonei gen. nov., sp. nov., an acidiphilic methanogen isolated from an acidic peat bog. Int. J. Syst. Evol. 2011, 61, 45–52. [Google Scholar] [CrossRef]
- Conrad, R.; Erkel, C.; Liesack, W. Rice Cluster I methanogens, an important group of Archaea producing greenhouse gas in soil. Curr. Opin. 2006, 17, 262–267. [Google Scholar] [CrossRef]
- Guerrero-Cruz, S.; Cremers, G.; van Alen, T.A.; Den Camp, H.J.M.O.; Jetten, M.S.M. Response of the anaerobic methanotroph “Candidatus Methanoperedens nitroreducens” to oxygen stress. Appl. Environ. Microbiol. 2018, 84, e01832-18. [Google Scholar] [CrossRef]
- Yu, T.; Wu, W.; Liang, W.; Wang, F. Growth of sedimentary Bathyarchaeota on lignin as an energy source. Proc. Natl. Acad. Sci. USA 2018, 115, 6022–6027. [Google Scholar] [CrossRef]
- Too, C.C.; Ong, K.S.; Lee, S.M.; Yule, C.M.; Keller, A. Draft genome sequence of Dyella sp. strain C9, isolated from a Malaysian tropical peal swamp forest. ASM 2018, 7, e01083-18. [Google Scholar]
- Hashidoko, Y.; Kitagawa, E.; Iwahashi, H. Design of Shingomonad-detecting probes for DNA array, and its application to investigate the behavior, distribution, and source of rhizospherous Sphingomonas and other Sphingomonads inhabiting an acid sulfate soil paddock in Kalimantan, Indonesia. Biosci. Biotechnol. Biochem. 2007, 71, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Weon, H.Y.; Anandham, R.; Kim, B.Y.; Hong, S.B.; Jeon, Y.A.; Kwon, S.W. Dyella soli sp. and Dyella terrae sp. nov., isolated from soil. Int. J. Syst. Evol. 2009, 59, 1685–1690. [Google Scholar] [CrossRef] [PubMed]
- Dedysh, S.N.; Liesack, W.; Khelenina, V.N.; Suzina, N.E.; Trotsenko, Y.A.; Semrau, J.D.; Bares, A.M.; Panikov, N.S.; Tiedje, M.J. Methylocella palutris gen. nov., sp. nov., a new methane-oxidizing acidophilic bacterium from peat bogs, representing a novel subtype of serine-pathway methanotrophs. Int. J. Syst. Evol. 2000, 50, 955–969. [Google Scholar] [CrossRef]
- Nalin, R.; Simonet, P.; Vogel, T.M.; Normand, P. Rhodanobacter lindaniclastus gen. nov., sp. nov., a lindane-degrading bacterium. Int. J. Syst. Evol. 1999, 49, 19–23. [Google Scholar] [CrossRef]
- Dziuba, M.; Koziaeva, V.; Grouzdev, D.; Burganskaya, E.; Baslerov, R.; Kolganova, T.; Chernyadyev, A.; Osipov, G.; Andrianova, E.; Gorlenko, V. Magnetospirillum caucaseum sp. nov., Magnetospirillum marisnigri sp. nov. and Magnetospirillum moscoviense sp. nov., freshwater magnetotactic bacteria isolated from three distinct geographical locations in European Russia. Int. J. Syst. Evol. 2016, 66, 2069–2077. [Google Scholar] [CrossRef]
- Bagnara, C.; Toci, R.; Gaudin, C.; Belaich, J.P. Isolation and characterization of a cellulolytic microorganism, Cellulomonas fermentans sp. nov. Int. J. Syst. Evol. 1985, 35, 502–507. [Google Scholar]
- Abraham, W.R.; Strömpl, C.; Meyer, H.; Lindholst, S.; Moore, E.; Christ, R.; Vancanneyt, M.; Tindall, B.; Bennasar, A.; Smit, J.; et al. Phylogeny and polyphasic taxonomy of Caulobacter species. Proposal of Maricaulis gen. nov. with Maricaulis maris (Poindexter) comb. nov. as the type species, and emended description of the genera Brevundimonas and Caulobacter. Int. J. Syst. Evol. 1999, 49, 1053–1073. [Google Scholar] [CrossRef]
- Vaksmaa, A.; Guerrero-Cruz, S.; van Alen, T.A.; Cremers, G.; Ettwig, K.F.; Claudia Lüke, M.S.M.J. Enrichment of anaerobic nitrate-dependant methanotrophic ‘Candidatus Methanoperedens nitroreducens’ archaea from an Italian paddy field soil. Appl. Microbiol. Biotechnol. 2017, 101, 7075–7084. [Google Scholar] [CrossRef]
- Spang, A.; Poehlin, A.; Offre, P.; Zumbrägel, S.; Haider, S.; Rychlik, N.; Nowka, B.; Schmeisser, C.; Lebedeva, E.V.; TRattei, H.; et al. The genome of the ammonia-oxidizing Candidatus Nitrososphaera gargensis, insights into metabolic versatility and environmental adaptations. Environ. Microbiol. 2012, 14, 3122–3145. [Google Scholar] [CrossRef]
- Adamczuk, M. Do experimental conditions bias plankton responses to increased concentration of dissolved organic matter (DOM)? A meta-analytical synthesis of the published results. Sci. Total Environ. 2021, 787, 147590. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Df | SumOfSqs | R2 | F | Pr (>F) | |
| Oxygen | 1 | 0.9022 | 0.1377 | 2.9917 | 0.002 |
| Environment | 1 | 1.0081 | 0.15387 | 3.3429 | 0.001 |
| Incubation time | 2 | 1.0226 | 0.15609 | 1.6956 | 0.04 |
| Residual | 12 | 3.6187 | 0.55234 | ||
| Total | 16 | 6.5515 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
DesRosiers, A.; Gassama, N.; Grosbois, C.; Lazar, C.S. Laboratory-Controlled Experiments Reveal Microbial Community Shifts during Sediment Resuspension Events. Genes 2022, 13, 1416. https://doi.org/10.3390/genes13081416
DesRosiers A, Gassama N, Grosbois C, Lazar CS. Laboratory-Controlled Experiments Reveal Microbial Community Shifts during Sediment Resuspension Events. Genes. 2022; 13(8):1416. https://doi.org/10.3390/genes13081416
Chicago/Turabian StyleDesRosiers, Alexis, Nathalie Gassama, Cécile Grosbois, and Cassandre Sara Lazar. 2022. "Laboratory-Controlled Experiments Reveal Microbial Community Shifts during Sediment Resuspension Events" Genes 13, no. 8: 1416. https://doi.org/10.3390/genes13081416
APA StyleDesRosiers, A., Gassama, N., Grosbois, C., & Lazar, C. S. (2022). Laboratory-Controlled Experiments Reveal Microbial Community Shifts during Sediment Resuspension Events. Genes, 13(8), 1416. https://doi.org/10.3390/genes13081416